四川花生地方品种主要农艺性状的遗传多样性分析
2021-12-13林显凤夏友霖敬昱霖毛金雄何晋宇廖俊华毛若涵
林显凤,夏友霖,敬昱霖,毛金雄,何晋宇,廖俊华,毛若涵,游 宇
(1南充市农业科学院,四川 南充 637000;2南充市种子质量监督检验站,四川 南充 637000;3中国农业大学农学院,北京 100193)
0 引言
花生(Arachis hypogaea)是全球范围内重要的油料作物,是人们生活必须的油脂和蛋白质的重要来源[1]。中国是花生种植、加工大国,种质资源丰富[2-3]。花生新品种培育和生物技术等研究均离不开种质资源的收集和保存。为了合理利用、研究种质资源,必须先对其进行系统鉴定和评价。目前,中国已收集保存8000余份来自世界范围内不同植物学类型的花生品种资源,姜慧芳等[4-5]考察分析了其中的6390份种质资源,明确了主要农艺性状和品质特性,阐述了不同的抗病性和丰富的遗传多样性等性状,以此为基础构建了花生核心种质(包含576份材料)和小核心种质(包含298份资源)。中国山东省花生研究所和中国农业科学院油料所也先后出版了《中国花生品种资源目录》等专著[6-8]。目前国内已有多位育种工作者对当地地方品种资源开展了研究和评价,如苗利娟等[9]对河南省花生农家品种资源农艺和品质性状进行了分析,筛选到10个高油地方品种;林茂等[10]用50对SSR引物综合评价了贵州不同区域的68份地方花生品种;白冬梅等[11]开展了山西省地方花生品种芽期耐寒性鉴定和农艺性状的遗传多样性分析,鉴定出3个高耐寒品种和7个耐寒品种;卞能飞等[12]利用SSR分子标记分析了133份江苏省地方品种的遗传多样性;崔顺立等[13]证实了河北省地方花生品种具有较高的多样性程度;夏友霖、侯睿等[14-15]利用AFLP和SSR分析了川渝地区花生品种遗传多样性,发现川渝地区花生品种的多样性程度较高,而品种资源间的亲缘关系与所处地理位置关系不大,地方品种与四倍体栽培品种间亲缘关系较远。以上研究均为这些材料的开发利用和花生育种奠定了基础。
四川盆地是中国花生主产区之一,南充市农业科学院出产的“天府花生”以独特的外观和口感品质享誉中外,但花生本身的遗传特性,如远缘杂交不亲和性等原因,导致在花生新品种培育过程中难以产生新的突破性品种[16-17]。四川地方花生品种具有较好的适应性、抗逆性和变异类型,是在当地自然条件下,经过长期自然选择或人工培育驯化而成的品种,有不少品种已成为主推品种的亲本材料。研究四川省地方花生品种农艺性状遗传多样性,了解品种资源特征,对遗传改良、花生育种和花生产业可持续发展具有重要意义。目前为止,尚未见对四川省地方花生品种遗传多样性分析进行研究报道。
