基于高通量测序技术解析三七不同部位内生细菌多样性
2021-12-14李秋桦夏体渊陈泽斌唐佐芯字学庆
李秋桦,夏体渊,任 禛*,裴 妍,尹 敏,陈泽斌,唐佐芯,字学庆
(1.昆明学院农学与生命科学学院/云南省高校生物炭工程研究中心,云南 昆明 650214;2.云南大学医学院,云南 昆明 650091)
【研究意义】三七(Panaxnotoginseng)是中国重要的中药材之一,有着“金不换”的美誉,它属于五加科人参属草本植物,云南文山是三七的原产地和主产地。三七具有极高的医疗价值,可以预防心脑血管疾病,降低三高以及消肿止痛等[1]。鉴于三七在中国中药材中的重要地位,对其进行生理生长、抗病耐病等方面的研究具有重要意义。植物内生菌在植物根、茎和叶等组织中广泛存在,是植物微生态系统的重要组成部分[2-4]。大部分植物内生菌不会使植物致病,并且可以与植物建立共生关系。在药用植物中,内生菌可以与植物协同进化,产生与植物相同或相似的生物活性物质[5]。【前人研究进展】目前研究发现植物内生细菌具有促进植物生长,降低病虫害等诸多功能[6-9]。刘莹等[5]运用纯培养法在三七植物组织中分离出11株内生菌,并筛选出一株可以产皂苷的内生菌株,经鉴定为肠杆菌属(Enterobacter)细菌。张玉洁等[10]对三七内生真菌进行分离,发现三七体内存在大量的内生真菌资源,并且通过抑菌实验可以得到众多抗根腐病病原菌活性的菌株。沈永昶等[11]通过对三七内生细菌进行筛选,发现了能够抑制三七根腐病原菌的菌株,经鉴定为解淀粉芽孢杆菌(Bacillusamyloliquefaciens)和枯草芽孢杆菌(B.subtilis)。可见,三七内含有丰富的内生菌资源,并在抵抗病害等方面具有积极作用。对三七内生细菌进行多样性研究,可为筛选促进三七生长以及增强其抗病性的优良菌株奠定基础。【本研究切入点】目前对三七内生细菌多样性的研究方式主要为纯培养,应用免培养技术的研究报道较少,三七中大部分内生细菌还未被检测到,限制了三七内生细菌资源的开发和利用。【拟解决的关键问题】采用高通量测序技术对云南文山三七根、茎和叶的内生细菌多样性进行分析。旨在全面揭示文山三七不同部位的内生细菌群落结构,为进一步探究三七内生细菌的生态功能以及互作机制提供理论基础。
1 材料与方法
1.1 样本采集
2020年1月在云南省文山市新沟农场采集的健康、无病虫害的三七植株,用清水洗净表面泥土后备用。该地区海拔约1662 m,年平均气温16.4 ℃,平均降水量为1206.8 mm,平均日照约2056.2 h,经纬度为104°5′40′′E,24°8′40′′N。
1.2 表面消毒
取新鲜健康的三七根、茎、叶,试验设置3组处理,每组样本3个重复,叶部样品标记为(PNL):PNLl、PNL2、PNL3;茎部样品标记为(PNS):PNSl、PNS2、PNS3;根部样品标记为(PNR):PNR1、PNR2、PNR3。在无菌条件下分别用蒸馏水进行多次冲洗,再用70%乙醇浸泡3次,每次浸泡1 min,最后用无菌水冲洗3~4次。随后提取样品的DNA。
