绿肥配施化肥对岩溶区水稻土壤细菌群落结构的影响
2021-12-05钟菊新唐红琴李忠义董文斌韦彩会何铁光
钟菊新,唐红琴,李忠义*,董文斌,韦彩会,李 强,何铁光*
(1 中国地质科学院岩溶地质研究所/自然资源部广西岩溶动力学重点实验室, 广西桂林 541004;2 联合国教科文组织国际岩溶研究中心, 广西桂林 541004;3 广西农业科学院农业资源与环境研究所, 广西南宁 530007)
我国南方岩溶生态系统脆弱,面临水土流失、生态功能衰退和环境污染等问题[1]。石灰性水稻土作为典型岩溶土壤,具有低磷和低钾等特性[2],农业生产过程中对化肥需求量较大,而过量化肥投入将会导致生态环境逐渐恶化,需要积极寻求化肥替代品。绿肥种植作为一种高效、可持续发展的农业生产技术策略,在提高土壤肥力、改善土壤质量、防止水土流失等方面起着重要的作用[3]。研究表明,种植绿肥能有效促进稻田土壤有机质积累[4],提高土壤氮素利用效率[5],改善土壤养分状况。同时,冬种绿肥在减施化肥20%~40%的条件下可保证水稻品质和产量稳定[6]。可见,绿肥在促进土壤养分循环和提高作物产量中发挥着重要的作用。
研究冬种绿肥作用下的土壤微生物群落特征有助于阐释其提升土壤肥力和作物生长的内在机制。研究表明,与单施化肥相比,绿肥配施化肥能显著改变红壤稻田土壤细菌多样性,提高土壤细菌总量及固氮、溶磷等多种功能菌属相对丰度[7-8]。长期绿肥施用能促使特定功能菌属的形成,在土壤碳、氮等元素循环过程中发挥重要作用。然而,由于岩溶区石灰性水稻土养分有效性低,在绿肥施用条件下,土壤养分状态、土壤细菌群落特征及其潜在的交互关系尚不明确。因此,以活跃在岩溶区棕色石灰性水稻土的土壤细菌为研究对象,探讨绿肥作用下的土壤养分状况、土壤细菌群落特征及其潜在的交互关系,以期为岩溶区绿肥替代化肥提供理论依据和数据支撑。
1 材料和方法
1.1 研究区概况
试验区位于广西南宁市那桐镇定典村 (23°0′41′N、107°51′21′E),该地属亚热带季风气候,年平均气温21.6℃,年均降水量达1300 mm,平均海拔64 m。土壤类型为典型的岩溶区棕黄色水稻土,成土母质为白云岩风化物。2017年试验开始前耕层(0—20 cm)土壤基础理化性状为: pH 7.03、有机质 37 g/kg、全氮 5.66 g/kg、有效氮 158 mg/kg、速效磷 11.7 mg/kg、速效钾 86 mg/kg。
1.2 试验设计
试验开始于2017年10月,种植模式为绿肥–早稻–晚稻连作模式,3月份正值绿肥盛花期,将绿肥翻压后开始种植早稻,7月底收割早稻,8月份种植晚稻,11月收割晚稻后播种绿肥。试验设3个处理,分别为单施化肥(CK)、单种绿肥(MV)和绿肥配施化肥(MF)。供试绿肥品种为紫云英(桂紫7号),播种量为 30 kg/hm2,翻压量为 22500 kg/hm2,单季施肥量分别为 N195 kg/hm2、P2O590 kg/hm2、K2O 120 kg/hm2,肥料种类为尿素 (含 N 46.4%)、过磷酸钙 (含 P2O518.0%)和氯化钾 (含 K2O 60%),其中各阶段的基肥∶分蘖肥∶攻穗肥均按4∶3∶3比例施入稻田。试验区每个小区面积为16.5 m2,每个处理3次重复,小区与小区之间用田埂分开,采用随机区组排列。
