高山杜鹃离体叶片高效再生及其再生途径研究
2021-12-13彭绿春廖多思解玮佳蔡艳飞李世峰
彭绿春,廖多思,解玮佳,宋 杰,蔡艳飞,张 露,李世峰
(1云南省农业科学院花卉研究所/国家观赏园艺工程技术研究中心/云南省花卉育种重点实验室/云南花卉技术工程研究中心,昆明 650205;2宁南县林业和草原局,四川 凉山 615400)
0 引言
杜鹃花属是世界著名的木本花卉。高山杜鹃是指利用杜鹃花属中常绿杜鹃亚属种质资源育成的一些革质、常绿、花冠硕大的品种类型[1],其观赏价值高,近年来成为中国年宵花和高档园林造景的新秀,高山杜鹃新品种培育在国内也日益引起重视[2]。因在打破种间杂交障碍、定向选育方面较常规杂交育种表现的优越性,基因工程技术已成为现今杜鹃花育种的热点问题[3]。叶片是遗传转化和细胞操作的常用受体材料[4-6],建立高效、稳定的离体叶片再生植株体系和发掘再生能力强的基因型,是杜鹃花基因工程育种成功的关键。
目前,杜鹃花植株离体再生方面的研究很多,但以离体叶片为材料进行的植株再生研究较少,主要涉及基本培养基类型、激素种类和浓度等方面[7-9]。研究认为低盐浓度的Anderson培养基、Read培养基和WPM培养基适用于大多数杜鹃的离体培养[10-12]。外源激素的选用主要是TDZ和ZT,且针对不同基因型杜鹃花离体叶片再生,2种激素诱导效果差异大。Zaytseva等[13]用 TDZ诱导Rhododendron sichotense和Rhododendron catawbiense‘Grandiflorum’的离体叶片再生不定芽,再生频率分别达93%和85%,且再生的芽均起源于叶片近轴面的表皮细胞,但前者来源于突起,而后者来源于胚状体结构。刘燕等[14]认为TDZ较ZT和2-iP更有利于桃叶杜鹃诱导愈伤组织。汪玲敏等[15]则认为诱导云南杜鹃离体叶片直接再生芽ZT作用好于TDZ,再生芽率为74%。吕秀立等[16]应用ZT诱导大叶常绿杜鹃离体直接再生不定芽率为70%。Pavingerova[17]以不同浓度TDZ诱导15个杜鹃栽培种离体叶片再生不定芽的研究发现,不同基因型所需TDZ浓度不同,且不定芽的再生频率从0%到100%差异很大。为建立高效、稳定的杜鹃离体叶片植株再生体系,一方面有必要针对特定种和基因型开展植株再生研究,一方面还需筛选再生能力强的基因型应用于遗传转化。
