不同入侵植物根际土壤养分、酶活性和微生物学特征
2021-12-09彭日民彭勇向国红杨泽良
彭日民 彭勇 向国红 杨泽良

摘要:为了研究入侵植物根际土壤酶活性及土壤微生物特征,以紫茎泽兰(Ageratina adenophora)、飞机草(Eupatorium odoratum)、三叶鬼针草(Bidens pilosa)、黄顶菊(Flaveria bidentis)4种入侵植物根际土壤为对象,连续5年(2013—2017年)研究不同入侵植物根际土壤酶活性及土壤微生物特征并探讨其入侵机制。结果显示:(1)入侵植物的根际土壤pH值排序为飞机草>黄顶菊>三叶鬼针草>紫茎泽兰,其中飞机草和黄顶菊间、三叶鬼针草和紫茎泽兰间的pH值差异不显著;土壤有机碳、全氮、全钾、微生物量碳和微生物量氮含量排序为紫茎泽兰>三叶鬼针草>黄顶菊>飞机草,不同入侵植物根际土壤有机碳、全氮、全钾、微生物量碳和微生物量氮含量间的差异均显著(P<0.05),而土壤全磷、微生物量磷含量间的差异均不显著。(2)不同入侵植物土壤微生物数量以细菌最多,其次是放线菌,真菌最少;土壤细菌、真菌、放线菌数量和微生物总数量表现出一致的变化趋势,其排序为紫茎泽兰>三叶鬼针草>黄顶菊>飞机草,不同入侵植物根际土壤细菌、放线菌数量和微生物总数量的差异均显著(P<0.05)。(3)不同入侵植物根际土壤微生物丰富度指数(H)、均匀度指数(E)、碳源利用丰富度指数(S)和优势度指数(Ds)的排序为飞机草>黄顶菊>三叶鬼针草>紫茎泽兰,其中不同入侵植物根际土壤微生物群落多样性指数的差异均显著(P<0.05)。(4)不同入侵植物根际土壤纤维素酶活性、硝酸还原酶活性、酸性磷酸酶活性、蔗糖酶活性、脲酶活性和转化酶活性呈一致的变化趋势,其排序为飞机草>黄顶菊>三叶鬼针草>紫茎泽兰。(5)相关性分析和主成分分析结果表明,土壤适宜pH值的降低有利于土壤微生物繁殖,土壤有机碳、微生物量是维持土壤酶活性、土壤微生物群落多样性的关键因子。由此可知,不同入侵植物可以改变土壤微生物特性和酶活性,创造对自身生长有利的土壤环境,并借此增强其竞争能力。
关键词:入侵植物;土壤微生物量;土壤酶活性;土壤微生物群落多样性
中图分类号: S451 文献标志码: A
文章编号:1002-1302(2021)21-0217-07
收稿日期:2021-04-26
基金项目:湘中地区园林植物有害生物调查与防控技术研究(编号:湘财政〔2015〕57号)。
作者简介:彭日民(1970—),男,湖南娄底人,硕士,实验师,主要从事植物保护与病虫害防治研究。E-mail:Rimingpengg@126.com。
通信作者:杨泽良,博士,硕士生导师,主要从事植物生态学研究。E-mail:bioinfo@qq.com。