1.3 样品采集
试验土壤样品采于2020年7月,按照5点采样法对0—20 cm耕作层土壤取样,5点混成一个样品,共9个土样,装入自封袋运回实验室。去除根系等杂物后,用四分法取10 g左右的鲜土送测序公司,进行细菌群落结构分析测定;另一部分自然风干后进行土壤理化性质测定。
1.4 样品测定
1.4.1 土壤理化性质 土壤pH采用电位法(水∶土= 2.5∶1,质量体积比)测定;土壤有机质采用重铬酸钾–硫酸氧化法测定;全氮采用全自动凯氏定氮仪测定;全磷采用碱熔—钼锑抗比色法测定;碱解氮以硫酸亚铁为还原剂,采用碱解氮扩散法测定;速效磷采用碳酸氢钠浸提—钼锑抗比色法测定;速效钾(available potassium,AK)采用乙酸铵交换—火焰光度计法测定;交换性钙(exchangeable calcium,E-Ca2+)和交换性镁 (exchangeable magnesium,E-Mg2+)采用乙酸铵交换—原子吸收分光光度计法测定。具体分析方法详见鲁如坤的《土壤农业化学分析方法》[9]。
1.4.2 土壤细菌 采用特异性引物 515F (5′–GTGC CAGCMGCCGCGGTAA–3′)和 970R (5′–CCGTCAA TTCMTTTRAGTTT–3′)对土壤细菌 16S rRNA 基因的V4–V5可变区建库测序,将构建好的混样文库经文库质检,合格后使用 Illumina Novaseq PE250平台进行测序。
1.5 数据处理与分析
利用FLASH软件对正向和反向测序结果进行拼接,采用Usearch软件去除低质序列和嵌合体,以最低细菌样品质量序列67733为抽平深度,以减少不同测序深度引起的潜在偏差;利用cluster命令对97% 相似性水平的 OTU (operational taxonomic unit)代表序列进行分类学分析,并把代表性OTU与Silva 132核糖体数据库进行细菌序列比对;将OTU表进行随机抽样到相同的测序深度后,于QIIME软件中计算土壤细菌α多样性指数和β多样性距离(weighted unifrac)矩阵。
利用SPSS (version 25)对数据进行统计分析,差异显著性采用单因素方差分析(One-way ANOVA),LSD (P<0.05)进行差异比较;将相对丰度≥0.5%的OTU定义为优势OTU,相对丰度≥0.1%的细菌门和属定义为优势细菌门和优势细菌属,利用Origin(version 2021)绘制土壤细菌物种分布图;利用R Studio (version 4.0.3)进行基于样本 Bray-curtis 距离(OTUs水平)的主坐标分析 (principal co-ordinates analysis,PCoA)和置换多因素方差分析(Adonis),绘制优势OTU热图和相关性热图;利用Canoco软件绘制冗余分析图(redundancy analysis,RDA);利用Spearman相关系数计算OTUs (相对丰度≥0.1%)的相关性,并选取相关系数r≥0.6、P< 0.01的OTUs为数据源[10],于 Gephi (version 9.2)中绘制土壤细菌的共现网络图。
2 结果和分析
2.1 土壤理化性质
由表1可知,与CK相比,绿肥处理(MV和MF)降低了土壤pH和C/N值,但显著增加了土壤全氮含量,其中MF处理的土壤全氮含量最高,达2.58 g/kg;不同施肥处理的土壤pH、全磷、全钾和E-Mg2+的变化趋势均表现为CK>MF>MV。

表1 不同施肥措施下土壤理化性质Table 1 Physical and chemical properties of soil under different fertilization
2.2 土壤细菌群落α多样性
16S rRNA基因测序结果表明,3个样品共得到原始序列863554条,样品含有序列条数67733~106840,以最小序列数67733为抽样深度,抽平后的OTUs数为4430~4495 (表2)。细菌群落Chao1指数、Shannon指数和Simpson指数在各处理间均无显著性差异,说明单施化肥和绿肥施用并未显著改变土壤细菌α多样性。
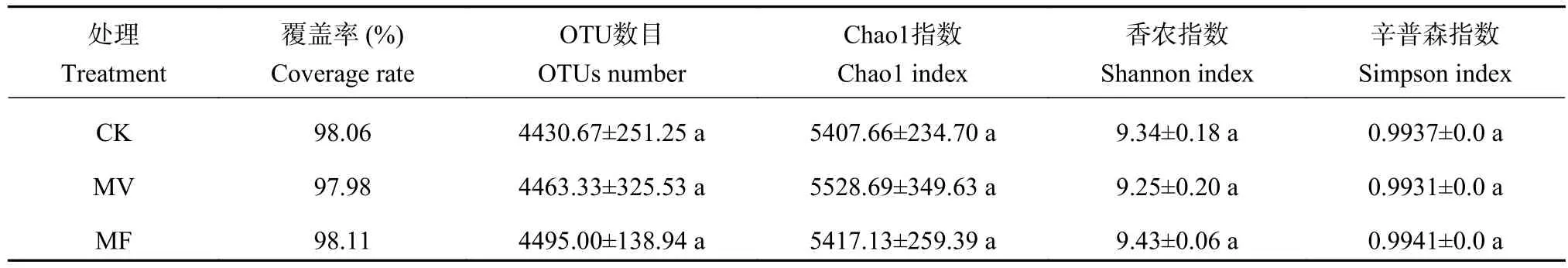
表2 不同施肥措施下土壤细菌α多样性Table 2 α-diversity of soil bacteria under different fertilization
2.3 土壤细菌群落组成与群落结构
在3种施肥处理中,绿弯菌门(Chloroflexi)、变形菌门(Proteobacteria)、硝化螺旋菌门(Nitrospirae)、拟杆菌门(Bacteroidetes)、酸杆菌门(Acidobacteria)、浮霉菌门(Planctomycetes)、疣微菌门(Verrucomicrobia)、放线菌门(Actinobacteria)和匿杆菌门(Latescibacteria)为细菌群落的优势菌门(图1A),CK、MV和MF处理的相对丰度>1%的细菌门分别占总细菌门数的96.63%、95.06%和96.77%。除CK和MF处理的匿杆菌门(Latescibacteria)相对丰度显著高于MV处理外,各处理间的优势菌门无显著性差异。在属水平上,地杆菌属(Geobacter)、厌氧绳菌属(Anaerolinea)、RBG-16-58-14、赭黄嗜盐囊菌属(Haliangium)、小梨形菌属(Pirellula)、厌氧粘细菌(Anaeromyxobacter)为前6位优势菌属(图1B),CK、MV和MF处理相对丰度≥0.5%的细菌属分别占总细菌属的15.26%、15.53%和15.33%。