鬼手海葵(Aiptasia pulchella)白化恢复后的基因表达差异
2021-12-04张倩闫昊王路
张倩, 闫昊, 王路
闽江学院海洋研究院, 福建 福州 350108
刺胞动物(Cnidaria)多数生活在海洋之中, 少数见于淡水(李新正 等, 2020), 刺胞动物门分水螅纲(Hydrozoa) 、 钵水母纲(Scyphozoa) 和珊瑚纲(Anthozoa)。珊瑚纲和虫黄藻(zooxanthellae)的胞内共生关系在海洋环境中普遍存在, 共生藻的存在是其进行正常生理活动所必不可少的。虫黄藻是一种黄褐色单细胞藻类, 属于共生甲藻属(Symbiodinium), 其与宿主的互利共生机制在生态系统中发挥着重要的作用。共生藻吸收宿主代谢产生的二氧化碳、磷酸盐和硝酸盐等, 并通过光合作用将能量转化为共生体所需要的营养, 而宿主生存的能量主要来自共生藻的光合作用产物, 同时宿主为虫黄藻提供庇护场所, 使其避免被植食性动物捕食(Freudenthal, 1962; Koch et al, 2020)。然而, 共生功能体对环境变化十分敏感, 虽然宿主可通过调节新陈代谢通路来应对环境改变, 减轻或消除环境胁迫所造成的影响, 但当环境条件恶化至共生功能体耐受极限时, 宿主就会排出体内的共生藻, 或者共生藻色素(叶绿素a、叶绿素b、类胡萝卜素、甲藻素)含量减少, 从而使得宿主外表逐渐变浅甚至变白,即“白化”(Douglas, 2003; Zhu et al, 2004; Van Treuren et al, 2019)。造成白化的原因相当复杂, 如海水营养或温度变化(Sawyer et al, 2001; Weng et al,2014)、化学污染(Morgan et al, 2012; De Orte et al,2019)、紫外线辐射增强(Baruch et al, 2005)、海水盐度不适以及病原体侵染等。白化会降低宿主的繁殖能力、生长速度, 改变共生藻的群落结构, 严重时会导致宿主死亡, 危害海洋生态环境(Van Treuren et al,2019)。因此, 研究共生功能体在抵御环境过程中如何进行自我调节、适应及修复, 已成为目前该领域亟待解决的科学问题。
在胁迫强度较轻且持续时间较短的情况下, 珊瑚或海葵可以通过生理上的保护机制以适应环境的变化, 且在环境条件好转后恢复健康。珊瑚礁白化后的恢复与白化程度有关, 大范围白化的珊瑚礁完全恢复需要几年到几十年。生理适应机制主要通过叶黄素循环, 珊瑚色素荧光, 活性氧清除系统, 分泌紫外线吸收物质和产生热休克蛋白等方式来实现(李秀保 等, 2007)。倘若因白化异常严重或环境威胁强度较大而导致持续白化, 宿主将会死亡。近年研究发现虫黄藻密度降低导致宿主白化的细胞机制主要有: (1)虫黄藻自身的死亡和降解; (2)虫黄藻的胞外分泌引发其从宿主体内被释放出来; (3)细胞凋亡和细胞坏死, 在白化过程中, 宿主可通过细胞凋亡和细胞坏死两种方式降低虫黄藻密度; (4)宿主细胞分离, 在环境胁迫下, 含有虫黄藻的完整内胚层细胞, 脱离宿主组织并很快分解, 从而释放出虫黄藻(Dunn et al, 2002; Bieri et al, 2016)。也有研究表明,在不同的环境胁迫下, 宿主失去虫黄藻的方式也会有所不同, 如Tamaki Bieri 以海葵(Aiptasia)为实验对象, 证明在热胁迫和光胁迫下, 宿主细胞将共生藻排出细胞是导致白化的主要方式; 在低温胁迫下,藻类的降解和宿主细胞自身的脱离是导致白化的主要方式(Bieri et al, 2016)。