整个生态系统由多种子生态系统构成,具有明显的复杂性和多变性,尤其是对于陆地及海洋生态而言,由于分布大量动植物,加上环境的动态变化[1-2],其生态系统更为复杂,在植物的人为迁移干预下[3],形成了明显的外来物种这一独特现象。外来物种具有正面效应和负面效应两方面的效应;对于西红柿、马铃薯等作物的引进种植而言,明显改善了作物种植种类,对居民生活有益;一些花草等引入也利于增强美感;但是引入水葫芦、豚草等植被后,则形成了明显的外来物种入侵[4-5],对于当地原有生态形成了较大威胁,破坏了原来的局部生态平衡,这也成为近些年来许多生物学家研究的重点,是生态学的研究焦点之一。对于外来植物而言,由于其產生巨大的排挤性,具有较强的生长繁殖能力,加上缺乏相应的竞争物种,因此呈现较强的蔓延性,容易泛滥成灾,造成原有植被获取的养分及水分下降,进而使得生长条件进一步恶化。对于土壤生态而言,其质量的重要性集中体现在活力和肥力两大方面,此外,微生物群落分布也是对土壤状况的体现,是其中的活跃因子,对于枯枝落叶等腐殖质的降解起着关键作用,参与养分循环,制约着土壤活性,从而影响根系对养分的吸收利用,在维持植被生长发育方面起着稳定作用;根际土壤由于距离植被根系较近,成为植被吸收利用土壤养分及水分的媒介,更是微生物活动的集中区,对于多种物质和能量交换起着明显的制约效应[6-7],在促进生态平衡中起着举足轻重的作用。在外来入侵植物的影响下,原有的根际土壤状况发生了明显改善,在一定程度上促进了微生物活动,这种微环境的改变反而对外来植被生长有利,进一步促进了植被扩张。因此从这个角度来看,探究外来物种入侵与微生物关系具有明显的现实意义[4,8-9],这也是近些年来许多学者研究的焦点之一,尤其是在外来植被影响下,原有植被群落分布结构被打破[10],枯枝落叶等凋落物构成也发生了明显改变,土壤肥力也会受到影响[11-12]。除此之外,化感物质也会被外来物种释放,在这种情况下,土壤微生物生长环境发生改变,其新陈代谢等活动受到影响,进而通过微生物影响土壤活性及养分循环[5];外来物种入侵到新的区域后,由于缺乏必要的生长竞争,很容易形成较大面积的大肆繁衍,在生长竞争中能够超越原有植被体系,土壤状况及微生物活动受到影响,进而改变生物多样性的特点[6-9]。在外来植被生长的影响下,土壤的养分循环发生改变,很容易在短期内有利于外来植被生长,其竞争优势将明显超越土著植被,这将明显加速外来物种入侵,例如在千屈菜入侵影响下,湿地的宽叶香蒲群落发生了明显改变,对于土壤状况而言,不仅有机质含量发生了较大变化,而且氮的矿化速率发生了变化,形成了更为充足的氮供应[13]。此外,葱芥的入侵,导致土壤酸碱度发生改变,同时改变了氮磷等养分状况,促进了养分循环,在短期内有利于植株生长发育微环境的改善[14]。受外来植被入侵的影响,由于其短期内形成明显的扩张繁殖,很容易导致土壤养分发生巨大改变,在短期内容易出现养分下降的状况,如冰草的入侵导致土壤碳、氮含量迅速下降[15-16]。