本研究以主要农艺性状变异、相关性和聚类等分析为基础,综合评价了南充市农业科学院花生研究所收集保存的100份四川省农家花生品种,以期明确四川地方花生品种的遗传特性和遗传多样性水平,提高地方品种的有效利用率,为遗传改良和花生育种提供理论基础和优质材料。
1 材料与方法
1.1 材料
试验材料为来自于四川省南充市、资阳市、绵阳市、内江市、德阳市、成都市、广元市和宜宾市等区域的100份花生品种,由南充市农业科学院花生研究所收集保存。品种名称及来源见表1,根据类型包括珍珠豆型(B)16份、龙生型(C)51份、普通型(D)33份。

表1 四川省供试地方花生品种名称和类型
1.2 方法
1.2.1 田间试验设计与实施 试验于2019年在南充市农业科学院花生研究所李渡镇石马滩基地进行,沙溪庙组灰棕紫泥土,中等肥力。每个品种单粒播种,种植2行,行长2 m,行距40 cm,窝距16.67 cm,3次重复,采用完全随机区组设计排列,生育期中耕除草3次,其余管理同大田试验。
1.2.2 主要农艺性状考察 成熟收获时,参照姜慧芳等编写的《花生种质资源描述规范和数据标准》[18],在每个小区内随机各取5株进行测定,并取其平均数进行统计分析。性状考察与统计包括生育期、主茎高、侧枝长、总分枝数、二次分枝数、结果枝数、单株果重、单株果数、百果重、百仁重、出仁率、荚果长宽比、种子长宽比、叶斑病和其他农艺性状。
1.2.3 数据处理 使用SPSS 19.0软件计算各农艺性状的最大值、平均值、最小值、变异系数、遗传多样性指数等。根据各性状的平均数(X)、标准差(S)将材料分为10级(从第1级X1<X-2S到第10级X10>X+2S),每0.5S为一级[9,19]。利用Shannon Wiener遗传多样性指数(H′)衡量群体遗传多样性大小。H′=-ΣPilnPi,其中,Pi为某一农艺性状第i级别内材料份数占总份数的百分比。同时,利用SPSS 19.0软件的Ward聚类法,对100份地方品种进行基于主要农艺性状的聚类分析。
2 结果与分析
2.1 农艺性状的多样性分析
统计分析参试材料14个性状的考种结果(表2),发现各农艺性状存在不同的变异,范围在0.99%~54.12%之间。其中二次分枝数的变异系数最大,为54.12%;其次是单株果重,变异幅度为36.14%,籽粒大小差异较大;生育期的变异幅度最小,为0.99%,100个品种中97个品种的生育期为121天左右,其余3个品种为128天。二次分枝数、单株果数和单株果重变异幅度较高,均大于30%,表明参试品种资源的上述3个性状具有丰富的遗传多样性。多样性指数0.19~2.99,平均为2.61,最大的为主茎高,最小为生育期。综合剖析,四川地方花生品种具有丰富的遗传多样性,在花生育种中具有较大的利用空间。

