我国海南省东南部台湾那屿藻的形态观察及系统发育分析
2021-12-04刘金梅黄冰心丁兰平王雪聪闫璟闫盼竹张瑶
刘金梅, 黄冰心, 丁兰平, 王雪聪, 闫璟, 闫盼竹, 张瑶
天津师范大学生命科学学院, 天津动植物抗性重点实验室, 天津 300387
那屿藻属(新拟名)YonaguniaKawaguchi et Masuda 隶属于红藻门 Rhodophyta 海膜科Halymeniaceae, 于 2004 年建立, 因其模式种YonaguniatenuifoliaKawaguchi et Masuda 采集于琉球群岛的与那国岛(Yonaguni Island)而得名。该属最初的报道包括模式物种Y. tenuifoliaKawaguchi et Masuda 和台湾那屿藻( 新拟名)Y.formosana(Okamura) Kawaguchi et Masuda 两个物种。那屿藻属的藻体一般扁平, 二叉式分枝或不规则分枝, 藻体表面或边缘偶有或具有许多小育枝, 藻体内部结构为多轴型, 具有紧密的皮层, 髓部由密集交错的髓丝组成, 髓丝为纵向排列; 果孢枝枝丛和辅助细胞枝枝丛具有几个简单的次生生殖枝丛; 辅助细胞与连络丝融合后, 辅助细胞枝丛的原始生殖枝丛和次生生殖枝丛的大部分细胞同时分裂, 每个细胞形成一簇小的球形细胞, 这些小球形细胞随后形成总生殖枝丛; 果孢子由松散的总生殖枝丛包围, 嵌在髓部; 四分孢子囊由皮层细胞产生, 嵌于皮层内,十字形分裂(Kawaguchi et al, 2004)。
至今, 那屿藻属已报道了8 个种:Y.atimo-vataeManghisi, M. Morabito, G.H.Boo, S. M. Boo et Le Gall;Y.formosana(Okamura) Kawaguchi et Masuda;Y.ligulata(Harvey ex Kützing) Manghisi, M.Morabito, De Clerck et Le Gall;Y.tenuifoliaKawaguchi et Masuda;Y.zollingeri(Sonder) G. H.Boo, J. K. Park, G. S.Gerung et S. M. Boo;Y.maillardii(Montagne et Millardet) Showe M. Lin, Y.C. Chuang et De Clerck;Y.palmataS. M. Lin, Y. C.Chuang et De Clerck;Y.taiwani-borealisShowe M.Lin, Y. C. Chuang et De Clerck(Guiry, 2020; Lin et al,2020)。