昌宁县中华蜜蜂种群遗传结构调查
———基于线粒体DNA COX1 分子标记
2021-12-09田虎兴李贵安普绍忠普光正
邱 勇,田虎兴,李贵安,曾 冠,普绍忠,普光正
(1.云南农业大学动物科学技术学院,云南 昆明 650201;2.昌宁县翁堵普家寨中蜂养殖专业合作社,云南 昌宁 678115)
云南省地处我国西南边陲,地形地貌复杂,气候类型多样,生物物种丰富,素有动植物王国之美誉。云南也是世界上蜜蜂遗传资源最丰富的地区之一,目前已发现的9 个蜜蜂(Apis) 种中有6 个在云南都有分布[1],被称为中国的“蜜蜂资源基因库”。尤其中华蜜蜂不仅种群数量居全国之首,其遗传多样性丰富度也首屈一指。根据杨冠煌等[2]于20 世纪70-80 年代开展的全国中蜂资源调查结果显示:云南省境内分布有云贵高原、滇南和西藏3 种中蜂生态类型。殷玲等[3]及冯毅楠等[4]分别利用微卫星DNA 和线粒体DNA分子标记对云南省6 个中华蜜蜂种群实施的遗传多样性分析发现:云南不同地理种群间的基因交流较少,相互间存在极显著的遗传分化。这些中蜂资源对于蜜蜂生物学、生态学、进化遗传学等方面的研究及后续的保种育种、商业利用等具有很大价值。与极高的资源禀赋不相匹配的是云南各地区对当地中蜂资源的挖掘、保护及利用相对滞后和盲目无序。为了能够科学规划并有序开展这些工作,有必要首先对当地中蜂资源实施多样性分析及种群遗传结构调查。
保山市昌宁县位于云南省西部,地处北纬20°14′至25°12′、东经99°16′至100°12′[5]。县域内低热、温热、温凉、高寒气候均有分布,很好地呈现了云南“一山分四季,十里不同天”的地理和生态特点。全县植物资源丰富,拥有发展养蜂业的得天独厚之条件。近些年来,尤其是全面实施“国家现代蜂产业技术体系建设”和“脱贫攻坚”行动以来,昌宁县积极推动中蜂产业发展,在一些植被丰富但耕地较少的山区、半山区大力拓展中蜂养殖规模。2019-2020 年,我们对当地的中华蜜蜂实施了种群遗传结构调查。