从目前的情况看,关于外来植物入侵的研究多集中在其对土壤、微生物的影响方面,对于根际土壤作用机制的研究相对较少,基于此,本研究一方面立足于紫茎泽兰、飞机草角度,另一方面立足于三叶鬼针草、黄顶菊角度来探究植被入侵对土壤酶活性的作用机制,同时对其作用于微生物的作用机制进行探究,为了增强试验的准确性,本研究开展了连续5年的实地观测分析,从而更全面地探究外来植被的综合影响,为土壤及微生物保护提供有益参考。
1 材料与方法
1.1 试验材料
本试验在湖南人文科技学院进行,试验开始于2013年,连续进行5年重复试验,紫茎泽兰、飞机草、三叶鬼针草、黄顶菊种子由湖南省农业科学院提供,籽粒饱满且大小较为接近、无虫害,首先用乙醚消毒种子,然后用蒸馏水进行冲洗,之后在日光温室内开展育苗试验。要求土质为红壤,待幼苗株高为30 cm时进行移植,要求其长势基本接近,将幼苗种植于塑料盆(直径、高度分别为25、35 cm)内,每盆种植2株。为了便于试验,每种植物种植30盆,然后将植物置于日光温室内,温度控制在25 ℃左右,湿度控制在40%~60%,每天分别在早、晚各浇1次水,要求持水量达到65%,待植株在花盆中生长1个月后进行施肥,主要施氮肥(NH4NO3)、磷肥[Ca(H2PO4)2],要求施肥质量浓度为0.55 g/L。
1.2 试验方法
采用抖落法对根际土进行取样[11-12],首先将完整根际土取出花盆,然后将大块土抖掉,并用小刀将非根际土刮下,同时借助刷子将根围5 mm内的土去掉,从而获取根际土,在此过程中注意保护根系,将不同的根际土进行混合,并分为3份:第1份直接置于速封袋内,用于后续进行含水量测试;第2份进行风干处理,对其酶活性等指标进行测定;第3份置于冰箱内,用于后续开展微生物状况的测定,土壤容重借助环刀法进行测定,并在此基础上求出孔隙度[17]。
1.3 测定方法
首先采用稀释法测定微生物数量,培养细菌、真菌、放线菌所用培养基分别为牛肉膏蛋白胨琼脂、马丁氏、高氏一号琼脂培养基。微生物量碳、氮含量的测定采用浸提法,浸提比例分别为0.38、0.45[16]。
采用比色法测定酶活性,测定的酶具有多种类型,不仅包括转化酶、纤维素酶(二者分别体现的是1 g土样在一定时间分解1 mg葡萄糖所消耗的酶量),还包括还原酶(体现的是1 g土样在 1 min 内分解1 μg NO2-所消耗的酶量)。此外,还需要对脲酶、酸酶进行相关指标的测定[17]。
在试验过程中还需要借助颜色平均变化率(average well color development,AWCD)分析方法对微生物的碳源利用状况进行分析,也能够从该角度反映微生物丰度,具体采用如下计算公式[18]:
AWCD=∑(Ci-Ri)/n;
Pi=(Ci-Ri)/∑(Ci-Ri);
H=-∑Pi(lnPi);
Ds=1-∑Pi。
式中:Ci、Ri分別Wie基孔、对照孔吸光度;Pi为吸光度的比值;n为基孔数;H为优势度指数;Ds为均匀度指数。