表2 参试品种农艺性状的数据统计
2.2 农艺性状的相关性分析
由表3可知,生育期与百果重、百仁重达到极显著正相关,与单株果数呈显著负相关。主茎高与结果枝数、单株果数、单株果重达到极显著负相关,与二次分枝数、百果重、百仁重和长宽比都达到极显著正相关。出仁率与主茎高、侧枝长、总分枝数、二次分枝数呈极显著负相关,与单株果数和单株果重呈极显著正相关。从供试品种14个农艺性状的相关性可以看出,多个因素影响着花生荚果产量,如生育期、结果枝数和二次分枝数等。
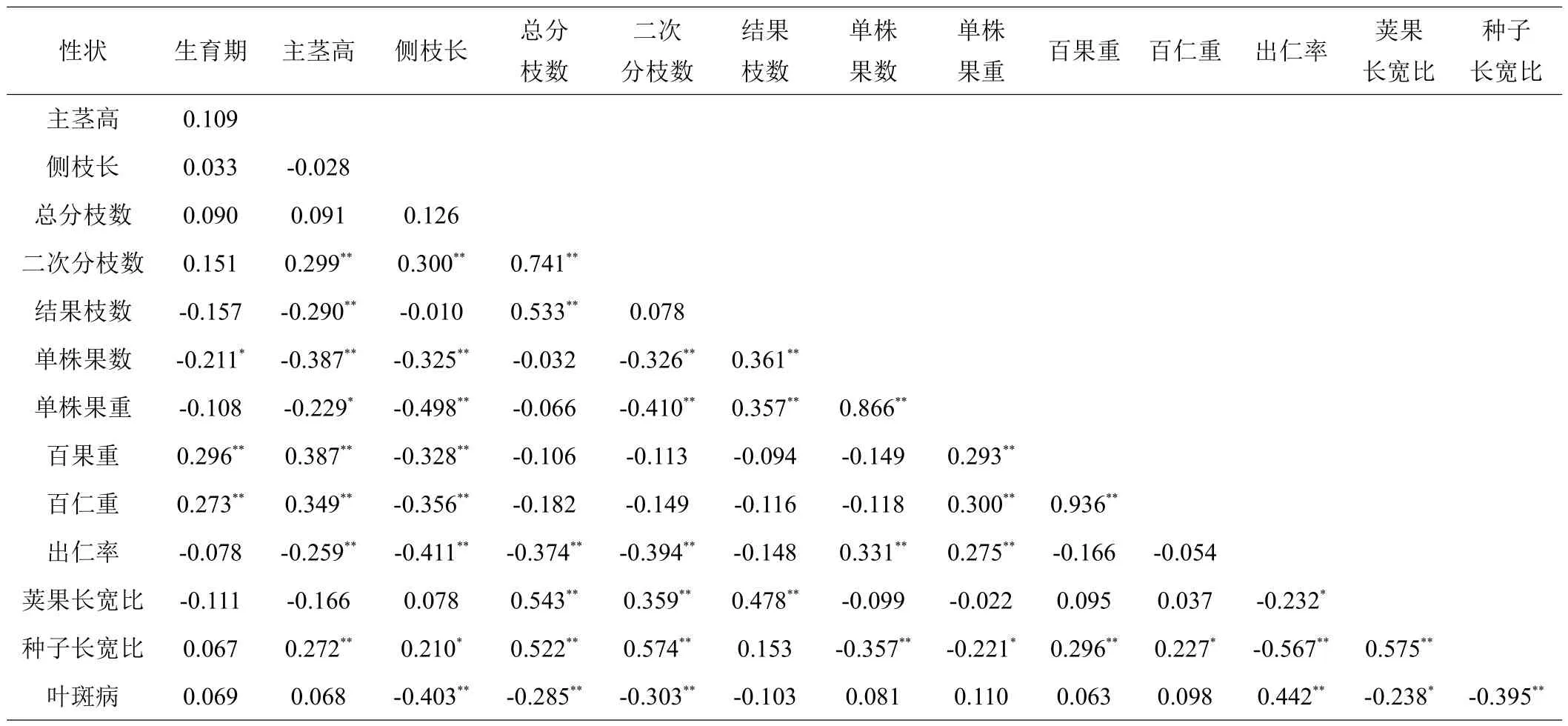
表3 地方品种农艺性状的相关性分析
2.3 农艺性状的聚类分析
以生育期、主茎高、侧枝长和总分枝数等14个农艺性状为指标对100个地方品种进行系统聚类分析。当遗传距离阈值为5时,100个参试品种分为3个类群,分别由54个品种、3个品种和43个品种组成(图1)。其中,Ⅰ类群主要为龙生型,占总数的72.22%;Ⅱ类群为普通型大果品种;Ⅲ类群普通型占48.84%,龙生型占30.23%,珍珠豆型占20.93%。

图1 四川省地方花生品种农艺性状聚类树状图
由表4可以看出,第Ⅰ类群除主茎高、单株果重、百果重、百仁重、种子长宽比和叶斑病低于其他类群外,其余性状均明显高于其他类群,说明该类群营养生长旺盛,生殖生长相对较弱;Ⅱ类群的3个品种均为普通型大果花生,生育期、主茎高、单株果重、百果重、百仁重、种子长宽比和叶斑病在3个类群中最高;第Ⅲ类群的农艺性状均居中。

表4 3个类群性状的平均表现
3 结论
本研究综合分析了四川省不同地理来源的100份花生品种农艺性状的遗传变异,从全局上明确了地方花生品种农艺性状的遗传多样性。研究表明,生育期与百果重、百仁重达到极显著正相关,与单株果数呈显著负相关;主茎高与单株果数、单株果重、结果枝数达到极显著负相关,与二次分枝数、百果重、百仁重和种子长宽比,达到极显著正相关;荚果产量受多个农艺性状的影响。聚类分析表明,在遗传距离阈值为5时,可将参试品种分为3个类群,Ⅰ类群主要为龙生型,占总数的72.22%,抗病性较好;Ⅱ类群为普通型大果品种,可作为高产育种的亲本选择;Ⅲ类群表现杂乱,普通型占48.84%、龙生型占30.23%、珍珠豆型占20.93%。同时,同一地理来源多个地方品种穿插在3个类群中,表明四川省地方花生品种的亲缘关系与地理来源和植物学类型关系不大。综上,四川省地方花生品种具有丰富的遗传多样性。
4 讨论
4.1 四川省地方花生品种农艺性状的多样性和相关性分析