动物细胞内的线粒体DNA(mtDNA)属核外遗传物质,具有分子量较小、基因组结构简单、不发生重组、种内变异丰富等特点。此外线粒体DNA 遵循母系遗传,即同一母本的后代具有完全相同的mtDNA。对于蜜蜂而言,通常一个蜂群中的所有个体都是同一蜂王的后代,因此1只工蜂的mtDNA 就能代表同群所有个体的。这些特点使mtDNA 非常适宜于对蜜蜂进行遗传多样性分析和种群遗传结构调查。本文以mtDNA上的细胞色素氧化酶亚基1 基因(COX-1)片段为分子标记,对昌宁县4 个采样点的96 群中华蜜蜂实施群体检测并利用相应的生物信息学软件进行数据分析。同时我们将本次实验所得数据与云南省其他地区的6 个中华蜜蜂种群的数据进行了对比性综合分析,研究结果可为准确定位、合理布局昌宁县中蜂养殖业提供基础数据。
1 材料与方法
1.1 样品采集
本实验中的中华蜜蜂样本于2019 年采集自云南省保山市昌宁县的4 个采集点,每个采样点采集10~30 群,共96 群蜜蜂样本,样本详细信息见表1 和图1。采集时直接从蜂巢中抓取30~50 只工蜂,即刻置于75%的乙醇中并尽快带回实验室保存至-75℃超低温冰箱中备用。针对每群蜜蜂样本随机选取1 只工蜂,利用全基因组DNA 提取试剂盒[天根生化科技(北京) 有限公司]提取全基因组DNA。
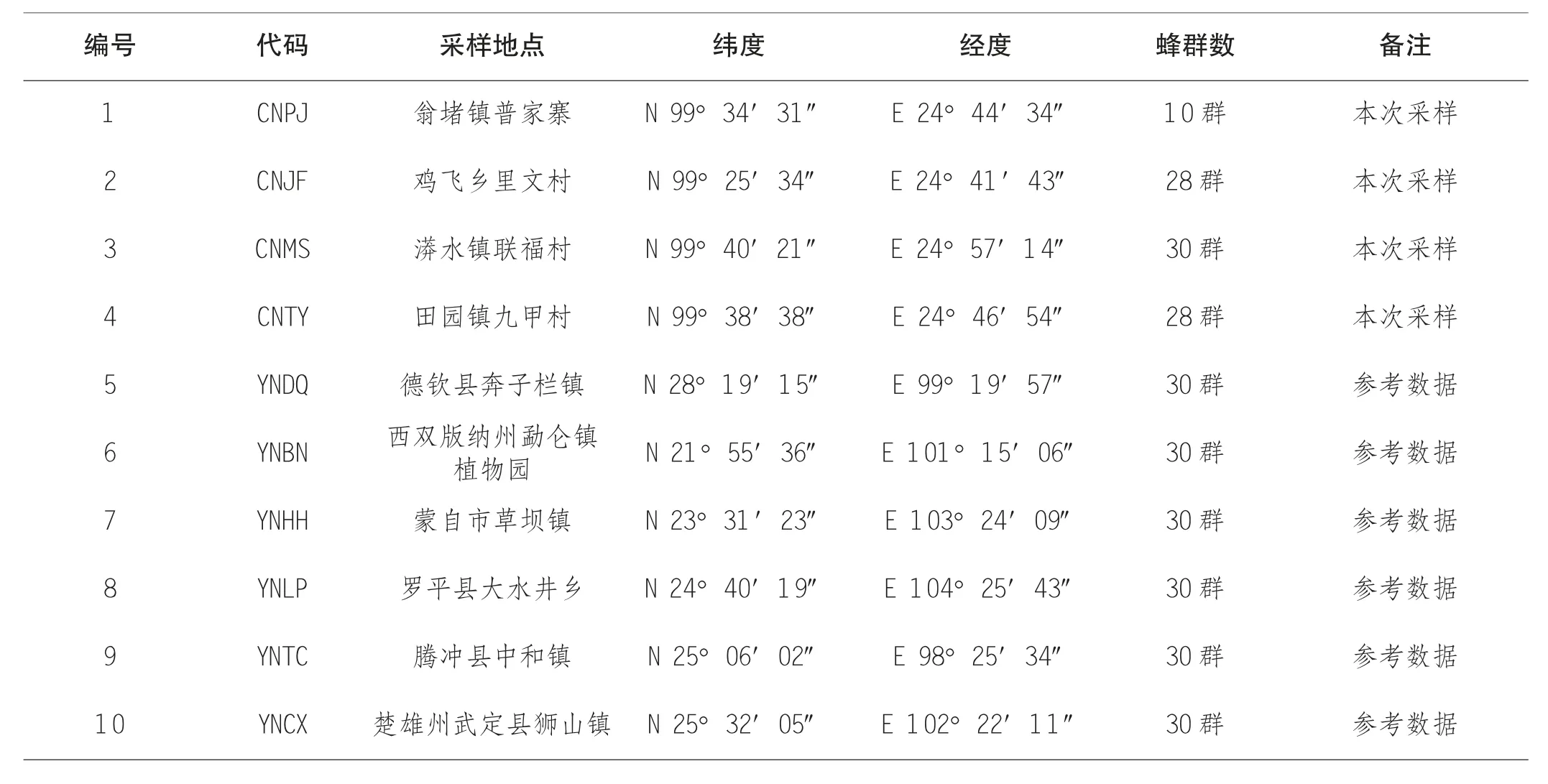
表1 中华蜜蜂样本采集点地理信息
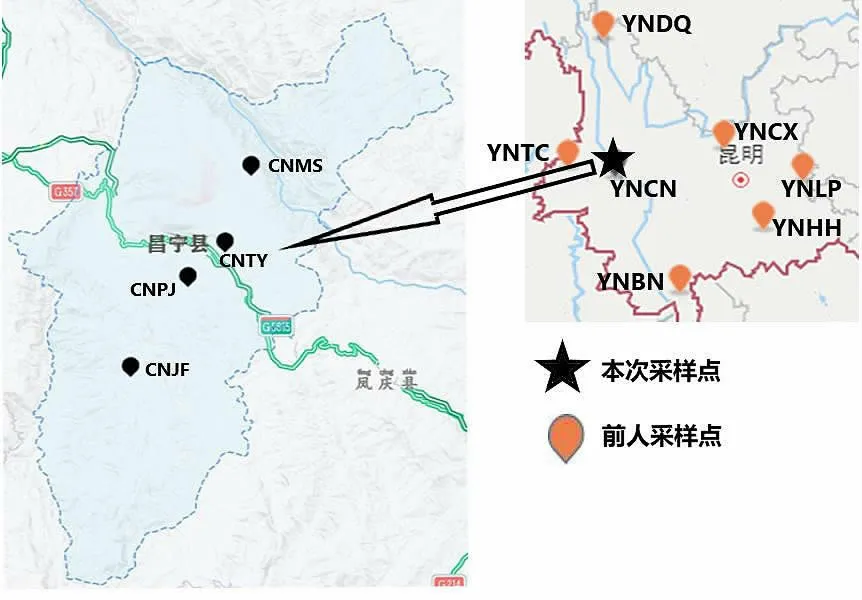
图1 中华蜜蜂采样地理位置
1.2 PCR 扩增及测序