本研究以6个高山杜鹃品种为材料,研究不同浓度TDZ和ZT对其离体叶片再生效率的影响,筛选出再生能力强的基因型,并通过再生过程形态学和组织细胞学观察对再生途径进行初步研究,以期为高山杜鹃离体叶片再生芽机理研究,提高其植株再生频率和遗传转化效率提供技术支持和试验基础。
1 材料与方法
1.1 试验时间、地点
试验于2018年6月—2019年12月在云南省农业科学院花卉研究所云南省花卉育种重点实验室进行。
1.2 试验材料
试验材料为云南省农业科学院花卉研究所组织培养实验室中保存的6个高山杜鹃品种的增殖瓶苗,分别 是‘Percy Wiseman’(D4)、‘Cunnigham’s White’(W10)、‘Halfdem Lem’(W17)、‘Madame Masson’(W22)、‘Scintillation’(W29)、‘XXL’(X)。
1.3 试验方法
1.3.1 不定芽诱导培养 以增殖培养30天的高山杜鹃瓶苗为材料,取植株顶部往下完全展开的第1和第2轮叶片,叶背划3条伤口,背面朝下接种于附加不同浓度ZT、TDZ和NAA的诱导培养基,激素配比见表1,每个配方接种3皿,每皿7个叶片。接种后每隔1天观察记录每个品种在不同浓度激素处理下的分化时间和分化的叶片数(叶片上只要有不定芽长出,则该叶片为分化的叶片),培养50天后,统计分化叶片数并计算分化率[式(1)~(2)];此时将试验材料转接到相应处理的新鲜培养基,再培养40天后,计数不同处理下各品种分化芽的数量[式(3)]。
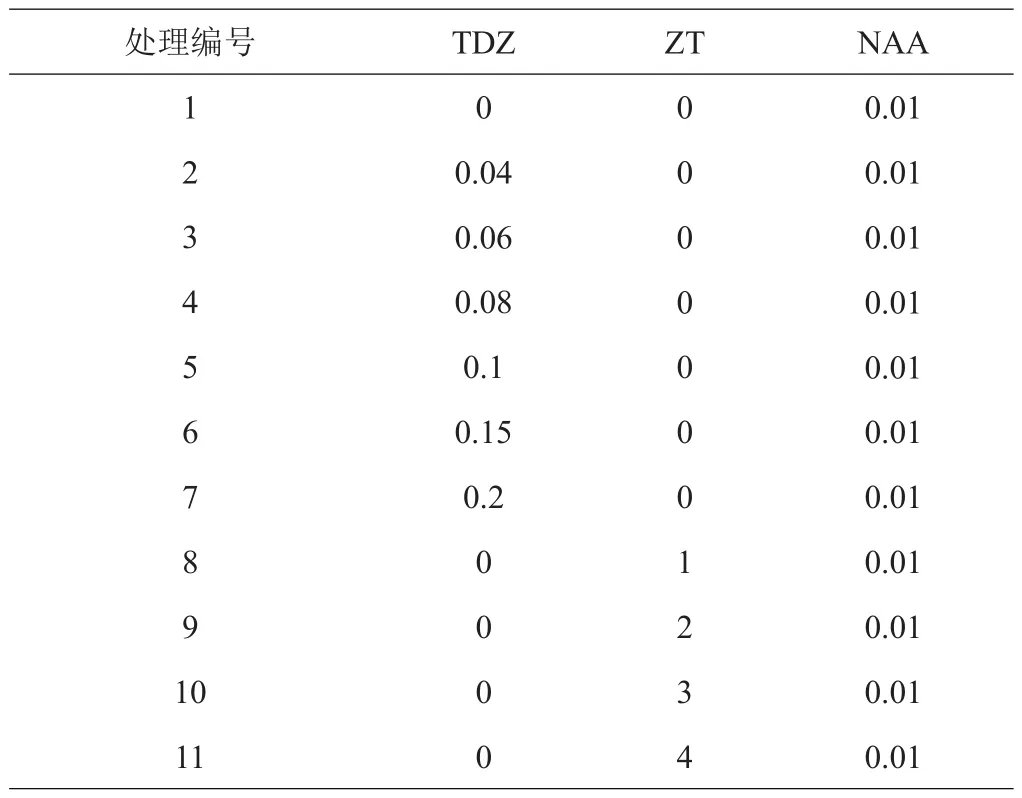
表1 离体叶片诱导芽激素组合 mg/L

诱导培养基以WPM为基本培养基,附加不同配比的激素,蔗糖30 g/L、琼脂8 g/L,pH 5.4,121℃高压灭菌25 min。培养光照强度2000 lx,光照时间12 h/d,培养温度(25±2)℃。