1.4 数据处理
在开展指标统计分析之前对数据进行相应的均值化处理,并充分考虑标准误差[19],然后用Excel 2010整理相关数据,用SPSS 21.0进行统计检验。
2 结果与分析
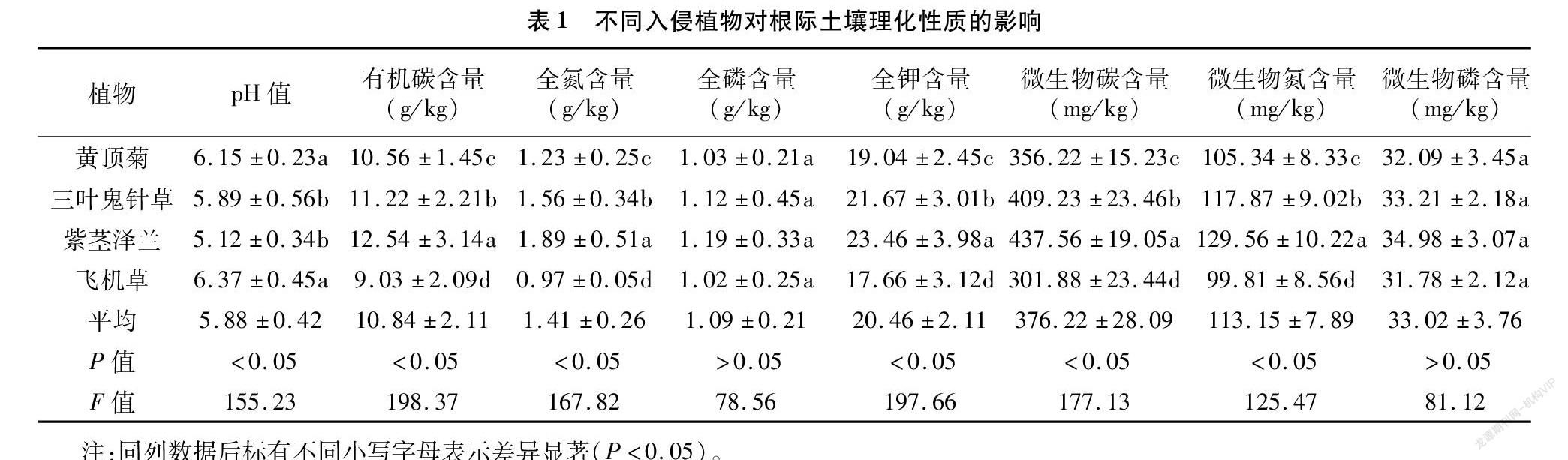
2.1 不同入侵植物对根际土壤理化性质的影响
由表1可以看出,4种入侵植物的根际土壤pH值为5.12~6.37,其排序为飞机草>黄顶菊>三叶鬼针草>紫茎泽兰,其中飞机草和黄顶菊间的差异不显著,三叶鬼针草和紫茎泽兰间的差异不显著;土壤有机碳含量为9.03~12.54 g/kg,其排序为紫茎泽兰>三叶鬼针草>黄顶菊>飞机草,不同入侵植物根际土壤有机碳含量间的差异均显著(P<0.05);土壤全氮含量为0.97~1.89 g/kg,其排序为紫茎泽兰>三叶鬼针草>黄顶菊>飞机草,不同入侵植物根际土壤全氮含量间的差异均显著(P<0.05);土壤全磷含量为1.02~1.19 g/kg,其排序为紫茎泽兰>三叶鬼针草>黄顶菊>飞机草,不同入侵植物根际土壤全氮含量间的差异均不显著;土壤全钾含量为17.66~23.46 g/kg,其排序为紫茎泽兰>三叶鬼针草>黄顶菊>飞机草,不同入侵植物根际土壤全钾含量间的差异均显著(P<0.05);土壤微生物量碳含量为301.88~437.56 mg/kg,其排序为紫茎泽兰>三叶鬼针草>黄顶菊>飞机草,不同入侵植物根际土壤微生物量碳含量间的差异均显著(P<0.05);土壤微生物量氮含量为99.81~129.56 mg/kg,其排序为紫茎泽兰>三叶鬼针草>黄顶菊>飞机草,不同入侵植物根际土壤微生物量氮含量间的差异均显著(P<0.05);土壤微生物量磷含量为31.78~34.98 mg/kg,其排序为紫茎泽兰>三叶鬼针草>黄顶菊>飞机草,不同入侵植物根际土壤微生物量磷含量间的差异均不显著。
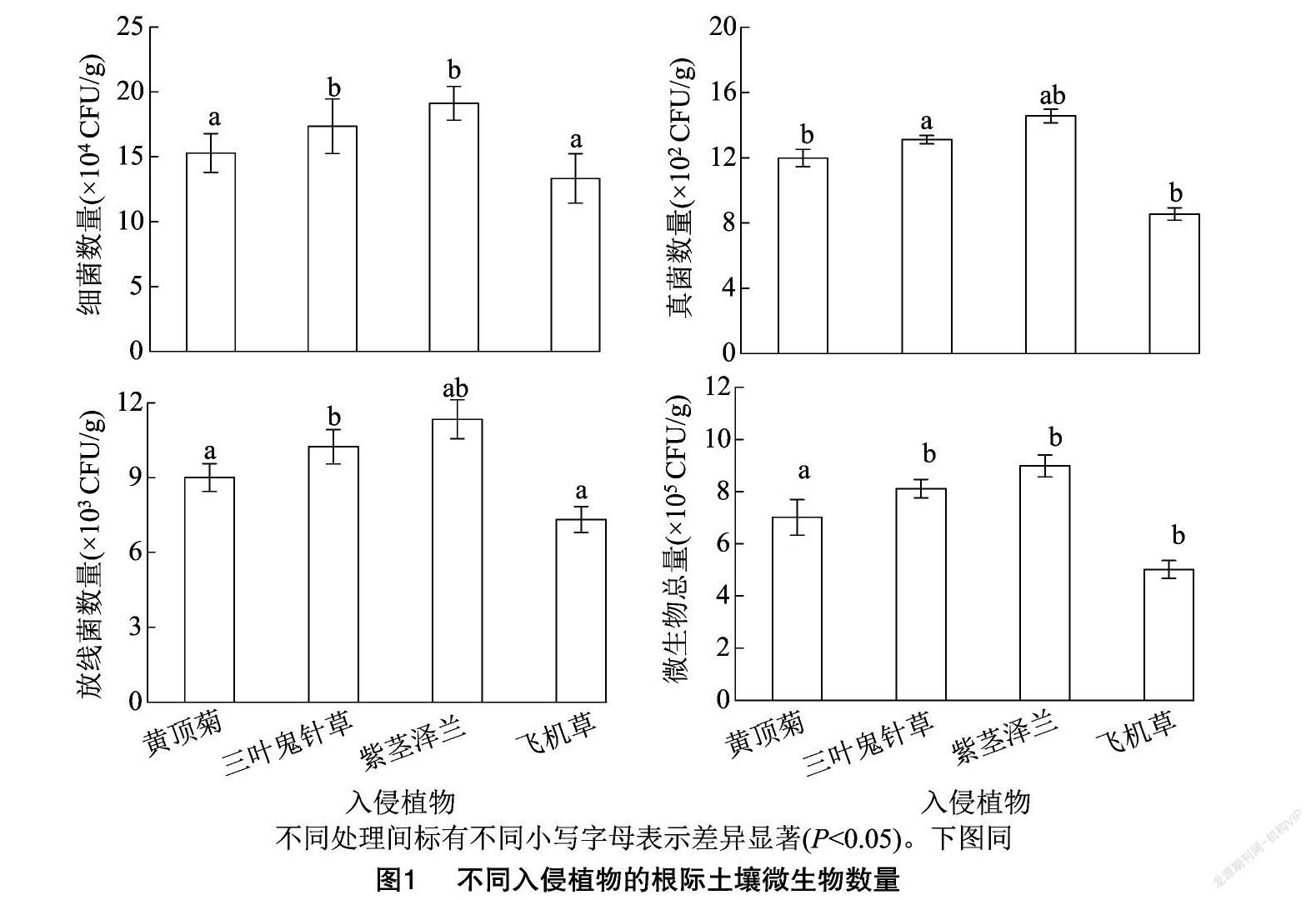
2.2 不同入侵植物对根际土壤微生物数量的影响
由图1可以看出,不同入侵植物根际土壤中的微生物数量以细菌最多,其次是放线菌,真菌数量最少;根际土壤细菌、真菌、放线菌数量和微生物总数量表现出一致的变化趋势,其排序为紫茎泽兰>三叶鬼针草>黄顶菊>飞机草,不同入侵植物根际土壤间细菌、放线菌数量和微生物总数量的差异均显著(P<0.05)。在根际土壤真菌数量上,其排序基本表现为紫茎泽兰>三叶鬼针草>黄顶菊>飞机草,飞机草和黄顶菊间的根际土壤真菌数量差异不显著,黄顶菊和三叶鬼针草间的根际土壤真菌数量差异不显著,紫茎泽兰、黄顶菊、三叶鬼针草根际土壤的真菌数量均显著高于飞机草(P<0.05)。
2.3 不同入侵植物对土壤群落多样性的影响