1.3 基因组DNA提取
将消毒后的三七根、茎和叶放入液氮中迅速降温,保护DNA不被破坏,然后根据TIANGEN植物基因组DNA提取试剂盒的说明对样品进行DNA提取。采用1%琼脂糖凝胶检测DNA的浓度和纯度,并分别加入超纯水将DNA样品浓度稀释到1 ng/μL。
1.4 PCR扩增及高通量测序
对样品进行16S rRNA基因V5~V7区的PCR扩增,上下游引物分别为799F(5′-AACMGGATTAGATACCCKG-3′)和1193R(5′-ACGTCATCCCCACCTTC C-3′),并引入接头。扩增体系:10×PCR buffer 5 μL,dNTP 4 μL,正反向引物各1 μL,DNA模板1 μL,Taq酶0.4 μL,补足ddH2O至50 μL。反应程序:95 ℃预变性5 min;94 ℃ 60 s,56 ℃ 60 s,72 ℃ 90 s,循环30次;最后在72 ℃延伸10 min。将获得的PCR产物经琼脂糖凝胶电泳检测后,送至上海美吉生物公司进行高通量测序。
1.5 序列登录
所有经过重组和筛选的原始序列都存储在NCBI SRA数据库中,叶部3次重复(PNLl、PNL2和PNL3)的登录号分别为SRR12078548、SRR12078549和SRR12078550;茎部3次重复(PNSl、PNS2和PNS3)的登录号分别为SRR12078551、SRR12078552和SRR12078553;根部3次重复(PNRl、PNR2和PNR3)的登录号分别为SRR12078554、SRR12078555和SRR12078556。
2 结果与分析
2.1 三七内生细菌OTU及多样性分析
如表1所示,三七根、茎和叶产生的原始序列在质控后获得V5~V7可变区的平均有效序列总数分别为24 334、18 094和22 064条,去除叶绿体和线粒体的序列数量之后,分别得到14 717、13 870和19 717条序列。同时按照97%相似性进行归类,三七根、茎和叶获得的可操作分类单元(OTUs)的均值分别为385、206和226。通过单因素方差分析可以看出,三七3个部位的有效序列数差异不显著,但叶部的细菌序列数量明显高于其它部位,而OTU数量整体表现为根>叶>茎,且根部的OTU数量显著高于其它部位。
Alpha多样性指数结果(表2)表明,三七根、茎和叶的Shannon指数和Simpson指数无显著性差异,但根部的ACE指数和Chao指数值显著高于叶部和茎部,说明根部的微生物群落丰富度较高,物种种类较多。此外,从Rank abundance曲线图(图1)可以看出,根部下降较缓,而茎部和叶部下降较快,说明三七根部的内生细菌丰富度较高,这与多样性指数结果一致。