根据已发表的中华蜜蜂线粒体基因组(NCBI 登录号:NC_014295) 为模板,设计引物、正向引物COX1-F 和反向引物COX1-R的目标片段为细胞色素C 氧化酶亚基1 基因部分序列,其中上游引物序列为5’-CTCCAGATATAGCATTTCCTCG-3’,下游引物序列为5’-TGCAAATACTGCTCCTATTGA-3’引物由生工生物工程(上海)股份有限公司合成。PCR反应总体系为30μL,其中DNA 样品6μL,上游引物3μL,下游引物3μL,2×Tap PCR Mix 18μL,扩增程序为: 95℃预变性5 min,94℃变性30 s,58℃复性30 s,72℃延伸30 s,运行36 个循环,72℃延伸5 min,4℃保存。所得产物经1%琼脂糖凝胶电泳检测后(图2),目标条带清晰,交由生工生物工程(上海)股份有限公司进行双向测序。

图2 经琼脂糖凝胶电泳检测后,目标条带清晰
1.3 样本序列处理及分析方法
首先利用SeqMan 软件对测序结果进行正、反向序列蜂图检查,校对修剪并生成一致序列。校对修剪后序列长度为794bp,由MEGA 6.06 打开对一致序列进行Alignment 计算,并用ClustalW 排序分析序列碱基组成。用DnaSP5.1 软件包计算单倍型种类、频率及单倍型多样度(Hd) 、核苷酸多样度(π);利用Arlequin3.5 软件包进行Tajima’s D 中性检验(表2) 和分子方差分析(AMOVA) (表4);使用Network5.03 软件(http://www.fluxus-engineering.com)中的Median-Joining 算法对本实验中鉴别出及前人研究数据发现的单倍型进行网络关系构建。
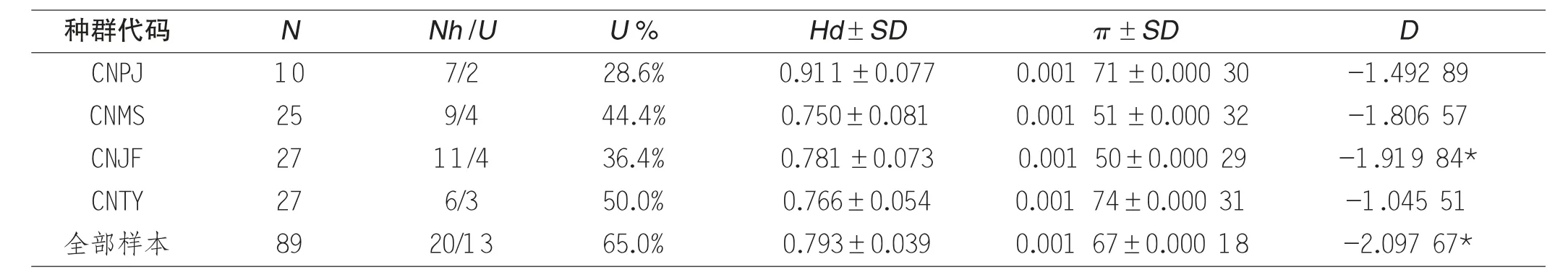
表2 云南省昌宁县中华蜜蜂各地理种群遗传结构指标分析结果
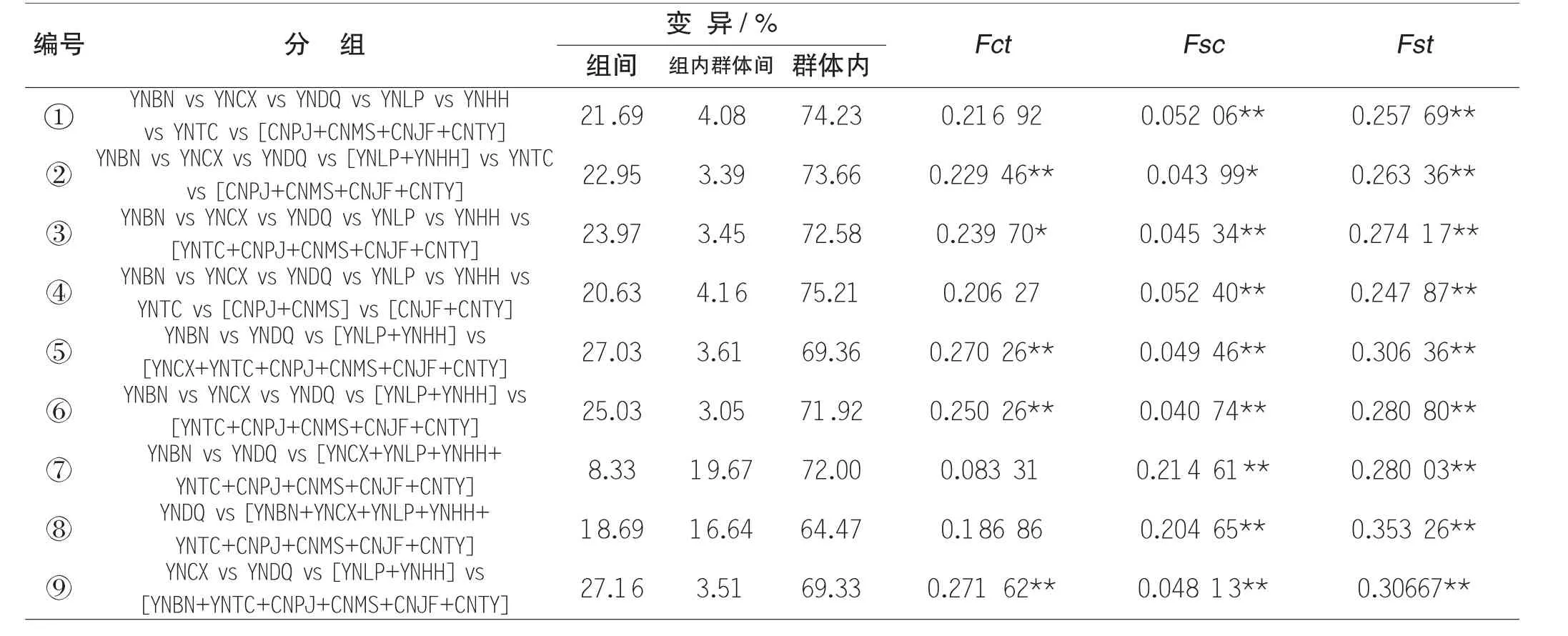
表4 分子方差分析结果
2 结果分析
2.1 序列遗传变异分析