其中,Y.formosana、Y. ligulata、Y.maillardii和Y.zollingeri是分别由锯齿藻属PrionitisJ.Agardh、海柏属PolyopesJ.Agardh 和盾果藻属CarpopeltisF.Schmitz 等属转入该属的物种。该属物种主要分布于热带至亚热带地区, 但它们的地理分布范围有所不同。有些物种分布较广泛, 如Y.maillardii广泛分布于印度洋—太平洋地区(Lin et al, 2020), 台湾那屿藻Y.formosana在我国台湾地区(Kawaguchi et al, 2004)和海南省(丁兰平 等, 2015)、日本(Titlyanov et al, 2016)、越南(van Nguyen et al,2013; Titlyanov et al, 2015)、马来西亚(Kawaguchi et al, 2004)、菲律宾(Ang et al, 2014)以及太平洋中部的波利尼西亚(Tsuda et al, 2013)等地均有报道; 而有些物种的分布范围比较窄, 如Y. atimo-vatae、Y.tenuifolia和Y.zollingeri分别仅在马达加斯加、日本(Manghisi et al, 2015)和印度尼西亚(Boo et al, 2013)有报道,Y.ligulata在马达加斯加和斯里兰卡有报道(Manghisi et al, 2015),Y. palmata和Y.taiwani-borealis仅在我国台湾地区(Lin et al, 2020)等地有报道。根据分子进化分析, 该属起源于始新世—渐新世, 物种多样性则主要集中在中新世(Lin et al, 2020)。
我国对那屿藻属的研究较少, 之前仅台湾那屿藻Y.formosana在我国台湾地区和海南省有报道,今年又增加了3 个种, 即Y.maillardii、Y. palmata和Y.taiwani-borealis(Lin et al, 2020), 但目前国际数据库中仅记录了来自越南和马来西亚的台湾那屿藻的分子数据(Kawaguchi et al, 2004), 而我国尚无对应的DNA 序列可供参考。因此, 我国那屿藻属在分子方面的研究对于补充该属物种的基因多样性具有一定意义。此外, 那屿藻属是一个较新的属, 对其了解较少, 该属大部分物种的地理分布范围还有待研究。基于此, 本文主要对采集于海南省三亚市大东海和琼海市博鳌地区的台湾那屿藻进行形态学研究, 生成了相应的rbcL 序列, 并结合已知的那屿藻属物种进行了系统发育分析, 为认知和丰富那屿藻属的多样性提供科学依据。