近年来, 由于受到全球气候变化的影响, 海洋表面平均温度升高, 珊瑚纲白化死亡现象日益严重,进而威胁海洋生态系统的平衡。随着分子生物学的发展, 探究白化机制已经成为了当今研究热点, 高通量测序为开展性状的分子基础研究提供了重要的技术支持, 是研究基因功能的基础, 对生物体的发育和疾病研究具有重要作用。通过分析宿主在环境胁迫条件下的基因表达与调控, 可以深入了解相关基因在宿主白化与恢复机制中的作用。目前, 越来越多的研究报道了宿主白化后的生理变化, 以及白化死亡过程中功能基因的表达量变化, 为探究白化机制奠定了基础。鬼手海葵(Aiptasia pulchella)属于珊瑚纲, 海葵目(Actiniaria), 因具有在实验室更易生长和繁殖的优点, 被广泛用于珊瑚纲生理学、共生功能体、白化机制和重金属富集等研究(Howe et al, 2012, 2014)。本文以鬼手海葵为研究对象, 通过转录组测序技术, 比较长期热胁迫后白化海葵和白化恢复后海葵之间的基因表达量差异, 探究海葵共生体系在共生失衡后进行适应性调节的分子机制,从而增进对内共生模式稳定与调节机制的理解。
1 材料与方法
1.1 样品收集和总RNA 提取
鬼手海葵培养于实验室的循环海水水族箱中,海水温度为(25±1)℃, 盐度32‰~34‰, pH=8.1, 光暗周期为12∶12, 光照强度162.5μmol·m-2·s-1。对鬼手海葵进行慢性热胁迫实验, 即改变环境温度(29±1)℃, 光照周期和光照强度不变, 两周后, 海葵整体发生白化。将海水温度恢复(25±1)℃, 在正常培养条件下继续培养两周, 约70%的海葵仍处于白化状态(图1a、b), 约30%的海葵逐渐恢复正常状态(图1c、d), 与正常培养的海葵相比无明显差异(图1e、f), 取白化海葵(AP_W)和白化恢复后的海葵(AP_H),每 6 个个体混合成一个转录组测序样品, 通过TRIzol 法提取RNA, 琼脂糖凝胶电泳和Nanodrop分析样品RNA 浓度和完整性, 用于转录组测序。

图1 海葵形态照片a、b 为白化后未恢复的海葵; c、d 为白化后恢复的海葵; e、f 为正常培养的海葵Fig. 1 Morphological changes of A. pulchella. (a) and (b) indicate bleaching A. pulchella. (c) and (d) indicate bleaching recovering A. pulchella. (e) and (f) are the control
1.2 cDNA 文库建库和转录组测序
鬼手海葵的转录组建库测序和拼接组装委托北京诺禾致源生物信息科技有限公司完成。建库使用的试剂盒为Illumina 的NEBNext® UltraTM RNA Library Prep Kit, 以mRNA 为模板, 构建cDNA 双链, 库检合格后, 通过Illumina Hi-seq 进行测序。为了保证数据分析的质量及可靠性, 对原始数据进行过滤, 去除带接头、含N、低质量reads, 同时对Clean data 进行Q20、Q30 和GC 含量计算, 后续所有分析均是基于Clean data。
1.3 基因注释和基因富集分析
从基因组网站(http://aiptasia.reefgenomics.org/download/)下载参考基因组和基因模型注释文件(Baumgarten et al, 2015)。使用HISAT2 v2.0.5 将配对末端 Clean reads 与参考基因组比对, 采用StringTie 进行新基因预测。基因表达水平的评估采用的是FPKM(Fragments Per Kilobase of transcript per Million mapped reads)值(Trapnell et al, 2010)。组间基因的差异表达分析使用edgeR R 软件包(3.18.1)进行, 使用 Benjamini&Hochberg 方法调整P值(padj), 将校正后的P值以及|log2(fold change)|作为显著差异表达的阈值来筛选差异表达基因(differentially expressed genes, DEGs), 筛选条件为|log2(fold change)|>1 并 且 padj<0.05, 通 过clusterProfiler R 软件实现差异表达基因的GO 富集分析和KEGG 通路中差异表达基因的统计富集。