另外,MF处理的部分优势菌属(相对丰度<0.5%)新鞘脂菌属(Novosphingobium)、互养棍状菌属(Syntrophorhabdus)、苯基杆菌属(Phenylobacterium)的相对丰度显著高于CK (图1C),而脱硫酸盐菌属(Desulfatiglans)的相对丰度显著低于CK,说明绿肥配施化肥一定程度上改变了土壤细菌在属水平上的群落组成。从整体来看,PCoA分析结果表明3种施肥处理对细菌群落产生影响(图1D),但Adonis分析(P(CK-MV)=0.7;P(CK-MF)=0.7;P(MV-MF)=0.7)结果表明,各处理间的土壤细菌群落结构无显著性差异。综上可知,绿肥处理并未改变土壤细菌的群落结构,但对不同物种分类水平的细菌丰度有影响。

图1 门水平和属水平土壤细菌优势物种丰度、优势属显著性差异及基于Bray-curtis距离的主坐标(PCoA)群落结构分析(门水平相对丰度<0.1%归为other;属水平相对丰度<0.5%归为other)Fig. 1 Dominant species abundance map at phylum and genus level under different fertilization, significant differences in dominant genera, and principal coordinate analysis based on Bray-curtis distance (relative abundance at the phylum level <0.1% was classified as other; relative abundance at genus level <0.5% was classified as other)
2.4 土壤细菌与土壤理化性质相关性分析
优势OTU热图的分析结果表明,不同施肥处理下的优势OTU有差异(图2A)。冗余分析(RDA)结果表明,环境因子对优势细菌群落结构差异解释量为74.88%,RDA1和RDA2解释量分别为45.44%和29.44%,土壤速效钾、交换性钙离子、pH和全氮是影响细菌群落结构的重要因子,其中速效钾(F=2.6,P=0.04)对微生物群落的影响达到显著水平(图2B)。
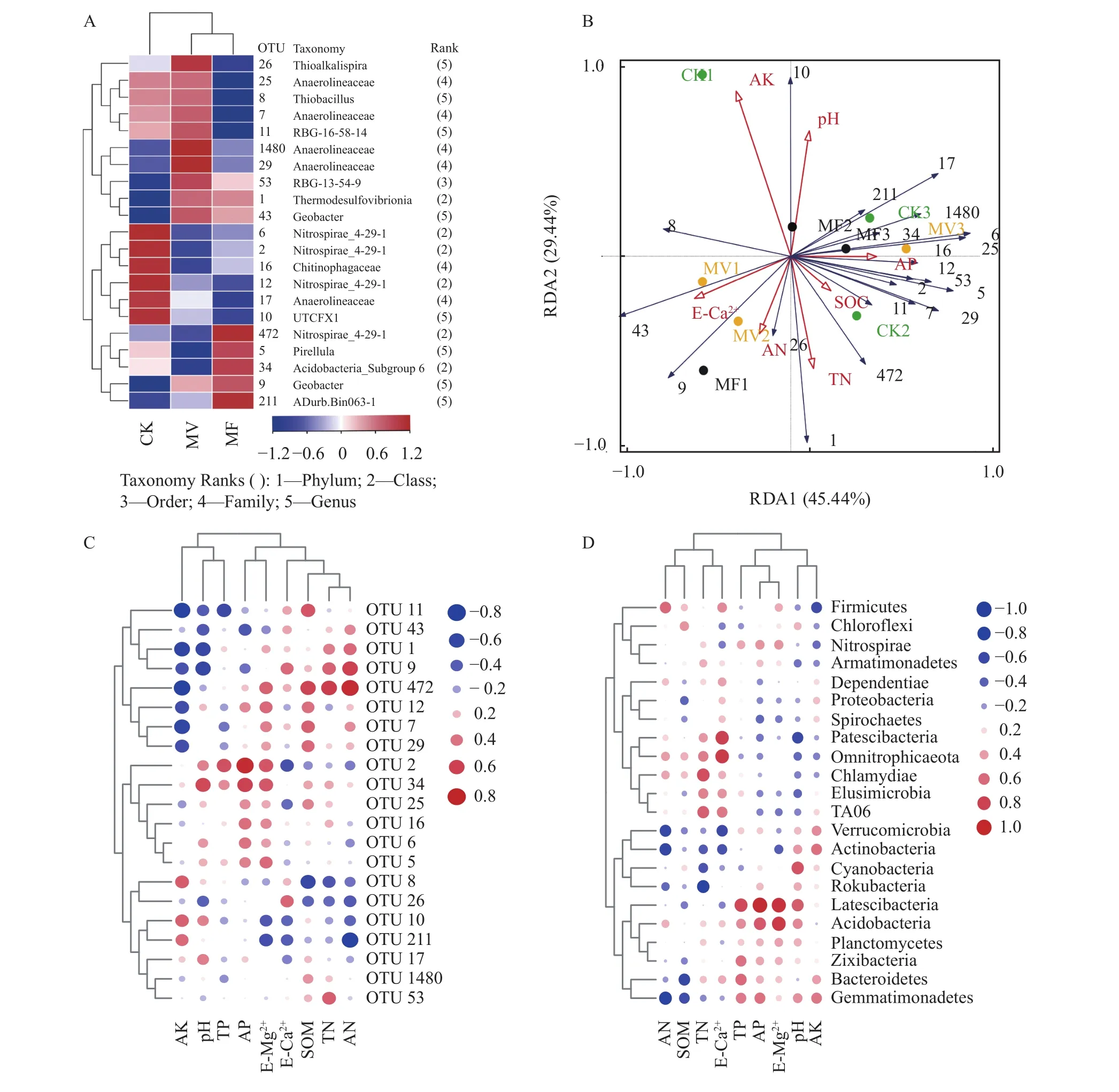
图2 优势OTU热图(A)、优势OTU的冗余分析(B)、优势OTU(C)及优势菌门(D)与土壤理化性质的Spearman相关性分析Fig. 2 Heat map of dominant OTU(A), redundancy analysis (RDA) of dominant OTU(B), Spearman correlation analysis of dominant OTU(C) and phyla (D) with edaphic physicochemical variables