1 材料与方法
1.1 材料
样品分别于2016 年12 月和2018 年4 月采集于海南省三亚市大东海和琼海市博鳌地区。样品处理分为三份, 即液浸标本(用浓度为4%的海水-福尔马林溶液浸泡, 主要用于形态学研究)、腊叶标本(用作凭证标本和形态学研究)和干燥样品(用于分子生物学的研究), 这三者的保存编号相同, 标本存放于天津师范大学生命科学学院藻类实验室。
1.2 实验方法
1.2.1 形态学研究
观察藻体的大体外观、色泽、质地、藻体主轴、分枝情况等(体式显微镜Nikon SMZ25); 再利用冷冻切片技术(生物冷冻切片机 LEICA CM 1860 UV)对样品的特征部位进行切片, 显微镜下观察藻体皮层细胞大小和层数、髓部细胞大小和排列情况、四分孢子囊着生位置和大小以及囊果的着生位置等(正置荧光显微镜LEICA DM5000 B); 最后对以上的各部分特征进行观察、数据测量及拍照。
1.3 系统发育学研究
1.3.1 DNA 样品处理
选取干燥样品进行DNA 提取, 通过选取引物rbcL(F-993 和R-rbcS start)(Freshwater et al, 1994)对所提取的DNA 进行PCR 扩增, 反应体系为25μL,包括12.5μL mix, 10μmol·L-1正向引物1μL, 反向引物1μL, ddH2O 9.5μL, 模板1μL。扩增程序: 95℃预变性4min, 94℃变性1min, 45℃退火2min, 72℃延伸3min, 35 个循环, 72℃终延伸5min。PCR 产物检测采取1%的琼脂糖凝胶电泳, 后将产物送至金唯智公司进行纯化及测序。
1.3.2 数据处理
选取蜈蚣藻Grateloupia filicina和Carpopeltis phyllophora作为外群, 在Genbank 中下载相关那屿藻属物种以及外群物种的 DNA 序列, 使用MEGA7.0 软件进行多序列比对(Clustal W), 遗传距离的分析, 以及系统发育树的构建, Bootstrap 重复1000 次, 其中最大似然树的最适模型为T92+G, 最大简约树的方法选择TBR。
2 研究结果
2.1 形态学
台湾那屿藻Yonaguniaformosana(Okamura)Kawaguchi et Masuda
基名:CarpopeltisformosanaOkamura
同型异名:Prionitis formosan(Okamura)Kawaguchi et Nguyen
生境: 着生于潮间带中部岩石上。