针对4 个采样点的96 只中华蜜蜂样本,共获得89 条可用的线粒体DNA 序列。对每条序列进行校对修剪后序列长度为794bp,经过与NCBI 数据库中已发表序列进行比对后确定为目标序列COX1 片段。序列碱基组成的A+T 含量为74.3%,表现出显著的A+T偏倚性。共发现23 个变异位点,其中转换变异位点21 个,颠换变异位点2 个。与华南地区中华蜜蜂参考序列(CH1-CH35)(NCBI 登录号:KC175488-KC175522)合并分析时同源片段为794 bp,序列对比发现52 个变异位点,形成了51 种单倍型,按照(CH+ 数字) 规则依次命名。其中4 种单 倍型(CH1,CH20,CH24,CH25) 在前人对中华蜜蜂的研究中已有报道,另外16 种单倍(CH36-CH51) 型为本文首次报道,将这51 种单倍型的变异位点如(表3) 所示。

表3 51 种单倍型的变异位点
将已获得的昌宁地区89 条一致基因序列与赵文正等[6]中所得到的云南省西双版纳、迪庆、楚雄、红河、罗平、腾冲等6 地中华蜜蜂线粒体DNA COX1 区间的基因序列及参考序列(CH1-CH35)进行处理生成一致序列同源片段为794 bp。形成了74 种单倍型,按照(CH+ 数字) 规则依次命名,其中27 种单倍型(CH1,CH20,CH24,CH25,CH35,CH52-CH74)在前人对中华蜜蜂的研究中已有报道。
2.2 种群内遗传多样性分析
在昌宁县4 个地理种群中,均发现一定比例(28.6%~50%) 的独有单倍型,全部样本的独有单倍型率达到65%,表明昌宁县的4个地理种群的遗传结构独特性较为显著。CNTY 群体的独有单倍型率达到50.0%,表明该群体与其它种群的基因交流相对较少。本次实验中全部样本的Hd值为0.793,表明昌宁地区的中华蜜蜂种群具有相对丰富的遗传多样性,其中CNPJ 群体的Hd值最高(0.911),C NMS、CNJF、CNTY 等3 个地理种群的Hd值均在0.7 到0.8 之间。