相关分析表明,大多数优势细菌对土壤速效钾、交换性钙离子和全氮含量的响应较为敏感。由图2C可知,土壤速效钾与多数优势OTUs呈负相关,其中速效钾与OTUs (11、472)呈显著负相关;土壤全氮与OTU (472)呈显著正相关。在门分类水平上,多数优势细菌门的相对丰度与交换性钙离子和全氮相关,其中Patescibacteria和Omnitrophicaeota菌门与交换性钙离子呈显著正相关;Chlamydiae菌门与全氮呈显著正相关,而棒状杆菌门(Rokubacteria)与全氮呈显著负相关(图2D)。
2.5 土壤细菌共现网络关系
不同施肥处理的土壤细菌群落表现出不同的共现网络模式(图3)。CK、MV和MF处理的土壤细菌共现网络分别由156、147和160个节点,2409、2099和2112条极显著正相关的边组成,其平均度分别为30.615、28.558和26.400,模块化指数分别为0.752,0.762和0.794。当模块化指数>0.4,表明共现网络具有模块化结构[11]。
由图3可知,CK和MV处理的共现网络相似,MF处理的细菌共现网络相对复杂,表明不同施肥处理对土壤细菌群落共现网络影响较大。由表3可知,各处理的细菌类群在共现网络中所占比例不同,变形菌门(Proteobacteria)和绿弯菌门(Chloroflexi)在共现网络所占比重最大,且MF处理的细菌类群最多。此外,地杆菌属(Geobacter)、厌氧绳菌属(Anaerolinea)、RBG-16-58-14为土壤细菌共现网络中前3位优势菌属,可认为是岩溶区石灰性水稻土的土壤细菌关键类群。
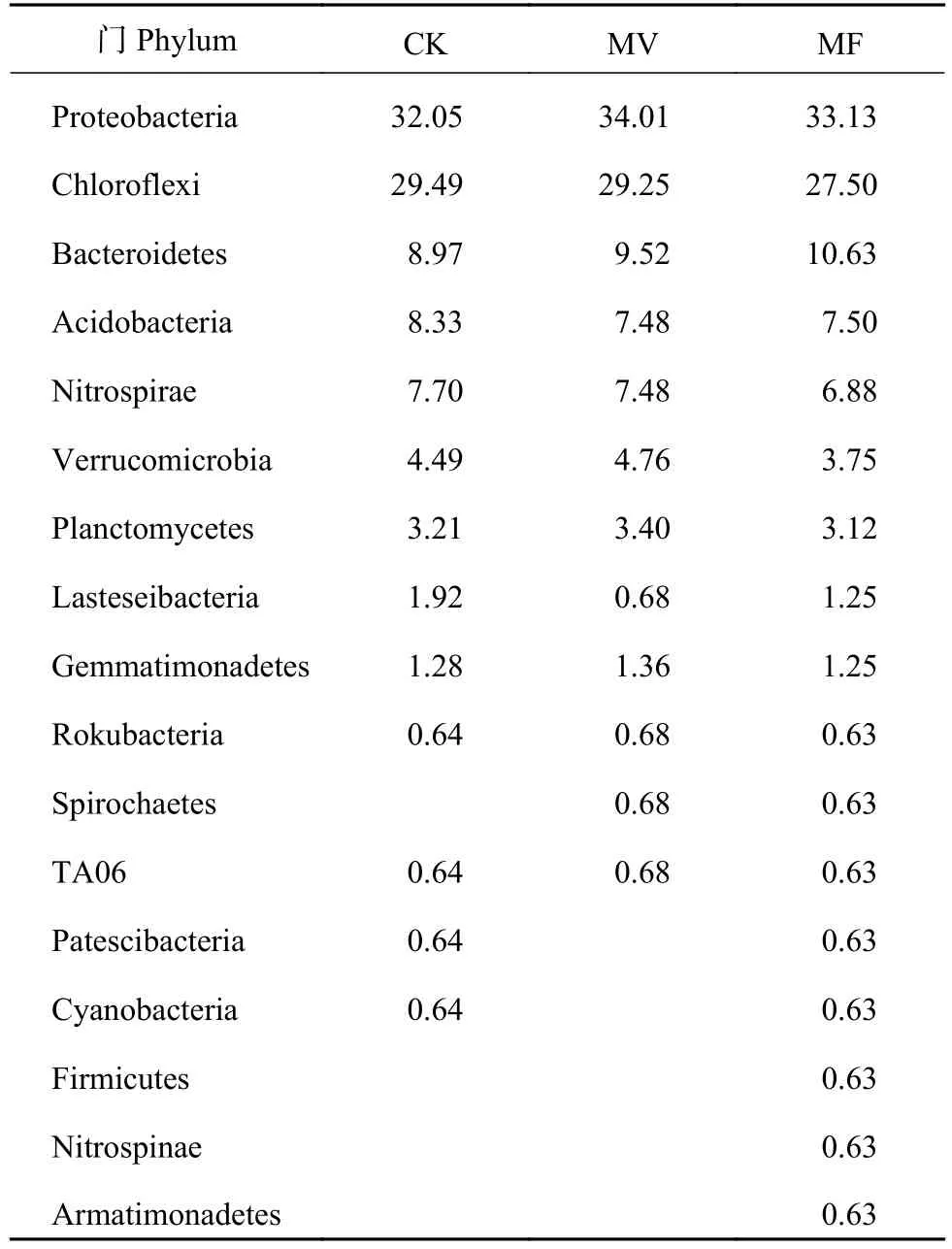
表3 细菌门在共现网络所占比重(%)Table 3 The proportion of bacteria phylum in co-occurrence network
3 讨论
3.1 绿肥施用对土壤理化性质的影响
前人研究表明,长期水稻–绿肥轮作能显著提高土壤有机质、全氮和有效氮的含量,维持土壤养分[12]。本研究中绿肥处理(MV和MF)显著提高了土壤全氮含量(表1),说明豆科绿肥可以改善稻田氮素状况。与CK相比,绿肥处理的土壤有机质和碱解氮含量增幅不明显,可能与试验时间短,土壤有机质积累速度缓慢有关[13–14]。此外,在无碳源输入的情况下,豆科绿肥可活化土壤养分,固定大气氮,翻压后增加土壤有效养分浓度[15],使得C/N值降低。本研究中MF处理的土壤C/N为7.44,CK处理的土壤C/N为8.38,两者存在显著性差异。低碳氮比的豆科绿肥可能迅速释放氮,导致土壤有机氮的矿化[16],且较低C/N的土壤有机质分解速率较快[17],进一步解释了绿肥处理增加土壤有机质和碱解氮含量,但增幅不显著的现象。