表1 三七不同部位的OTUs及相关序列指数

表2 三七不同部位内生细菌多样性指数
OTU稀释曲线(图2-A)结果表明,随着测序深度的增加,OTU数量增幅变缓。同时,从Shannon指数稀释曲线(图2-B)结果可以看出,当测序深度大于3000时,三七根、茎和叶的Shannon指数曲线逐渐趋平,说明群落种类的多样性不再增加。此外,所测样品的OTU覆盖率均高达99%以上(表2)。综合上述结果来看,3个样本测序数量足够,可真实反应三七不同组织部位内生细菌群落的组成。

图1 Rank abundance曲线图Fig.1 Rank abundance curves
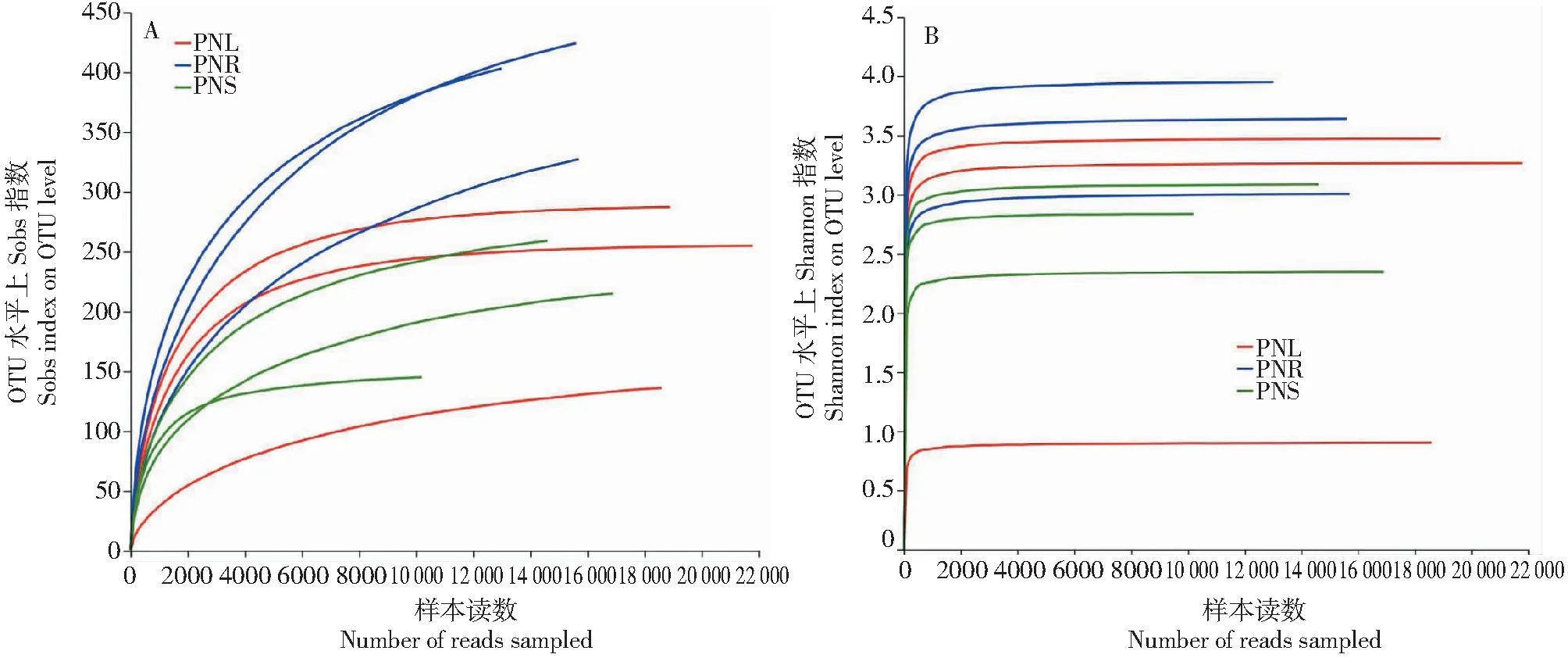
图2 三七不同部位内生细菌OTU稀释曲线(A)和Shannon指数稀释曲线(B)Fig.2 Rarefaction curves of the number of OTUs (A) and Shannon indexes (B) of bacterial endophyte communities detected in different parts of Panax notoginseng
2.2 ANOSIM分析
ANOSIM分析中Between为分组之间的差异,PNL、PNS、PNR分别代表叶、茎、根各自的组内差异,纵坐标刻度表示距离值。由ANOSIM分析(图3)可以看出,P值为0.011<0.05,表示组间和组内的差异均具有统计学意义,R值为0.6049>0,且接近1,说明组间差异远大于组内的差异,且组间具有显著性差异。
2.3 三七不同部位内生细菌群体聚类分析
采用主坐标分析(PCoA)和偏最小二乘法判别分析(PLS-DA)对样本进行分组考察,对不同样本内生细菌群体进行聚类分析。基于所选距离矩阵进行构建PCoA图(图4-A),结果显示,三七根部的群落结构相似性较高,能够较好的聚集;叶部聚集性较低,群落结构相似性有所差异;茎部较为散乱,说明群落结构相似性较低。基于物种丰度矩阵和样本分组数据构建的PLS-DA图(图4-B),可以看出根、茎、叶各部位的计算结果均可以聚集在同一象限内,能够较好的区分,表明不同分组的内生细菌群落结构在组间产生了显著差异,且在组内具有较高的群落结构相似性。