特征描述: 藻体直立, 丛生, 暗紫红色, 偶有蓝灰色, 硬软骨质。藻体扁压, 高2.8~8.5cm, 基部具有盘状固着器; 二叉式分枝或不规则分枝(图1a、b), 枝端形状多样(图1c—g), 卵形或长圆形; 藻体表面和边缘具有许多小育枝(图1h、i), 约0.5~3mm。藻体干燥后变硬, 制成的腊叶标本不能附着于纸上。藻体内部构造由皮层和髓部组成(图1j); 藻体横切面皮层厚约16~83.5μm, 由3~9 层小的椭圆形细胞组成(图1k、l); 髓部厚约260~475μm, 由丰富的纵向错综的髓丝组成(图1n), 髓丝直径5~14μm(图1m)。
四分孢子囊着生于皮层内(图1p), 长卵形, 宽约7~9.4μm, 长约19~25.6μm, 十字形分裂或对半分裂(图1q、r)。囊果着生于最末小枝或小育枝枝端, 微突。囊果切面观, 位于髓部间, 呈球形或近球形; 果孢子囊为近圆形或不规则卵形, 周围具有不育丝组成的囊果被(图1s、t)。未见精子囊结构。
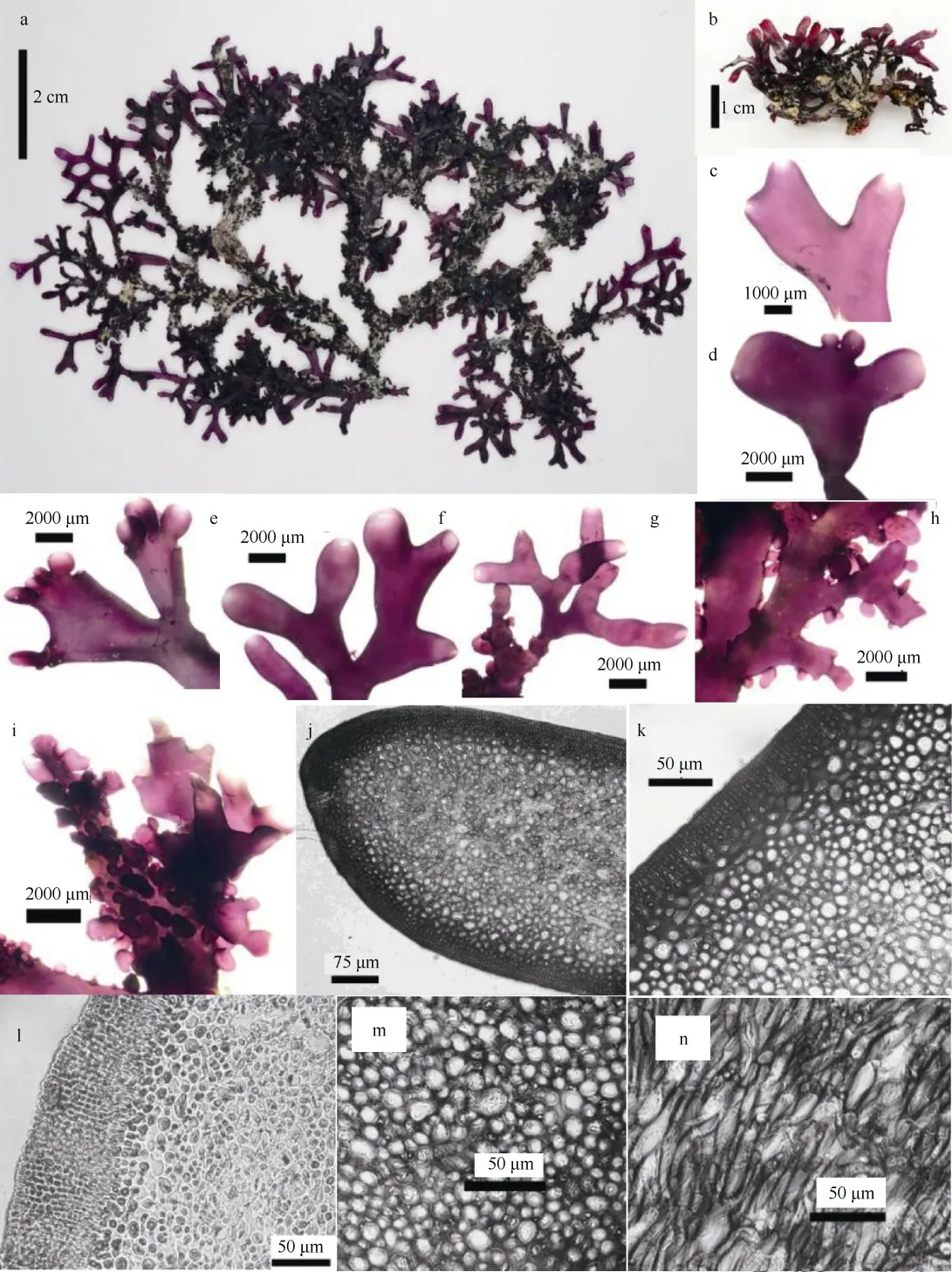
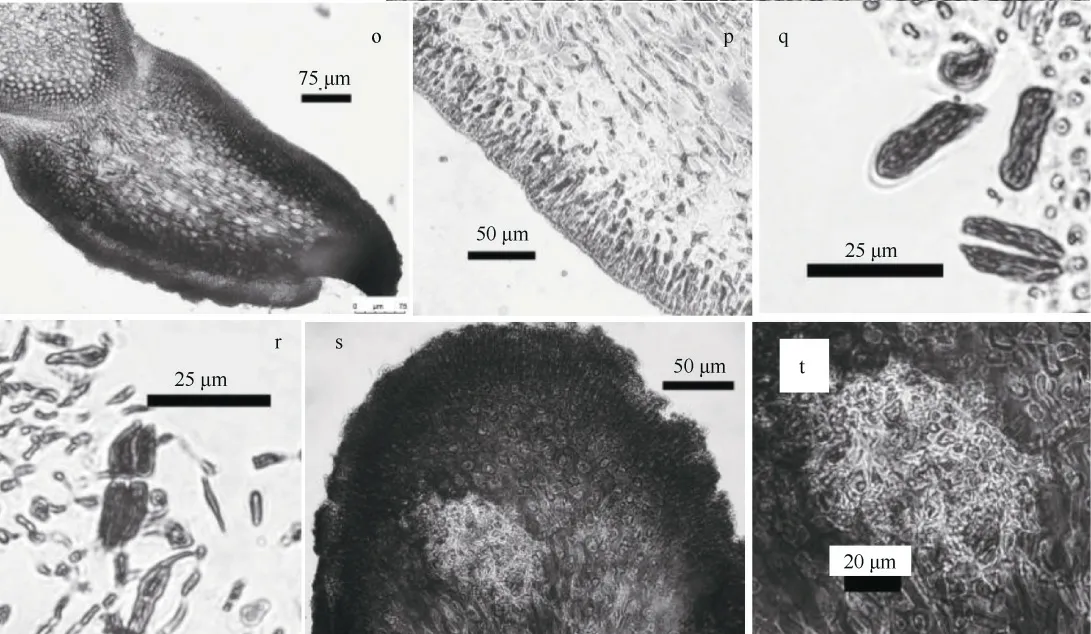
图1 台湾那屿藻的形态和结构a—b. 藻体外观; c—g.枝端形状; h—i. 藻体小育枝(边缘、表面); j. 藻体部分横切面; k—l. 皮层细胞; m. 髓部横切面; n. 髓部纵切面;o. 小育枝纵切面; p—r. 四分孢子囊; s—t. 果孢子Fig. 1 Yonagunia formosana: a-b. habit sketch of the frond; c-g. upper portion of the branchlets; h-i. proliferations (edge,surface); j. transection of part of a frond; k-l. cortical cells; m. transection of medulla; n. longitudinal section of medulla; o.longitudinal section of proliferations; p-r. tetrasporangia; s-t. carpospores
模式产地: 中国台湾地区兰岛(Orchid Island);
标本采集地: 中国海南省三亚市大东海和琼海市博鳌
国外分布: 越南、马来西亚、菲律宾、日本;
国内分布: 海南省、台湾地区
研究编号: 161203005A(三亚)、201804180 09(博鳌)、180418009N(博鳌)
2.2 基于rbcL 序列的系统发育分析