在本研究中,土壤pH、速效磷和速效钾含量在不同施肥处理中均表现为CK>MF>MV(表1),但作用机制各不相同。本研究发现,绿肥处理(MV和MF)土壤pH显著低于CK,说明施用绿肥造成土壤pH下降,与严嘉慧等[8]、林叶春等[18]的研究结果一致。可能原因在于尿素水解、硝化过程中产生硝酸根离子[19]或者绿肥腐解过程中产生有机酸[20],导致土壤酸化。磷是植物和生物生长的必需元素,缺磷会导致作物发育迟缓[21],对土壤细菌多样性和群落也起限制作用[22]。石灰性水稻土中碳酸钙含量高[2],施入的磷肥会与钙镁结合,被固定于土壤中成为无效态磷[23],使磷肥的有效性降低。此外,绿肥可提升土壤碳、氮养分,激发微生物生长活性[24],使得微生物对磷的需求增加,导致有效磷优先被土壤微生物所固持[25],这便解释了绿肥处理的土壤有效磷含量显著低于CK原因。而岩溶地区碳酸盐岩钾含量低,绿肥生长需吸收大量钾素[26-27],因此绿肥处理降低土壤速效钾含量(表1),与Wen等[28]研究结果一致。
3.2 绿肥施用对土壤细菌群落多样性的影响
土壤细菌对施肥方式极其敏感,土壤环境的微小变化可导致土壤细菌群落多样性产生显著差异[29–31]。长期施用绿肥显著改变酸性红壤稻田土壤细菌多样性[7],而本研究中绿肥处理并未显著改变石灰性土壤细菌的多样性(表2),与严嘉慧等[8]和周艳飞等[32]研究结果一致。PCoA分析结果表明,不同施肥方式的土壤细菌群落结构有所差异(图1D)。与CK相比,MF处理提高了变形菌门(Proteobacteria)和拟杆菌门(Bacteroidetes)的相对丰度,降低了绿弯菌门(Chloroflexi)、放线菌门(Actinobacteria)和蓝藻菌门(Cyanobacteria)等的相对丰度。此外,MF处理显著增加了新鞘脂菌属(Novosphingobium)、互养棍状菌属(Syntrophorhabdus)、苯基杆菌属(Phenylobacterium)的相对丰度,降低了脱硫酸盐菌属(Desulfatiglans)的相对丰度(图1B和C)。特定土壤条件导致土壤细菌群落组成产生显著差异,如土壤pH[33]、碳氮可利用性[34]和速效磷[35]等环境因子。RDA分析结果表明,土壤速效钾是影响土壤细菌群落组成变化的关键环境因子,且多数优势细菌与土壤速效钾呈显著正相关(图2C和D)。土壤钾主要来源于母质矿化和生物地球化学循环补充,因此微生物可通过酸解、螯合、氧化还原等多种方式的协同作用对含钾岩石生物转化释放钾素[36]。本研究中交换性钙离子含量与土壤细菌呈显著正相关,这是因为钙离子具有影响微生物生长、信号调节、基因表达等生理作用[37]。有研究表明,土壤交换性钙离子是岩溶地区荒漠化过程中影响微生物群落动态变化的关键因子之一,可改变具有碳、氮代谢能力的土壤微生物群落组成,进而影响土壤氮素的富集和有机碳的降解[38]。
3.3 绿肥施用对土壤细菌共现网络的影响