1.3.2 离体叶片分化不定芽形态及组织细胞学观察 接种后,用体视镜观察离体叶片的形态响应,接种后至培养第30天隔1天观察,培养30天后隔5天观察,记录离体叶片分化芽的时间和形态学特征。离体叶片分化形成完整的不定芽之前,每次观察时取培养中的叶片进行石蜡切片,观察叶片分化过程中的组织细胞学变化,石蜡切片制作参照Zaytseva1等[13]的方法进行。
1.4 数据处理
原始数据用Excel统计、计算,采用SPSS Statistics 18.0软件分析不同处理间分化率和分化不定芽数的差异显著性,显著水平P<0.05。
2 结果与分析
2.1 激素种类和浓度对高山杜鹃离体叶片再生芽效率的影响
2.1.1 分化率 表2结果显示,不添加外源细胞分裂素即可一定程度诱导6种高山杜鹃离体叶片切口处的膨大反应,并最终有极少量不定芽形成。从分化率看,TDZ浓度为0.04 mg/L时,除D4和W22外其余品种的不定芽分化率均可达到100%。而D4和W22的分化率也是随TDZ浓度增加而提高,分别于0.08、0.06 mg/L时达到100%。ZT对6个基因型高山杜鹃离体叶片不定芽的诱导效果显著弱于TDZ。随ZT浓度增加,各基因型叶片的分化率提高。当ZT浓度为4 mg/L时,W10、W17、W29、X分化率与TDZ 0.04 mg/L处理无显著差异,但D4和W22的分化率显著低于TDZ处理。

表2 不同处理对6种高山杜鹃离体叶片分化率的影响%
从响应时间来看,在各自分化率最高的TDZ浓度下,W10、W29和X的响应最早,培养3~5天即可见叶片切口的膨大反应(图1)。W29培养18天后离体叶片即全部开始分化不定芽,X培养20天后分化率达100%,W10培养22天后全部分化,W17培养28天后叶片分化率达100%,而W22和D4叶片培养35天后分化率达100%。虽然6个基因型高山杜鹃的叶片分化率均达到了100%,但其分化出的不定芽状态有差异,且从离体叶片对外源TDZ的反应时间看,W29最早,W10和X稍晚,再次是W17,W22和D4最晚。
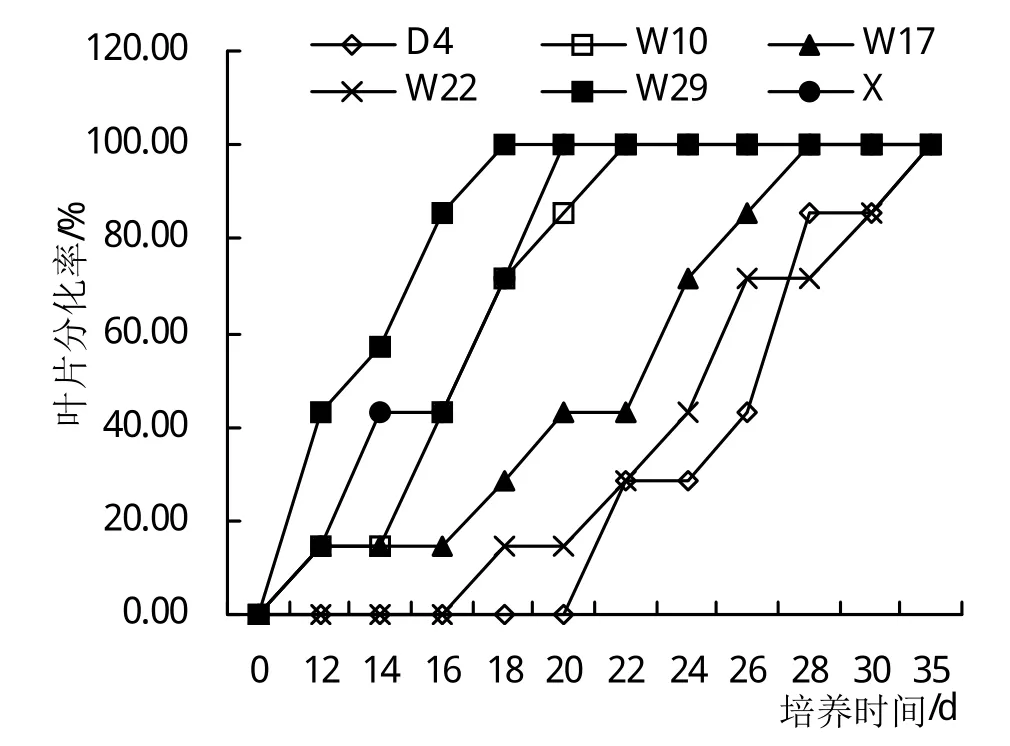
图1 不同基因型高山杜鹃离体叶片分化趋势