利用GSEA v4.10 软件(https://www.gsea-msigdb.org/gsea/downloads.jsp)对28050 个基因进行KEGG和GO 基因集富集分析(Subramanian et al, 2005), 数据库(Molecular Signatures Database, 简称MSigDB,网址: https://www.gsea-msigdb.org/gsea/msigdb/collections.jsp) 的“c2.cp.kegg.v7.1.symbols.gmt” 和“c5.all.v7.1.symbols.gmt”作为参照数据集, Number of permutations 设置重复为 1000 次, Nominal p-value<0.05 的富集结果被认为差异具有统计学意义(Subramanian et al, 2005)。
2 实验结果
2.1 鬼手海葵转录组数据质量和基因统计
经过原始数据过滤、测序错误率检查、GC 含量分布检查等, 获得后续分析使用的Clean reads, 数据汇总如表1 所示。白化的鬼手海葵样品(AP_W)经过HiSeq2500 高通量测序获得51938216 条原始reads 数, 数据过滤后得到50109686 条reads 数, Q20值和Q30 值为96.52%和90.82%, GC 含量为41.42%,基因数目是24565 个, 新基因有1886 个。白化恢复后的鬼手海葵样品(AP_H)测序后获得43920530 条原始reads 数, 原始数据过滤后得到43163786 条reads数, Q20 值和Q30 值为97.86%和93.97%, GC 含量为43.42%, 基因数目是24157 个, 新基因有1869 个。

表1 鬼手海葵的转录组测序基本信息Tab. 1 Information of transcriptome sequencing of A. pulchella
2.2 差异表达基因及基因注释
将白化海葵和恢复的海葵转录组进行比较,共存在214 个差异表达基因, 其中白化海葵高表达的基因有101 个, 恢复的海葵高表达基因有113 个(图2)。
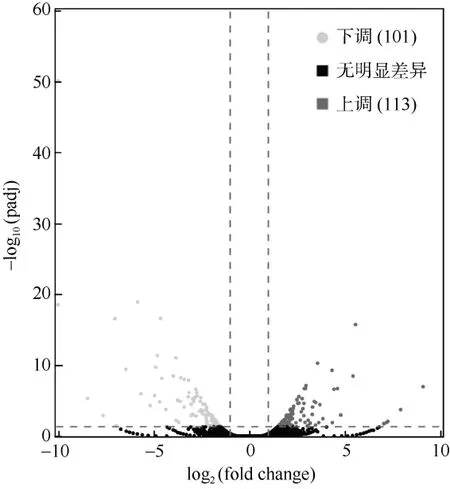
图2 白化海葵和恢复的海葵差异表达基因火山图浅灰色表示下调, 即白化恢复的海葵显著低表达基因有101 个;深灰色表示上调, 即白化恢复的海葵显著高表达的基因有 113个 ; 黑色表示无显著差异; -log10(padj)为基因差异表达显著性;|log2(fold change)|表示基因表达量差异倍数Fig. 2 Volcano plot of differentially expressed genes between bleaching and recovering A. pulchella. Light gray indicates 101 down-regulated in recovering A. pulchella,and dark grey indicates 113 up-regulated in recovering A.pulchella. Black indicates no significant difference.-log10(padj) is significant difference in gene expression.|log2(fold change)| indicates the fold change of gene expression.