用于构建rbcL 最大似然树的序列共28 条, 其中25 条来自GenBank(23 条为那屿藻属序列, 2 条为外群), 3 条为新生成的序列, 序列的最终比对长度为335bp, 其中, 那屿藻属序列间相似度为93.41%,保守位点为300bp, 可变位点为35bp, 简约信息位点为34bp。基于ML 法(Maximum Likelihood)、MP法(Maximum parsimony)和NJ 法(Neighbor-joining)分别构建了该属rbcL 序列的系统发育树, 鉴于这三种方法所构建的系统发育树的拓扑结构大致一样,因此本文选择了ML 法构建的系统发育树显示(图2),其中节点上的数值表示为自展支持值, 从左到右或从上到下依次为ML、MP、NJ。结果显示, 目前可参考的那屿藻属物种构成了 6 个进化支(Y.maillardii、Y.palmata和Y.taiwani-borealis尚无可参考的序列), 其中本研究序列161203005A(三亚)、180418009N(博鳌)和2018041 8009(博鳌)与越南和马来西亚的Y.formosana聚为一支, ML、MP 和NJ 的自展支持值为100%、99%和100%, 碱基差异为0, 遗传距离为0, 表明采集于海南省三亚市大东海和琼海市博鳌地区的样品与Y.formosana为同一物种;Y.formosana进化支与马达加斯加岛和斯里兰卡的Y.ligulata构成的进化支互为姐妹分支, 碱基差异为8bp; 来自日本的Y.tenuifolia以及来自马达加斯加岛的Y. antimo-vatae分别进化为一个分支, 碱基差异为19bp; 而来自印度尼西亚的Y.zolligeri与Vranken(来自比利时根特大学, 采集于海洋水族馆, 具体位置未知)鉴定的Y.zolligeri序列(LT969680、LT969695)分别单独进化为一个分支, 碱基差异为5bp。
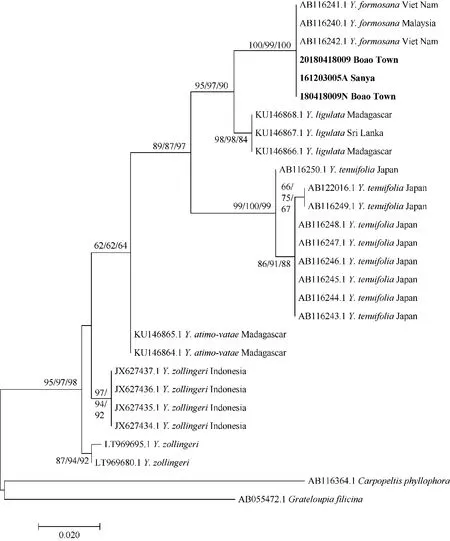
图2 基于rbcL 序列构建的最大似然树Fig. 2 Maximum likelihood tree constructed based on rbcL sequence
3 讨论
海膜科是由Bory 于1828 年建立的一个科, 目前总共报道了 38 个属。该科曾被归为隐丝藻科Cryptonemiaceae 和蜈蚣藻科Grateloupiaceae(夏邦美,2004), 但根据命名规则, Guiry(1978)将海膜科修订为有效名称。由于该科物种的形态变化较大, 关于该科完整的分类系统还未建立, 因此海膜科也被认为是最难分类的科之一(米伟男 等, 2009)。