图3 Anosim分析Fig.3 Anosim analysis
2.4 三七内生细菌种群归类分析
排除不可分类的细菌类型后再进行种群归类(表3),结果发现三七叶部内生细菌归属于16个门,27个纲,74个目,134个科,222个属;茎部内生细菌归属于12个门,20个纲,58个目,108个科,178个属;根部内生细菌归属于19个门,24个纲,75个目,137个科,202个属。从表3可以看出,三七不同部位的内生细菌在门、纲、目、科和属等分类阶元上的数量明显不同,但在各阶层呈现的规律较为一致,整体表现为根>叶>茎,这与三七不同部位内生细菌的OTU数结果(表1)指向一致。
2.5 三七不同部位内生细菌在不同分类水平的群落组成分析
2.5.1 OTU水平分析 Venn图可以直观比较三七根、茎和叶样品的OTU组成相似性和重叠情况。基于Venn图分析(图5),三七根、茎和叶样品的全部细菌OTU数为877个,其中共有的OTU数目为167个,约占全部的19.04%,这些OTU主要由放线菌门(Actinobacteria)、变形菌门(Proteobacteria)、厚壁菌门(Firmicutes)组成。根部独有的OTU数目较多,约占全部的32.61%;叶部次之,约占全部的18.36%;茎部独有的OTU数目较少,约占全部的9.46%。由此可以看出,茎部与其它部位的细菌的OTU数量差异区别较大,且明显低于其它部位。

图4 三七不同部位内生细菌群落PCoA图(A)和PLS-DA图(B)Fig.4 The PCoA (A) and PLS-DA (B) analysis of endophytic bacterial community in different parts of Panax notoginseng

表3 三七不同部位内生细菌在各分类阶层的总数
2.5.2 门水平分析 三元相图(图6)可以直观的反映三七不同部位内生细菌在门水平上的分布比重和关系,图中每个圆圈代表不同细菌属,圆圈颜色相同表示属于同一个门。结果显示放线菌门(Actinobacteria)在三七叶部的分布比重较高,变形菌门(Proteobacteria)在三七茎部和根部的分布比重较高。同时,可归类门水平的群落多样性(图7)表明,在三七叶部丰度水平前5的菌门为放线菌门(Actinobacteria,52.78%)、变形菌门(Proteobacteria,38.87%)、厚壁菌门(Firmicutes,6.92%)、拟杆菌门(Bacteroidetes,0.66%)和酸杆菌门(Acidobacteria,0.51%);在茎部丰度水平前5的菌门为变形菌门(Proteobacteria,69.77%)、放线菌门(Actinobacteria,22.78%)、厚壁菌门(Firmicutes,6.38%)、拟杆菌门(Bacteroidetes,0.45%)和酸杆菌门(Acidobacteria,0.34%);在根部丰度水平前5的菌门为变形菌门(Proteobacteria,78.99%)、放线菌门(Actinobacteria,14.12%)、厚壁菌门(Firmicutes,2.42%)、酸杆菌门(Acidobacteria,1.23%)和拟杆菌门(Bacteroidetes,0.82%)。从上述结果可以看出,三七不同部位内生细菌的优势门类型相同,但各优势门在各部位内生细菌总量的占比并不相同。

图5 OTU水平的Venn图分析Fig.5 Venn analysis on OTU level

图6 三七不同部位内生细菌群落在门水平上的三元相图Fig.6 Ternary analysis of endophytic bacterial community atphylum level in different parts of Panax notoginseng