根据214 个差异表达基因的注释信息, 主要分为4 类基因(表2): (1)与DNA 复制有关的基因: 解旋酶、组蛋白和核酸内切酶, 这些差异基因均是在AP_H 组中高表达, 说明白化恢复后中的海葵在DNA 复制方面更活跃; (2)与代谢有关的基因, 如磷脂酶、葡萄糖醛酸酶、碳酸酐酶等酶类, 说明白化影响宿主的新陈代谢; (3)与离子转运有关的基因,如阳离子转运蛋白或转运体, 在白化和白化恢复后存在显著差异; (4)与胶原蛋白相关的基因: 包括腱生蛋白、基质胶、胶原蛋白、纤维蛋白等11 个基因,即该类基因与海葵白化关系密切。
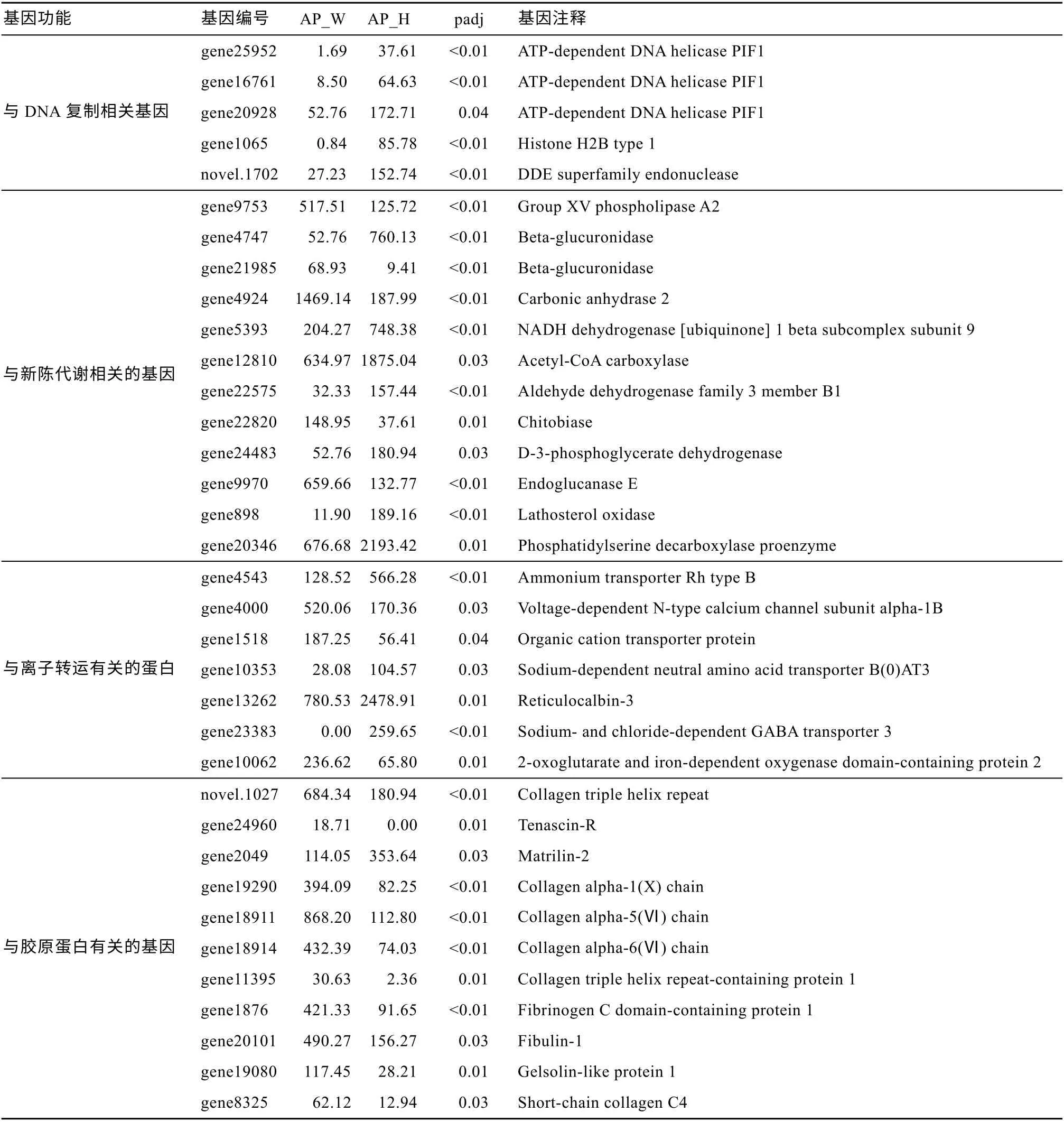
表2 与DNA 复制、代谢、离子转运和胶原蛋白相关的差异表达基因Tab. 2 Differentially expressed genes associated with DNA replication, metabolism, ion transport, and collagen
2.3 差异基因和基因集富集分析
对214 个差异基因进行GO 富集分析结果发现,共有23 个GO 条目显著富集(padj<0.05)(图3), 富集最高的 2 个 GO 条目为“脂质代谢过程”(lipid metabolic process)和“神经递质传输”(neurotransmitter transport), 均属于生物过程。由于差异表达基因数目过少, 没有富集到有显著差异的KEGG 通路。
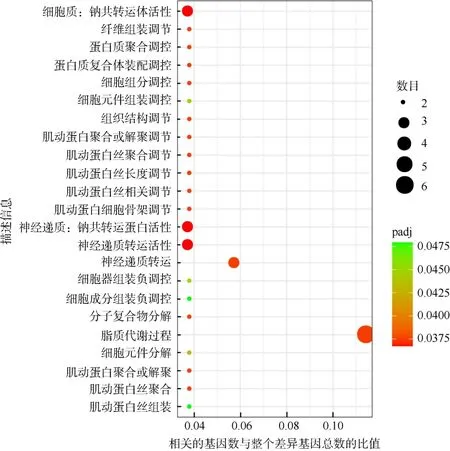
图3 白化海葵和白化恢复后的海葵之间差异表达基因富集的GO 功能分类Fig. 3 GO term distributions of differentially expressed gene enrichment between bleaching and recovering A. pulchella
对28050 个基因进行基因集富集分析, 在以c2.cp.kegg.v7.1.symbols.gmt 为参照基因集的KEGG基因集富集分析结果显示, 白化恢复后的海葵在DNA 复制(DNA_REPLICATION)、同源重组(HOMOLOGOUS_RECOMBINATION)、核苷酸修复(NUCLEOTIDE_EXCISION_REPAIR)和蛋白酶体(PROTEASOME)等信号通路显著富集, 白化海葵仅在配体和受体的相互作用(N E U R O A C T I V E_LIGAND_RECEPTOR_INTERACTION)通路显著富集(Nominal p-value<0.05)(图4)。以c5.all.v7.1.symbols.gmt 为参照基因集的GO 基因集富集分析中,白化恢复后的海葵在核染色体分离(NUCLEAR_CHROMOSOME_SEGREGATION)、染色体分离(CHROMOSOME_SEGREGATION)、催化活性对 DNA 的作用(CATALYTIC_ACTIVITY_ACTING_ON_DNA)、DNA 构象变化(DNA_C O N F O R M AT I O