海膜科中的大多数属以辅助细胞生殖枝丛的结构特征进行区分, 包括了 5 个类型的辅助细胞生殖枝丛, 即Aeodes型、Cryptonemia型、Grateloupia型、Halymenia型和Thamnoclonium型(Chiang, 1970;Kawaguchi et al, 2002, 2004)。其中,Grateloupia型为该科内最简单的辅助细胞生殖枝丛类型, 属于该类型的有Dermocorynus、Grateloupia、Kintokiocolax、Phyllymenia和Zymurgia等5 个属。在 Kawaguchi 等(2004)对那屿藻属辅助细胞生殖枝丛开展研究后, 也将其归入到Grateloupia型中,并根据其特有的辅助细胞生殖枝丛发育方式以及基于rbcL 的系统发育分析结果, 将其命名为新属。目前, 那屿藻属在分类学上已被分类学者所接受(Boo et al, 2013; Manghisi et al, 2015; Lin et al,2020)。
本文的研究样品(161203005A、180418009N、20180418009)符合那屿藻属物种的特征(Kawaguchi et al, 2004)。在形态上, 这3 个样品的藻体均扁平,表面和边缘着生有许多小育枝, 这与该属内的Y.formosana和Y. ligulata比较相似, 然而这三者的大部分测量数据与Y.formosana和Y. ligulata存在一定的差异, 但总体来说与Y.formosana比较接近(表1)。从地理分布范围看,Y.formosana主要报道于太平洋地区(我国台湾地区和海南省, 以及日本、越南、马来西亚、菲律宾和波利尼西亚), 而Y.ligulata则主要报道于印度洋地区(马达加斯加、 斯里兰卡)。基于rbcL 序列的系统发育分析, 样品161203005A(三亚)、180418009N(博鳌)和20180418009(博鳌)与越南和马来西亚的Y.formosana聚为一支, 且基于不同方法构建系统发育树的自展支持值均较高。因此,结合形态学、地理分布以及分子序列的分析结果,采集于海南省的这3 个那屿藻属样品可鉴定为台湾那屿藻Yonaguniaformosana。
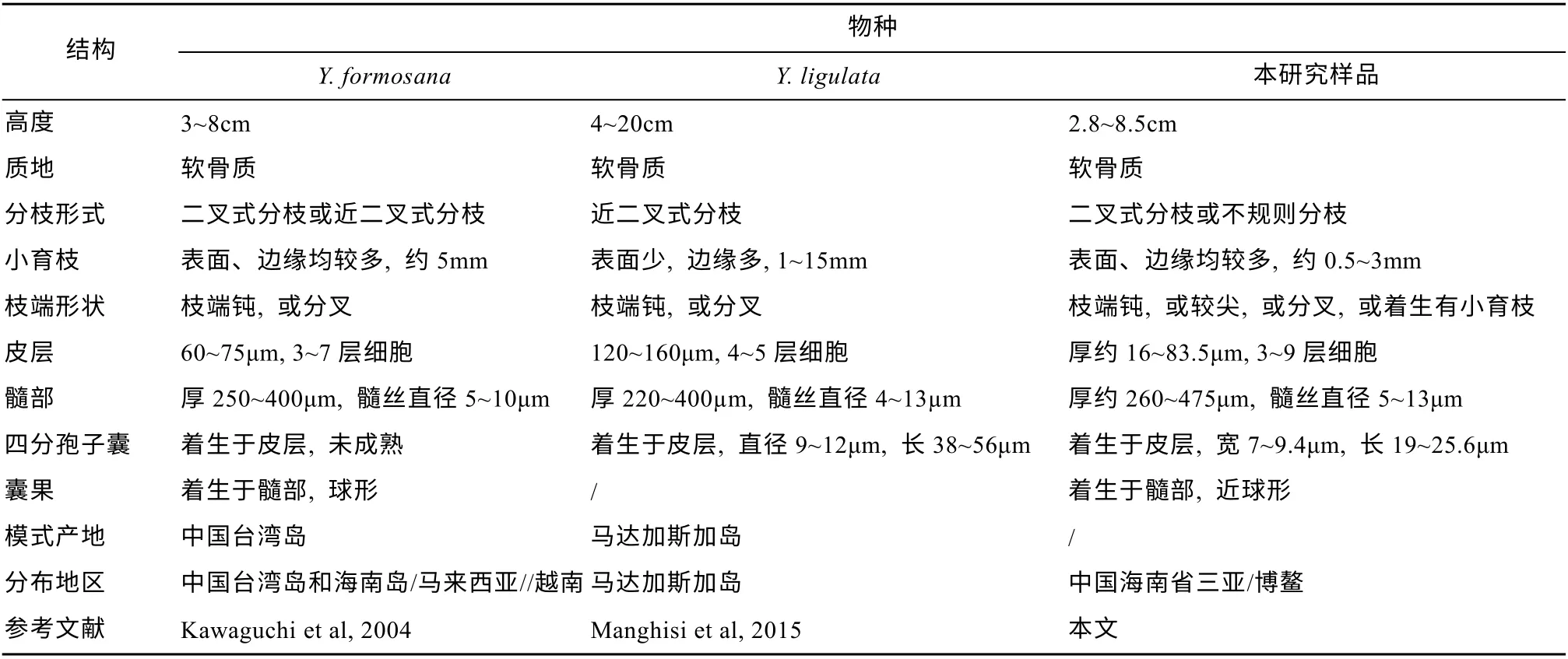
表1 台湾那屿藻、Y. ligulata 和样品的比较Tab. 1 Comparison of Y. formosana, Y. ligulata and the sample