由图2可知,4种入侵植物根际土壤微生物丰富度指数(H)为8.67~15.42,其排序为飞机草>黄顶菊>三叶鬼针草>紫茎泽兰,且不同入侵植物间的根际土壤微生物丰富度指数差异均显著(P<0.05);土壤微生物均匀度指数(E)为0.55~0.92,其排序为飞机草>黄顶菊>三叶鬼针草>紫茎泽兰,且不同入侵植物间的根际土壤微生物均匀度指数差异均显著(P<0.05);土壤微生物碳源利用丰富度指数(S)、土壤微生物优势度指数(Ds)分别为0.51~0.78、18.78~25.11,且不同入侵植物间的根际土壤微生物碳源利用丰富度指数和土壤微生物优势度指数差异均显著(P<0.05)。
2.4 不同入侵植物对根际土壤酶活性的影响
由图3可以看出,4种入侵植物根际土壤纤维素酶、硝酸还原酶、酸性磷酸酶和蔗糖酶活性的排序为飞机草>黄顶菊>三叶鬼针草>紫茎泽兰,且不同入侵植物根际土壤纤维素酶、硝酸还原酶、酸性磷酸酶和蔗糖酶活性间的差异均显著(P<0.05);脲酶和转化酶活性的排序为飞机草>黄顶菊>三叶鬼针草>紫茎泽兰,且三叶鬼针草和紫茎泽兰间的差异不显著,二者的酶活性显著低于飞机草、黄顶菊(P<0.05)。
2.5 土壤酶活性、微生物数量与土壤理化性质的相关性
不同入侵植物土壤酶活性、微生物数量与土壤理化性质的相关性分析结果(表2)表明,土壤pH值与物种丰富度指数、碳源利用指數呈极显著负相关(P<0.01),与均匀度指数、优势度指数呈显著负相关(P<0.05);土壤有机碳、微生物量碳含量与物种丰富度指数、优势度指数、碳源利用指数呈极显著正相关(P<0.01),与均匀度指数呈显著负相关(P<0.05);细菌数量与物种丰富度指数、优势度指数呈极显著负相关(P<0.01),与均匀度指数、碳源利用指数呈显著负相关(P<0.05)。由表3可见,土壤硝酸还原酶活性与物种丰富度指数、碳源利用指数呈极其显著负相关(P<0.01),与均匀度指数、优势度指数呈显著负相关(P<0.05);土壤酸性磷酸酶活性与微生物群落多样性指数均呈极显著负相关(P<0.01);转化酶活性与碳源利用指数呈极显著正相关(P<0.01),与物种丰富度指数、均匀度指数和优势度指数呈显著正相关(P<0.05),表明土壤pH值适当降低有利于土壤微生物繁殖,土壤有机碳和微生物量是维持土壤酶活性、土壤微生物群落多样性的关键因子。
2.6 土壤酶活性、微生物数量与土壤理化性质的主成分分析
本研究以土壤酶活性、微生物数量与土壤理化性质作为原变量,通过计算变量方差和协方差矩阵的特征量,将多个变量通过降维对土壤酶活性、微生物数量与土壤理化性质进行集中和提取,识别出起主导作用的指标。由表4可以看出,第1、第2主成分特征值占总方差的比例已经大于90%,即前2个主成分对监测指标所涵盖的大部分信息进行了概括,其中第1主成分携带的信息最多,达到80%以上,第1、第2、第3主成分的累计贡献率达到95.310%,因此选取前3个因子作为主成分。表5中的土壤酶活性、微生物数量与土壤理化性质对应2个主成分的荷载,反映了主成分与变量的相关系数,载荷值大的可认为是重要因子。第1主成分负荷最高的几个指标分别为土壤pH值、有机碳、微生物量碳含量与硝酸还原酶和酸性磷酸酶活性;第2主成分最高的几个指标分别为土壤pH值、有机碳含量、微生物量碳含量、细菌数量和硝酸还原酶活性;第3主成分最高的几个指标分别为土壤pH值、微生物量碳含量、细菌数量和硝酸还原酶和酸性磷酸酶活性,与相关性分析结果相一致。