核苷酸多样度(π)是指从群体中随机抽取的两条序列之间平均每个位点上的核苷酸差异数,常用于度量种群内部存在的遗传差异[7]。一个群体内包含的相互间序列差异越大的个体越多,则π值越高,反之π值越低。4 个地理种群的π值在0.001 50~0.001 74 之间波动,表明了昌宁地区的中华蜜蜂群体存在一定的遗传分化,但是4 个地理种群内部的遗传差异化程度不大,实验样本总π值为0.001 67 均比前人研究中所报道的云南省6 个地理种群除西双版纳群体以外(迪庆、腾冲、楚雄、罗平) 的π值高,推测昌宁地区可能蕴含着比迪庆、腾冲、楚雄、罗平、红河等5 个地区更丰富的遗传多样性。Tajima D 值是对微进化事件进行中性检验时表征显著性的度量值,Tajima D 呈显著正值时推测被检验群体近期经历过种群缩减(瓶颈效应);Tajima D 呈显著负值时推测被检验群体近期经历过种群扩张或定向选择。本实验中,总样本的Tajima D 值达到负值显著水平,表明了该地区的中华蜜蜂群体近期可能经历过一定程度的种群扩张。
2.3 种群遗传结构分析
为了进一步探究昌宁县中华蜜蜂地理种群在云南省中华蜜蜂种群遗传结构中所处位置及“扮演”的角色,我们将前人研究中云南省6个中华蜜蜂地理种群(迪庆、腾冲、楚雄、罗平、红河、西双版纳) 及华南地区中华蜜蜂的线粒体DNA 数据纳入本文,分别进行了分子方差分析(AMOVA) 和单倍型进化网络关系(Network) 图构建。针对10 个地理种群进行了9 次分子方差分析,结果表明:无论按照何种分组方式,群体内变异对总变异的贡献率最大(≥64.4%),表明云南省不同地理种群内部所蕴含的遗传多样性丰富度都较高。其中按照9 号分组方式的组间变异对总变异的贡献率相对最高(27.03%) 且组间变异指数、组内群体变异指数、群体内变异指数均达到极显著水平,表明按第9 号的分组方式来划分这10个地理种群的遗传类群相对最合理。