台湾那屿藻最早被命名为Carpopeltis formosana(Okamura, 1931), Kawaguchi 等(1998)将其转移至锯齿藻属Prionitis中, 命名为Prionitis formosana; Kawaguchi 等(2004)又因建立那屿藻属Yonagunia而将P.formosana修订为Y.formosana。目前Y.formosana的命名在分类学上得到确认。但由于上述拉丁种名以及近些年来国际属种名称的快速变化, 在一定程度上导致了其中文名称的使用问题。在我国, 该种的中文名称分别出现过如下变化:台湾盾果藻Carpopeltisformosana(黄淑芳, 2000)、台湾锯齿藻Prionitisformosana(夏邦美, 2004)、台湾盾果藻Carpopeltisformosana和台湾锯齿藻Prionitisformosana(曾呈奎 等, 2005)、台湾锯齿藻Yonaguniaformosana(丁兰平 等, 2015)。根据我们的文献记录, 到2014 年, 锯齿藻属Prionitis因物种转出等原因而没有确定的记录物种, 可被视为废弃的或同物异名属, 因此我们在2015 年继续采用了锯齿藻属和台湾锯齿藻为Yonagunia和Y.formosana的中文名。然而, 近年来, 原Prionitis中的一些物种又得到确认, 导致该属废而又立, 其中文属名“锯齿藻属”应该继续保留。近年的一篇文献又将Yonagunia的中文名称命名为“与那国藻属”(赵树雨, 2016)。然而, 有评阅人认为“与那国藻属”不太符合中文的取名式样。因此本文结合评阅人的建议, 将Yonagunia和Y.formosana的中文名称分别修改为那屿藻属和台湾那屿藻。
台湾那屿藻Y.formosana的模式产地为中国台湾, Lin 等(2020)对中国台湾地区的Y.formosana进行了分子系统发育研究, 但其分子序列在数据库中尚不可查阅。其与本文分子序列是否存在差异还有待探讨。海南省东南部地区的Y.formosana与目前国内外所报道的Y.formosana在形态上比较相似,但本文样品的枝端形状比文献报道的更加多样化,藻体主要特征(分枝形式、小育枝、藻体皮层等)的大小也与Kawaguchi 等(2004)所报道的存在一定的差异, 是否为形态多样化仍有待研究。
4 结论
本研究通过对2016—2018 年采集于海南省东南部地区的那屿藻属的种类样本的形态学研究, 并结合rbcL 序列的系统发育分析, 共鉴定台湾那屿藻一个物种, 研究结果对那屿藻属的分类学研究具有一定的参考意义, 丰富了那屿藻属种类分布的地域信息, 也丰富了我国沿海大型藻类的物种多样性。