2.1.2 再生不定芽数量 从表3可看出,各基因型中,TDZ诱导的不定芽数量显著高于ZT。对于同一个基因型,不同TDZ浓度下离体叶片分化不定芽的数量表现出差异,随浓度升高,芽数量呈先增加后降低的趋势,其中,D4、W17、W29、X在TDZ为0.06 mg/L时分化的芽数最多,显著多于其他处理,W10和W22在TDZ 0.04~0.06 mg/L时分化的芽数最多,显著多于其他处理。各基因型高山杜鹃中,W10离体叶片诱导出的不定芽最多,平均达13个/片,而X诱导出的芽数最少,平均只有5.67个/片。通过形态观察发现,各基因型在TDZ浓度高于0.06 mg/L时,可分化出大量密集的芽点,但这些芽点大部分不能伸长生长发育成正常的芽,故统计其分化的芽数量反而降低。
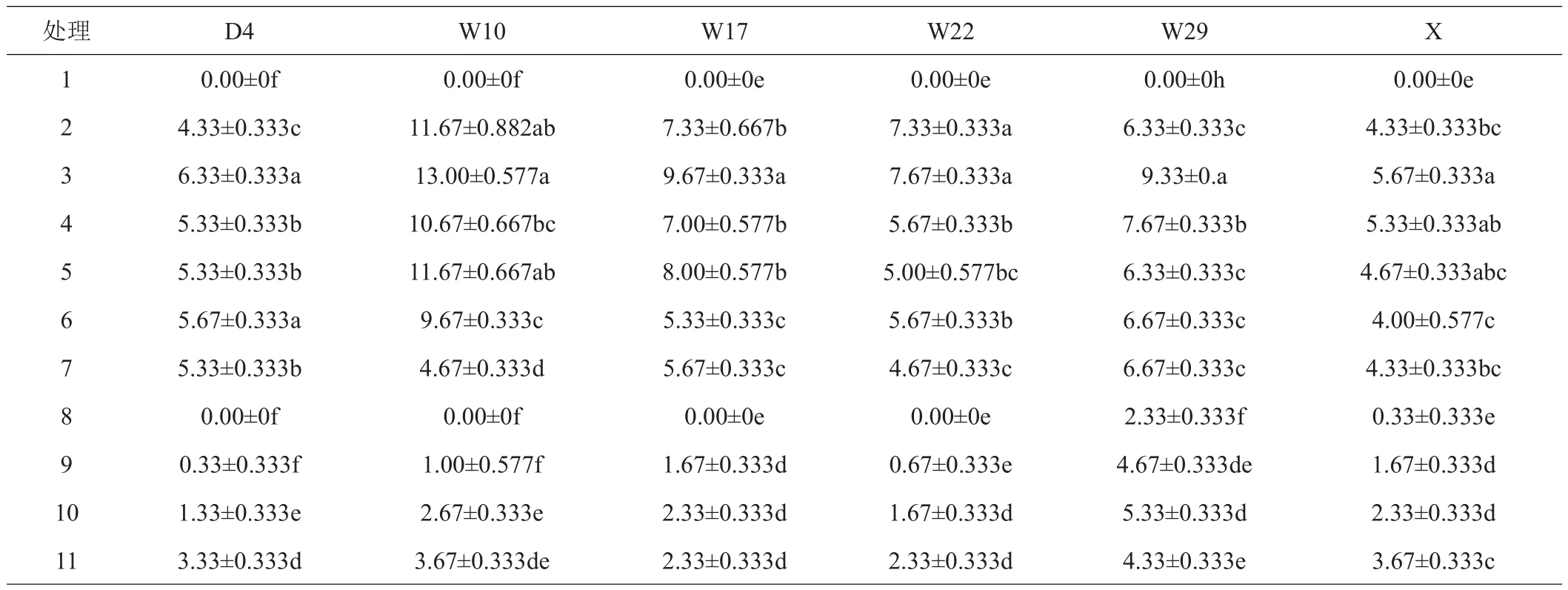
表3 不同处理对6种高山杜鹃离体叶片分化芽数量的影响 个/片
2.2 高山杜鹃离体叶片再生芽途径
对离体叶片再生过程的形态学和组织细胞学观察发现,在TDZ和ZT诱导下,W10、W29、X和 W17通过器官直接发生途径再生不定芽,而W22和D4除了器官直接发生途径外,在TDZ诱导下存在少量叶片通过愈伤组织间接再生途径分化不定芽。
高山杜鹃离体叶片由上下表皮、栅栏组织、海绵组织和维管组织组成(图2G)。器官直接再生途径中,离体叶片接入培养基后,切口处日益膨大(图2A),此时叶片上表皮细胞及其以下的数层细胞分裂活跃,细胞染色深,处于脱分化状态(图2H);培养10~13天时,这些细胞经过多次分裂,突出表皮,形成染色深的近球形的分生细胞团(图2I、J),即为叶片切口膨大处观察到透明的不同大小的球形突起(图2B);随培养时间延长球状突起长大,变绿或变红(图2C),培养约15天后,突起进一步长大、分化,可见叶片表面分化出不同发育阶段的芽点(图2D、E、K);最后芽点长成结构完整的不定芽,整个叶片上长出丛生的小植株(图2F)。
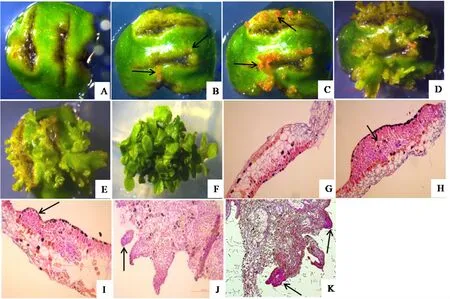
图2 0.04 mg/L TDZ诱导下,R.‘Scintillation’(W29)离体叶片直接再生芽的形态和组织细胞学观察