单倍型进化网络关系(Network) 见图2。由图2 可知:单倍型网络关系图呈星状分布,CH1 单倍型为优势单倍型,其余单倍型均与CH1 呈衍生关系且出现频率较低,呈现出典型的辐射演化型网络关系,推测云南省及华南地区的中华蜜蜂种群来自同一祖先。云南省各地理种群间共享单倍型数为10,占本区域发现全部单倍型数量的22.7%,表明云南省中华蜜蜂种群间近期的基因交流相对较少,各地理种群在近期经历了相对独立的演化过程。
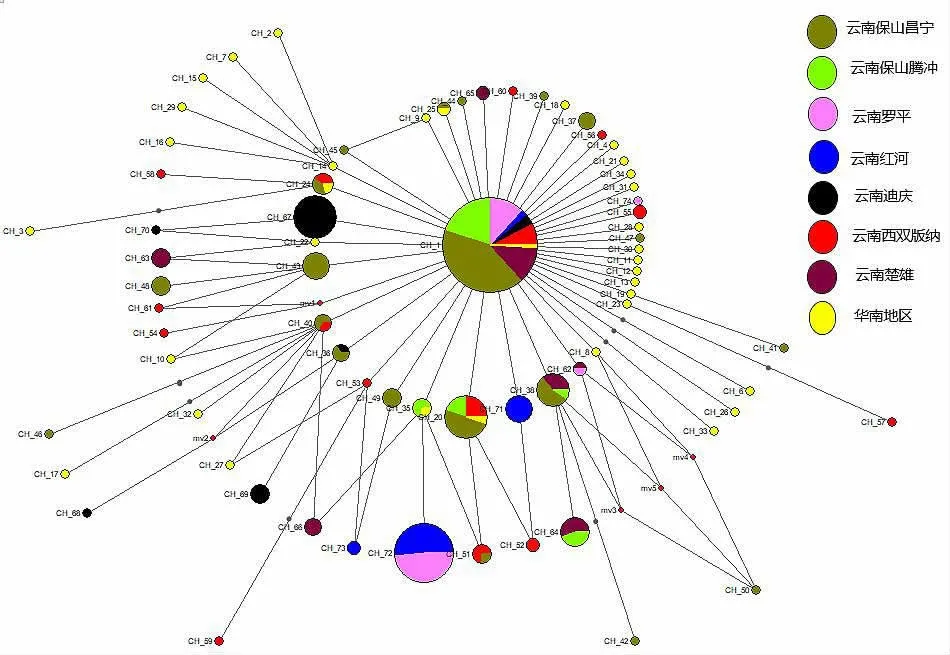
图3 云南省地区中华蜜蜂单倍型网络关系
Fct组间变异固定指数;Fsc组内群体间变异固定指数;Fst群体内变异固定指数*P≤0.05, **P≤0.01
3 讨论
由表2 来看,CNTY 群体与昌宁地区其他群体基因交流较少,所处的地理位置海拔普遍要低于其余3 个地理种群,但徐浩等[8]通过对湖南龙山不同海拔中华蜜蜂种群的遗传多样性进行分析后得出海拔因素可能不直接影响中华蜜蜂的种群遗传多样性的结论。提示可能是由于CNTY 群体所处的地理位置靠近公路,周边交通便利,一定程度上人为因素阻隔了种群间的交流。
从云南省地区单倍型网络关系图来看,表现出云南省丰富的遗传多样性和较高的单倍型多样性。其中保山昌宁的群体中表现出了与迪庆地区群体之间的群体的基因交流趋势,但迪庆地区的群体与保山市腾冲地区的群体并未表现出基因交流趋势,说明了昌宁的群体通过某种渠道与迪庆地区群体产生了交流,表明了迪庆类群出现了向外的种群扩张趋势。