图7 三七不同部位内生细菌群落在门水平上的群落多样性分析图Fig.7 Community diversity analysis of endophytic bacterial community atphylum level in different parts of Panax notoginseng
2.5.3 属水平分析 三元相图(图8)结果可以反映不同细菌属在三七不同部位中的分布差异,图中每个圆圈代表不同细菌种,圆圈颜色相同表示属于同一个属。其中Cutibacterium在三七叶部的分布比重较高;假单胞菌属(Pseudomonas)在三七茎部的分布比重较高;Burkholderia-Caballeronia-Paraburkholderia和埃希氏-志贺菌属(Escherichia-Shigella)在三七根部的分布比重较高。从归类属水平的群落多样性(图9)显示,三七叶部丰度水平前5的菌属为Cutibacterium(28.70%)、劳尔氏菌属(Ralstonia,14.57%)、红球菌属(Rhodococcus,12.34%)、Burkholderia-Caballeronia-Paraburkholderia(11.94%)和甲基杆菌属(Methylobacterium,4.63%);茎部丰度水平前5的菌属为假单胞菌属(Pseudomonas,45.50%)、红球菌属(Rhodococcus,6.75%)、链霉菌属(Streptomyces,5.20%)、劳尔氏菌属(Ralstonia,4.13%)和Burkholderia-Caballeronia-Paraburkholderia(3.78%);根部丰度水平前5的菌属为Burkholderia-Caballeronia-Paraburkholderia(24.78%)、埃希氏-志贺菌属(Escherichia-Shigella,21.94%)、鲍特菌属(Bordetella,5.23%)、链霉菌属(Streptomyces,5.13%)和不动杆菌属(Acidibacter,3.91%)。可见,三七不同部位内生细菌的优势属种类有明显差异,仅Burkholderia-Caballeronia-Paraburkholderia为3者共有的优势属,但该属在不同部位的丰度占比不同,同时也可以看出三七根部与其它部位的优势属种类差异较大。
2.6 三七不同部位内生细菌系统进化分析
选取在属水平中丰富度位于前20且可进行分类的序列构建系统进化树,并以环形图(图8)的形式呈现物种的系统发生进化关系。图中每条线代表一个属,线条长度为物种差异距离,线条颜色相同则表示属于同一个门。图中绿色为厚壁菌门(Firmicutes),该门有3个属,且物种差异程度较小;红色为放线菌门(Actinobacteria),该门有5个属,且进化距离较近;紫色为变形菌门(Proteobacteria),该门有12个属,且部分属的物种差异程度较大。
2.7 三七不同部位内生细菌群落的功能预测分析
使用COG数据库对三七不同部位内生细菌群落进行功能预测(图9),在不同部位中共获得了23个功能分组,但丰度随部位的不同而异。在三七内部,丰度较高的微生物群落功能为:氨基酸转运与代谢功能(Amino acid transport and metabolism,8.59%~9.07%)、转录功能(Transcription,7.33%~8.01%)、能量产生和转换功能(Energy production and conversion,7.12%~7.35%)和无机离子运输与代谢功能(Inorganicion transport and metabolism,6.10%~6.53%)。另外,叶部具有碳水化合物运输和代谢(Carbohydratetransport and metabolism)功能的细菌丰度较高;茎部具有信号转导(Signal transduction mechanisms)功能的细菌丰度较高;根部能够合成细胞壁、细胞膜等物质(Cell wall/membrane/envelopebiogenesis)功能的细菌丰度较高。

图8 三七不同部位内生细菌群落属水平的三元相图Fig.8 Ternary analysis of endophytic bacterial community at genus level in different parts of Panax notoginseng

图9 三七不同部位内生细菌群落属水平的群落多样性分析Fig.9 Community diversity analysis of endophytic bacterial community at genus level in different parts of Panax notoginseng
3 讨 论
3.1 高通量测序技术在植物内生细菌研究中的应用
植物内生菌广泛存在于植物的各个器官和组织中,是植物体内的正常菌群,包括众多的互惠共生细菌,它们不仅不会使植物受到伤害,还可以在植物的生长发育等过程中发挥重要作用[12-14],所以对植物内生菌的研究与开发尤为重要。目前,在植物内生菌多样性的研究中运用较多的方法是高通量测序法,该技术具有信息量大、可靠性高等优点[15-18],与纯培养相比可以获得较为全面的生物种群信息,并且可准确的反映植物内生微生物的种类组成和真实比例[19]。张爱梅等[20]对榆中县中国沙棘种子内生细菌多样性进行研究时,采用纯培养的方法仅得到3个属的内生细菌,而采用高通量测序得到68个属的内生细菌。刘晓静等[21]对库车小白杏内生细菌进行高通量测序,共检测出94个属,而采用纯培养的方法仅分离获得28个属的菌株。符玉松[22]采用高通量测序技术在菖蒲的根部检测到56个科的细菌类型,而采用纯培养法分离仅得到12个科的细菌种类。可见,应用高通量测序技术可以克服纯培养的不足,能够更准确全面地检测到植物内生细菌种类。