3 讨论
由本研究结果看出,虽然入侵植物种类不同,但是对于根际土壤养分的增加起着积极作用,能够促进微生物量的增加,但是对于土壤的pH值起着降低作用,对于入侵植被而言,由于其生长优势等原因,呈现大量扩散的生长态势,根系分布更为密集,这对于土壤的孔隙度起着明显的降低作用,同时还导致土壤容重下降;对于土壤质量而言,其体现指标不仅包含肥力和活性,酸碱度及微生物量等均能够体现其质量状况,在酸碱度适宜的情况下,能够显著促进植被生长发育,过高或过低的pH值都不利于植被生长,也不利于微生物的新陈代谢,一旦pH值出现较大幅度下降,很容易出现根区酸化的现象,这对于养分的吸收利用起着促进作用,会增加其酶活性,对于微生物活动起着积极作用。通过研究发现,对于4种入侵植被而言,其根际pH值均呈现降低水平,主要原因在于在入侵植被影响下,其根系在土壤中呈扩展式分布,土壤结构受此影响也发生改变,土壤养分得以提升,促进了阴阳离子的溶解,从而形成了较低的pH值。
不少学者通过实证研究发现,入侵并成功存活的植被能够在短期内吸收大量土壤养分,从而制约了原有植物对养分的吸收利用,进而破坏原有植被群落分布,打破了局地生态平衡,形成明显的生物入侵危害[18-22]。对于外来植被而言,在其入侵影响下,土壤的根际微生物分布活动受到影响[9-21],养分条件改变。通过连续5年的观测分析可知,由于入侵植被不同,土壤微生物结构也存在较大差异,其中数量最多的是细菌,而放线菌、真菌数量相对较少,这3种菌群的数量变化走势与微生物总数基本一致,对于这4种植被而言,其微生物数量最多的是紫茎泽兰,其次是三叶鬼针草、黄顶菊,飞机草的数量最少,且不同植被的根际微生物分布状况呈现显著的差异水平,并通过了0.05水平的显著性检验。从另一个角度看,不同入侵植被对微生物群落的影响也存在较大差异[21,23],但总体而言微生物数量得以明显提升。在土壤生态系统中,微生物能够有效地促进物质和能量交换,在有机质降解过程中起着关键作用,从而促进养分的转化,利于植物吸收利用。综合看来,在植被入侵过程中,无论是土壤酶活性还是微生物,都起着关键作用,受入侵植被影响,原本的土壤和植被平衡被打破,对于群落分布产生着重要影响[5,9]。微生物自身新陈代谢等活动需要从土壤中获取一定的能量,但更为关键的是微生物能够有效地分泌酶,直接在养分循环过程中发挥积极作用,从而促进土壤活性的提升[24-25]。通过对比分析得知,受入侵植被的影响,无论是还原酶、纤维素酶,还是酸性磷酸酶、脲酶等,其含量分布均体现出类似的态势,含量最高的植被区为飞机草,其次是黄顶菊、三叶鬼针草,而紫茎泽兰植被区含量最低,主要原因是对于不同的入侵植被而言,其根系特点存在较大差异,因此释放的化感物质也不同,从而影响微生物分布,进而导致不同的酶活性[4,6,23]。