四川省地方花生品种生育期绝大多数在121天左右,只有‘三台二广东’、‘仪陇大罗汉(1)-1’和‘仪陇大罗汉(1)-3’为128天。所有品种均具有较强的适应性和中等的抗病性,大多数株型属于匍匐和半匍匐型,产量低。地方品种农艺性状的变异幅度较大,平均值为18.86%,具有丰富的遗传多样性。其中,影响产量的单株果数和单株果重达到30%以上,为花生高产育种提供了较大的可能性。由相关分析可知,主茎高、百果重与百仁重呈极显著正相关,这与白冬梅、苗丽娟等[9,11]研究结果一致,而生育期与单株果重呈负相关、与单株果数呈显著负相关,这与前人研究发现的花生生育期与单株生产力的相关性小[20-21]不一致,这可能是由于2019年南充市气候极端,花生试验材料遭遇连续1个月干旱后1个月洪涝,严重影响了花生的生长和生产;而百果重和百仁重等产量性状受多个农艺性状的影响,再次说明作物的产量是由多个性状因素共同决定的[22-23],即在未来花生育种中应选择百果重、百仁重、结果枝数、单株果数和单株果重等都高的花生品种做亲本。
4.2 不同地理来源的四川省地方花生品种聚类结果比较
本研究通过聚类分析,在遗传距离阈值为5时,将供试材料划分为3个类群。类群组成与刘立峰等[24]的研究结果一致,即多粒型与普通型、大粒品种与小粒品种,甚至同一地理来源、同一类型的多个地方品种以及龙生型花生均在3个类群中交叉出现,如小粒品种‘资中小罗汉(1)’和大粒品种‘阆中大麻壳’聚在同一类群中,多粒型花生‘三台连三子’和普通型花生‘宜宾早花生’聚到同一类群中;同一地理来源的‘仪陇大罗汉’共4个材料分散于3个类群中,揭示了四川地方花生品种的亲缘关系与地域分布关系不大,这进一步说明同一植物学类型的品种亲缘关系并不是最近的。而‘金堂鸡窝’和‘资阳鸡窝’等来自不同地理区域的相同类型和名称的品种分别聚到第Ⅰ和第Ⅲ类群中,表明不同地区间同一品名的品种间具有较大的遗传差异,这极有可能是由于农民在品种流通过程中错误的售卖或者按照花生表象命名所致。本研究聚类结果显示,以生育期、主茎高等14个农艺性状为基础并不能完全区分多粒型、龙生型、普通型和珍珠豆型等不同植物学类型的地方花生品种,这与白冬梅、苗利娟等[9,11]的研究结果基本一致。
4.3 四川省花生地方品种资源的利用和创新
受地理环境和天气的影响,四川花生生育期内为高温、多雨少日照、伏旱秋涝的极端天气,严重影响花生的生长发育。研究发现四川种植面积最大的天府花生骨干亲本及其衍生品种和主推品种间遗传基础狭窄[25],因此,合理挖掘和有效利用四川省地方花生品种资源,可以改善四川花生遗传基础狭窄和抗逆性等问题,比如Ⅱ类群的3个品种均为大粒、高产花生,可以作为改良花生品种的亲本来源。本研究中所选取指标皆局限于农艺性状,还有待于考察和分析四川地方花生品种的品质性状,并结合分子水平上的分析作进一步研究。