图10 物种环形进化树图Fig.10 Phylogenetic tree

图11 COG功能分类统计柱状图Fig.11 COG function classification
3.2 三七内生细菌在不同部位的分布特征
采用高通量测序技术检测文山三七不同部位内生细菌,在三七根、茎、叶组织中分别获得了385、206和226个OTU,其中共有的OTU数为167个,占比19.04%。此外,Alpha多样性指数结果表明三七各部位中均含有丰富的内生细菌种类,其中根部的群落丰富度较高,茎部和叶部较低。众多研究表明,内生菌的群落多样性受多种因素的影响,同一植物不同部位的内生菌亦有差异。林丽等[23]应用高通量测序法检测烟草不同部位内生细菌,发现不同部位的Alpha多样性指数为茎>根>叶,呈现明显的器官差异性。邓平香等[24]研究2种生态型东南景天茎、叶内生细菌多样性,结果均表现出叶>茎,呈现内生细菌多样性因部位不同而异。沙月霞[25]研究水稻不同部位内生细菌多样性,发现宁夏水稻品种宁粳43号叶部内生细菌群落的丰富度、群落多样性明显较高,茎部次之,根部的多样性水平较低。
通过高通量测序分析鉴定出三七叶部内生细菌归属于16个门,27个纲,74个目,134个科,222个属;茎部内生细菌归属于12个门,20个纲,58个目,108个科,178个属;根部内生细菌归属于19个门,24个纲,75个目,137个科,202个属,呈现出三七各部位的内生细菌在不同分类阶元上的数量为根>叶>茎。在门水平上,各部位的优势门类型相同,但丰度不同;在属水平上,各部位的优势类型均有差异,但不同部位仍具有相同的优势类型。其中,红球菌属(Rhodococcus)为三七叶部和茎部的优势属,据相关研究报道,该属菌在工业合成转化中有广泛应用,且在预防环境污染以及环境治理等生物技术上有很广阔的应用前景[26]。链霉菌属(Streptomyces)为三七根部和茎部的优势属,该菌属的细菌能够产生抗生素和植物生长激素类物质,在促进植物生长、提高抗逆性等方面发挥重要作用[27]。因此,在三七不同部位存在一定的功能菌株,具有一定的研究利用价值。
3.3 三七不同部位内生细菌群落功能差异
在三七根、茎、叶中既发现了相同物种又发现了特有物种,可能是由于土壤中的微生物通过根部进入植物体内,部分微生物向上输送到茎部以及叶部,同时叶部可以通过气孔和叶片表皮毛等让叶围的微生物进入植物体内,部分微生物可以向下输送至其它部位,使得根、茎和叶有着相同的物种,但由于后期不同部位周围环境的不同,导致出现特有物种[25];植物茎部与外界微生物的交换能力较弱,可能是该部位内生细菌较少的主要原因。三七内生细菌参与植物组织中物质转运和代谢、能量产生与转换、细胞壁和细胞膜的合成、信号转导以及转录等。叶部参与各种功能活动的物种丰度最高,其碳水化合物运输和代谢功能优于其它部位,可以推测由于叶部进行光合作用产生的有机物需要向茎部和根部输送,所以叶部该功能物种丰度较高;根部参与各种功能活动的物种丰度较高,其产生细胞壁、细胞膜等物质的功能优于其它部位,可能由于根部细胞壁为土壤与植物的交界面,是抵御重金属胁迫、土传病害侵入以及修复调控过程的关键部位,所以根部该功能物种丰度较高;茎部参与各种功能活动的物种丰度较低,其信号转导功能优于其它部位,可以推测由于茎部是信号传输、营养输送上通下达的必经之路,所以茎部该功能物种丰度较高。
4 结 论
首次采用高通量测序技术对三七不同部位的内生细菌多样性进行分析,并对各部位内生细菌在不同分类水平的群落组成进行探究。结果发现三七根、茎和叶部位中均含有丰富的内生细菌,且不同部位的丰度不同,整体表现为根>叶>茎。此外,在三七不同部位中发现部分优势属细菌具有一定的功能作用,可进行后续的开发与利用。三七内生细菌在植物体内的代谢以及调控等方面均发挥积极作用,对其进一步的研究尤为重要。三七不同部位内生细菌多样性的研究结果为后续筛选生防菌株奠定基础。