通过试验对比分析得知,受入侵植被的影响,无论是微生物数量还是酶活性均呈现出明显的上升趋势,以往学者也发现了这一现象[11-12,21],虽然入侵植物种类不同,但是对于根际土壤养分的增加起着积极作用,能够促进微生物量的提高,但是对于土壤的pH值起着降低作用[11-13]。入侵植被对土壤养分的作用机制具有明显的复杂性,一方面与土壤特性及环境因子有密切关系,另一方面受到入侵时间及土著植被生长状况的影响[5-7],此外,还受到植被类型差异的制约,从而形成了一系列生物及非生物影响因素。通过相关性分析得知,土壤环境受到其理化特点的显著影响,同时一方面受到酶活性制约,另一方面受到微生物的制约,此外,微生物活动及群落分布也受到土壤理化特质的制约,只是具有不同的作用机制;此外,酶活性及微生物也能够对土壤理化特性产生制约效应,也就是说二者具有明显的相辅相成特点。综合分析可知,在不同的入侵植被影响下,土壤状况也会发生明显改变。
由于入侵的植被有明显差异,它们对微生物数量的影响效应也有差异[26-27],其中数量最多的是细菌,占比超过了90%,而真菌数量相对较少,总体而言,微生物数量有所提升。通过相关分析得知,土壤理化特性决定着微生物的生长环境,反之,酶活性及微生物也能够对土壤理化特性产生制约效应,三者之间存在密切关系。适宜的土壤pH值能够促进微生物新陈代谢等活动,促进其生长发育,进而改善土壤活性,增加酶的分泌,在改善土壤理化环境的同时改善自身生长环境。对于植被入侵的研究,仅从根际土壤方面加以探究还存在明显的局限性,应从多个角度探究其与土壤、微生物、环境因子之间的关系,通过系统性研究探究其作用机制,揭示其入侵响应机制。
参考文献:
[1]杨 星,张利辉,郑 超,等. 黄顶菊入侵对土壤微生物、土壤酶活性及土壤养分的影响[J]. 植物营养与肥料学报,2012,18(4):908-915.
[2]杨 琼,梁 羽,杨 剑,等. 入侵植物薇甘菊的根际土壤微生物特征[J]. 生态科学,2015,34(2):148-155.
[3]向 丹,陈保冬,李 欢,等. 入侵植物光梗蒺藜草对土著丛枝菌根真菌群落的影响[J]. 作物研究,2015,29(5):534-541.
[4]付 伟,王 宁,庞 芳,等. 土壤微生物与植物入侵:研究现状与展望[J]. 生物多样性,2017,25(12):1295-1302.
[5]胡文武,辛国荣,郭健桦,等. 广州地区7种菊科入侵植物丛枝菌根侵染和根际土壤孢子密度的生境差异性分析[J]. 热带亚热带植物学报,2015,23(2):183-190.