同时,昌宁的群体与其余(除迪庆外) 的5 个地区群体之间的基因交流较少,但也存在6 种共享的单倍型,说明这些地区的中华蜜蜂与保山昌宁的群体间也存在着有限的基因交流,推测保山昌宁的中华蜜蜂群体是迪庆地区中华蜜蜂群体向外扩张的过渡群体,这与李华等[9]通过对云南省中西部中华蜜蜂的形态特征分析后所得出的云南省中西部的中华蜜蜂群体是云南省境内两大中华蜜蜂类群的过渡带的结论相吻合,提示着昌宁地区的种群很可能具有与迪庆地区种群相似的生物学特性。刘建等[10]通过对云南省地区中华蜜蜂进行形态学分析后得出迪庆地区的种群拥有比西双版纳种群更强的飞行能力,推测昌宁地区的种群也应具有相似的特性。同时单倍型网络关系图显示CH20 单倍型为西双版纳、昌宁、腾冲地区种群所共享,CH24、CH40、CH51 为西双版纳与昌宁地区所共享,表明西双版纳、昌宁、腾冲地区种群的遗传差异相似,这与分子方差分析结果中的第9 号分组方式相吻合,同时也与殷玲等[11]所得出的腾冲群体与西双版纳群体聚为一类的结论相吻合。从地理位置上看,西双版纳位于滇南地区,腾冲、昌宁位于滇西南部,地理位置相距较远,一般认为,距离隔离或地理条件造成的隔离,使地理种群间基因流动受到限制,会促进地理种群间遗传差异的形成[12]。但是本实验与殷玲等[11]的结果均表明在西双版纳与腾冲、昌宁地区的种群似乎不遵循这一结论,而西双版纳的种群却与其地理位置较近的红河地区种群形成了较明显的遗传差异,推测自然地理因素并不是影响昌宁地区种群遗传结构的决定因素,可能是人为活动影响的结果,比如人为饲养(转地,买卖) 等造成了昌宁地区种群与西双版纳种群间的基因交流。董霞等[13]和殷玲等[3]也提出过各地中华蜜蜂群体的交流可以通过蜂王的流通实现,大量的人工选择可能会导致各地理种群的遗传多样性减小,而遗传多样性降低会影响蜂群的生存能力[3],对此我们建议应立即对昌宁地区的中华蜜蜂群体进行保护,避免盲目地引进外地蜂群。鉴于昌宁地区的种群很有可能是迪庆地区种群向外扩张的窗口,因此建立昌宁中华蜜蜂自然保护区也能很好地保护迪庆地区的种群。