N_C H A N G E)和端粒组织(TELOMERE_ORGANIZATION)等方面显著富集,而白化海葵在神经递质受体的活动(NEUROTR ANSMITTER_RECEPTOR_ACTIVITY)、突触后神经递质受体活动(POSTSYNAPTIC_NEURO TRANSMITTER_RECEPTOR_ACTIVITY)、胶原蛋白三聚物(COLLAGEN_TRIMER)、细胞外配体门控离子通道活性(EXTRACELLULAR_LIGAND_GATED_ION_CHANNEL_ACTIVITY)和配位门控离子通道活性(LIGAND_GATED_ION_CHANNEL_ACTIVITY)方面显著富集(Nominal p-value<0.05)(图5)。

图4 GSEA 富集中差异显著的KEGG 基因集(Nominal p-value<0.05)FDR: 错误发现率; Enrichment score (ES): 富集评分; Normalized ES: 归一化的富集评分; Signal2Noise: 信噪比; Ranked in ordered gene set: 基因在特定基因集中的分布Fig. 4 Significantly different KEGG pathway of Gene Set Enrichment Analysis (Nominal p-value<0.05). FDR indicates false discovery rate. ES indicates enrichment score. Normalized ES indicates normalized enrichment score. Signal2Noise indicates signal to noise ratio; Ranked in ordered gene set, the distribution of the gene ranked in ordered gene set.

图5 GSEA 富集中差异显著的GO 基因集(Nominal p-value<0.05)FDR: 错误发现率; ES: 富集评分; Normalized ES: 归一化的富集评分; Signal2Noise:信噪比; Ranked in ordered gene set: 基因在特定基因集中的分布Fig. 5 Significantly different GO pathway of Gene Set Enrichment Analysis (Nominal p-value<0.05). FDR indicates false discovery rate. ES indicates enrichment score. Normalized ES indicates normalized enrichment score. Signal2Noise indicates signal to noise ratio; Ranked in ordered gene set, the distribution of the gene ranked in ordered gene set.