[6]焦润安,李朝周,刘高顺,等. 生草对油橄榄根际土壤丛枝菌根真菌多样性的影响[J]. 生态与农村环境学报,2019,35(4):459-468.
[7]闫 静,张晓亚,陈 雪,等. 三叶鬼针草与不同本地植物竞争对土壤 微生物和土壤养分的影响[J]. 生物多样性,2016,24(12):1381-1389.
[8]宋 振,纪巧凤,付卫东,等. 黄顶菊入侵对土壤中主要功能细菌的影响[J]. 应用生态学报,2016,27(8):2636-2644.
[9]梁作盼,李立青,万方浩,等. 土壤微生物对紫茎泽兰生长与竞争的反馈:不同灭菌方法的比较[J]. 中国生态农业学报,2016,24(9):1223-1230.
[10]邓丹丹,刘 棋,蒋智林,等. 紫茎泽兰与不同植物群落土壤养分及酶活性差异[J]. 生态环境学报,2015,24(9):1466-1471.
[11]周 婕,苗一方,方 楷,等. 紫茎泽兰内生真菌及其根际土壤真菌的多样性研究[J]. 生态科学,2019,38(5):1-7.
[12]邰凤姣,朱珣之,韩彩霞,等. 外来入侵植物意大利苍耳对土壤微生物群落、土壤酶活性和土壤养分的影响[J]. 生态科学,2016,35(4):71-78.
[13]阿的鲁骥,何 兵,王长庭,等. 入侵植物土荆芥对川西北高寒草甸3种培育牧草根际土壤酶活性、微生物数量及土壤养分的影响[J]. 西南农业学报,2015,28(2):815-821.
[14]尚春琼,朱珣之. 外来植物三叶鬼针草的入侵机制及其防治与利用[J]. 草业科学,2019,36(1):47-60.
[15]殷吉林,张晓亚,闫 静,等. AMF对入侵植物黄顶菊与棉花竞争生长的反馈作用[J]. 生态环境学报,2015,24(7):1132-1136.
[16]王 月,张玉曼,李 乔,等. 黄顶菊入侵域不同土层土壤微生物群落结构的比较[J]. 河北农业大学学报,2016,39(1):35-42.
[17]闫雅楠,叶小齐,吴 明,等. 入侵植物加拿大一枝黄花根际解钾菌多样性及解钾活性[J]. 植物生态学报,2019,43(6):543-556.
[18]张泰劼,崔 烨,郭文磊,等. 外来植物阔叶丰花草的研究进展[J]. 杂草学报,2019,37(3):1-5.
[19]田秋阳,周鸿章,鲁 萍. 外来杂草反枝苋对大豆根际土壤微生物碳源利用和土壤理化性质的影响[J]. 作物杂志,2018,28(2):24-30.
[20]肖生鸿,刘锴栋,刘晚苟,等. 广东徐闻沿海滩涂红树林保护区入侵植物现状及防治策略[J]. 杂草学报,2019,37(4):22-30.
[21]王桔红,张丽娜,陈学林,等. 入侵植物对根际土壤微生物群落影响的研究进展[J]. 生态科学,2016,35(6):204-210.
[22]罗 欢,覃丽婷,刘可丹,等. 南宁市园林绿地系统外来入侵植物物种调查与分析[J]. 杂草学报,2019,37(4):31-36.
[23]陈学林,张丽娜,王桔红,等. 红毛草入侵程度对根际土壤微生物的影响[J]. 西北农林科技大学学报(自然科学版),2017,45(4):165-172.
[24]毛 璐,宋 春,徐 敏,等. 栽培模式及施肥对玉米和大豆根际土壤磷素有效性的影响[J]. 中国生态农业学报,2015,23(12):1502-1510.
[25]康小武,代婷婷. 马缨丹入侵对根际和非根际土壤理化性质的影响[J]. 现代农业科技,2016(20):84-86.
[26]管 铭,潘小翠,张崇邦,等. 受外来植物入侵的土壤微生物生态学研究进展[J]. 浙江大学学报 (理學版),2015,42(4):445-452.
[27]郑 洁,刘金福,吴则焰,等. 闽江河口红树林土壤微生物群落对互花米草入侵的响应[J]. 生态学报,2017,37(21):1502-1509.