共现网络结果表明,CK和MV处理的土壤细菌共现网络相似,而MF处理土壤细菌群落的网络较为复杂(图3),网络复杂性随土壤养分(如土壤碳、氮的可利用性)增加逐渐复杂[39–40],可见MF处理共现网络复杂性的提高与土壤有机碳、全氮和有效氮含量的提高密切相关。变形菌门(Proteobacteria)、绿弯菌门(Chloroflexi)在3种施肥方式中均占主导地位,但所占比重不同(表3)。MF处理下变形菌门(Proteobacteria)、拟杆菌门(Bacteroidetes)类群在共现网络的比重高于其他处理,绿弯菌门(Chloroflexi)、酸杆菌门(Acidobacteria)、硝化螺旋菌门(Nitrospirae)和疣微菌门(Verrucomicrobia)等细菌类群的比重低于其他处理。变形菌门、拟杆菌门为富营养性细菌,在富营养环境中快速繁殖[41],如拟杆菌门主要分布在上层腐殖质层或根际,在活性有机碳较高的环境中富集[42]。相反,绿弯菌门、酸杆菌门等为寡营养性细菌,具有缓慢生长特性[43–44],在营养贫乏的土壤中普遍存在[45–47]。当微生物所处环境的底物浓度变高时,富营养性会取代寡营养性群落定植在富裕的环境中[48],说明MF处理利于富营养性细菌群落生长,对寡营养性细菌群落生长起限制作用。此外,MF处理的共现细菌类群多于CK和MV处理(表3),说明MF处理有利于土壤细菌群落共享生态位。
关键类群被认为是微生物群落结构和功能的重要驱动力[49]。Zhang等[50]对长期施用绿肥的稻田土壤研究发现,不动杆菌属(Acinetobacter)、假单胞菌属(Pseudomonas)是酸性红壤稻田的关键功能菌属,其相对丰度在31年间翻了5.7~10倍,说明细菌群落经历生态演替,优势细菌占据特定生态位并发挥特定功能。本研究中地杆菌属(Geobacter)、厌氧绳菌属(Anaerolinea)和RBG-16-58-14为土壤细菌共现网络的关键菌属。地杆菌属是专性厌氧固氮菌,常见于淹水条件或深层土壤缺氧环境[51–52],对硝酸盐还原和铁还原有重要作用[53]。厌氧绳菌属是厌氧菌[54],具有分解多种碳化合物的潜力[55],与有机碳的降解和有机化合物的降解密切相关[56],且适应氨氮浓度较高的底物,可能具氨氧化功能[57–58]。而RBG-16-58-14被发现与沉积物种的硝酸盐减少相关[59],在水稻生态系统中还需进一步证明。
4 结论
与单施化肥相比,绿肥处理均能提高土壤全氮含量,降低土壤pH、速效钾含量和C/N值。不同施肥处理均未显著改变细菌群落多样性,绿肥配施化肥处理改变了土壤细菌在门和属水平上的群落组成,土壤速效钾、交换性钙离子和全氮是影响土壤细菌群落组成变化的关键环境因子。单施化肥和单种绿肥处理的土壤细菌共现网络相似,绿肥配施化肥处理增加了土壤细菌网络复杂性及变形菌门、拟杆菌门等富营养性细菌的相对丰度。因此,综合3种施肥方式可知,绿肥配施化肥能提高土壤养分含量和改变土壤细菌群落组成,在岩溶区稻田生态系统发挥着重要作用。