3 讨论
白化是一个复杂的过程, 越来越多的研究人员开始从功能基因差异表达等角度来解释白化机制(Hou et al, 2018; Hu et al, 2020)。在珊瑚白化死亡过程中, 会引起细胞的骨架重排、新陈代谢和信号传导等细胞过程改变, 相关的基因表达量也会发生变化(Desalvo et al, 2008)。有研究提出了白化机制假想模型, 该模型认为温度胁迫导致宿主细胞、虫黄藻中的活性氧簇(reactive oxygen species, ROS)释放,ROS 在细胞内积累从而扰乱了细胞内Ca2+平衡, 细胞内Ca2+浓度失衡导致细胞骨架重排、粘附性改变、钙化率下降并最终导致细胞死亡(Sawyer et al,2001)。也有研究人员认为活性氧的产生和积累导致氧化应激, 如超氧自由基、单态氧、过氧化氢和羟自由基等还原氧中间体的产生和积累会导致脂质、蛋白质和DNA 的损伤, 这也是导致珊瑚白化的原因之一(Nii et al, 1997; Lesser, 2011)。为了探究海葵—虫黄藻共生体对高温的响应机理, 本文对鬼手海葵进行了慢性热胁迫实验, 通过分析热胁迫后白化与白化恢复个体的基因表达量变化, 来探究海葵对外界环境进行响应与适应的分子调节机制。
在本研究中, 与白化的海葵相比, DNA 复制和核酸修复相关基因(解旋酶、组蛋白和核酸内切酶)在白化恢复后的海葵中表达量更高, 而且KEGG 和GO 基因集富集分析的结果也显示在白化恢复后的样品中, DNA 复制、同源重组、核苷酸修复、染色体分离、DNA 构象变化、端粒组织显著富集。已有研究表明, 环境异常会引起共生体DNA 损伤, 机体会分泌抗炎性物质修复受损细胞, 以去除受损大分子相关基因、维持DNA 稳定与修复(Douglas, 2003)。另有研究显示, 诱导珊瑚白化的恶劣环境会对共生体的DNA 造成损伤(Baruch et al, 2005)。丛生盔形珊瑚(Galaxea fascicularis)因热胁迫发生白化后, 与DNA 重组、DNA 代谢过程和DNA 融合相关的基因显著上调(Hou et al, 2018)。与未出现白化的个体相比, 白化恢复后的加勒比海珊瑚(Orbicella faveolata)与DNA 结合和RNA 加工过程相关的基因表达水平更高(Pinzón et al, 2015)。
除了与DNA 复制和核酸修复相关的差异基因,我们还筛选到11 个与胶原蛋白相关的基因的表达量水平在两个转录本中存在显著差异。此结果与Sawyer 等(2001)报道的鬼手海葵在受到热胁迫后,细胞粘附功能发生紊乱的现象所一致。在加勒比海珊瑚白化过程中, 与细胞粘附和识别相关的通路和基因也发生了显著变化(Pinzón et al, 2015)。此外,还有一些差异基因和离子转运以及代谢有关, 而且基因集富集分析结果也显示两个比较组在与离子通道相关的基因集方面存在显著差异。在鬼手海葵中,铁转运蛋白ApTF的表达量会因热胁迫而受到显著抑制(Song et al, 2015), 与阳离子结合相关的基因也会发生改变, 严重时会诱发细胞凋亡和细胞死亡(Bieri et al, 2016)。
目前已有的研究多关注于胁迫后的白化过程,而对白化恢复后的适应性机制研究较少。白化适应性体现在个体本身的恢复力(resilience), 海葵白化后若生活环境有所改善, 又可恢复到原来的共生稳态(黄晖 等, 2013)。珊瑚或海葵在白化后的恢复现象表明, 宿主确实具有自我修复与适应的能力(Buddemeier et al, 1993)。无论是在受到环境胁迫还是处在恢复阶段, 宿主基因表达水平变化明显, 检测其基因表达量情况, 同时结合其他生理生态数据,能在一定程度上有效评价生态系统的健康情况并预测白化或者恢复, 为资源保护提供帮助。基于此, 本研究从转录组水平初步探索了参与鬼手海葵白化与恢复的分子机制, 可为后续相关研究提供参考。