在离体叶片培养的初始阶段,愈伤组织间接再生途径与直接再生途径一样经历了切口处膨大、细胞分裂活跃和增殖(图3A,H),与直接途径不同的是间接途径叶片切口处持续膨大,大约培养16天时形成愈伤组织(图3B)。愈伤组织来源于叶片上表皮细胞及其以下的几层细胞的增殖,突出的愈伤组织细胞排列松散,细胞染色有深有浅(图3I)。随培养时间延长,有的愈伤组织褐化、干枯,最后死亡(图3F、G),培养约25天时,在愈伤组织细胞染色深的区域分化形成突出的细胞团(图3J),即愈伤组织上形成的突起结构(图3C),之后这些突起进一步分化、长大,形成芽(图3D、K、L),离体叶片在培养约65天后通过愈伤组织间接途径形成不定芽(图3E)。
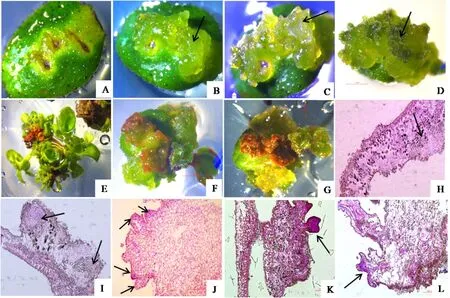
图3 0.08 mg/L TDZ诱导下R.‘Percy wiseman’(D4)离体叶片愈伤组织途径再生芽的形态和组织学观察
3 结论
TDZ诱导6种高山杜鹃离体叶片再生的效果好于ZT。TDZ浓度为0.04 mg/L时,‘Cunnigham’s White’(W10)、‘Halfdem Lem’(W17)、‘Scintillation’(W29)、‘XXL’(X)不定芽分化率达到100%,TDZ浓度分别为0.08、0.06 mg/L 时‘Percy Wiseman’(D4)和‘Madame Masson’(W22)的分化率为100%;‘Cunnigham’s White’离体叶片诱导出的不定芽最多,平均达13个/片,而‘XXL’诱导出的芽数最少,平均只有5.67个/片;离体叶片分化芽的响应时间‘Scintillation’最早,‘Cunnigham’s White’和‘XXL’稍 晚 ,其 次 是‘Halfdem Lem’,‘Madame Masson’和‘Percy Wiseman’最晚。6个基因型都可作为遗传转化的候选材料,其中‘Scintillation’、‘Halfdem Lem’和‘Cunnigham’s White’的再生能力更强。6种高山杜鹃离体叶片的再生途径主要为器官直接再生,其中‘Percy Wiseman’和‘Madame Masson’除器官直接发生途径外,在TDZ诱导下有少量叶片存在愈伤组织间接再生途径分化不定芽。研究结果为高山杜鹃离体叶片再生芽机理研究,提高其植株再生频率和遗传转化效率提供技术支持和试验基础。
4 讨论
基因型是影响植株再生的关键因子[18-19]。本研究中6种高山杜鹃离体叶片在TDZ处理下分化率均可达到100%,表明它们可作为高山杜鹃遗传转化应用的候选材料。
TDZ和ZT是杜鹃属植物组织培养中应用较多的植物激素,具有较强的细胞分裂素活性[20],但两者对杜鹃属不同种植物组培苗增殖和离体叶片分化的效应有差异[14-15]。本研究中,从离体叶片的响应时间、分化率和分化芽数看,TDZ对6种高山杜鹃离体叶片分化芽的效果均好于ZT,只是不同基因型所需的TDZ浓度有差异。而云南杜鹃、大叶常绿杜鹃的研究中又认为ZT对叶片分化的效果好于TDZ。吴晓军等[19]在小麦成熟胚再生体系中认为,胚处理方式与基因型之间可能存在着一定的互作关系,找到最适合的胚处理方法,可将基因型的再生能力最大化。在考虑到基因型间差异的同时,本研究中观察到一定浓度TDZ能诱导大量不定芽,但芽体大多短缩,最终不能形成正常的芽和植株。刘淼等[21]在牛皮杜鹃分化芽的研究中发现,TDZ对组培苗愈伤组织诱导及分化的效果显著优于ZT,但继代培养时增殖效果不如ZT。可见,在前期应用TDZ高效诱导离体叶片分化芽点的基础上,后续有必要进一步研究在培养的不同阶段组合低浓度ZT对芽点成芽的作用。
高山杜鹃离体叶片可通过器官直接或间接发生[22-23]和体细胞胚胎发生[24]2条途径再生不定芽,再生途径除与基因型有关,还与外源植物生长调节剂刺激下的内源激素水平密切相关。罗彭等[25]采用0.8 mg/L TDZ结合0.2 mg/L NAA诱导大白杜鹃、美容杜鹃、喇叭杜鹃愈伤组织,愈伤组织可分化丛芽。而本研究中,虽然6种高山杜鹃器官直接发生频率均达到100%,但‘Percy Wiseman’、‘Madame Masson’所需TDZ浓度分别为0.08、0.06 mg/L,较其他基因型高,且除器官直接发生途径外,还存在少量愈伤组织间接再生途径。说明在不同浓度外源激素诱导下,高山杜鹃再生可能存在不同途径。故后续可针对特定基因型和特定植株再生处理方法,探讨离体叶片再生不同阶段的形态和组织细胞发育动态,以提高植株再生频率和遗传转化效率。
