不同立地基生枝年龄对笃斯越桔有机营养特性的影响1)
2021-12-03吴燕陈露白永超侯智霞
吴燕 陈露 白永超 侯智霞
(北京林业大学,北京,100083)
笃斯越桔(Vacciniumuliginosum. L)属杜鹃花科(Ericaceae)越橘属(Vaccinium)落叶灌木。我国笃斯越桔资源丰富,主要分布于大小兴安岭及长白山地区[1]。其果实富含花青素、类黄酮、单宁等增强人体免疫、抗衰老等功效的物质,具有较高研究价值[2-4]。但长期缺乏有效管理及保护,导致笃斯越桔植株长势较弱且产量降低[5-6]。有机营养物质对植物生长发育有重要影响,但目前关于笃斯越桔有机养分质量分数特点的研究较少,其分布特点、作用机理仍不清楚。因此了解笃斯越桔植株营养特性并采取有效抚育措施至关重要。
植物枝条年龄与营养物质质量分数密切相关,植物运输营养物质的能力影响植物生长。研究表明:枝条年龄对植物可溶性总糖、淀粉的积累有显著影响[7-8];不同类型枝条对营养储存的水平不同,叶片、枝条对营养物质利用特征不同[9]。可溶性总糖、淀粉等是重要的渗透调节物质[10-12],植物可溶性总糖优先供应处于生长中心的组织或器官[13],没被利用的可溶性总糖则分配到枝条等储藏器官[14]。可溶性总糖是植物代谢发育中能量的主要来源[15];是碳水化合物在植物体内合成、代谢、运输情况的反应[16]。
研究发现,植物基生枝营养水平、生长发育状况反应植物生长潜力,影响植物产量[17-18]。植物自身生长代谢、环境条件促使笃斯越桔植株基部根态茎上潜伏芽萌发。笃斯越桔植株形成行茎后,先后进行营养生长、生殖生长,以分枝取代的形式进行衰老、死亡、更新[19]。不同立地条件对提高笃斯越桔营养物质质量分数、果实品质等方面都有重要影响[20-22]。
研究不同立地类型、不同年龄笃斯越桔基生枝条的有机营养特点对笃斯越桔生长潜能(新梢生长量)有重要意义。本研究分析了3种立地类型不同年龄阶段笃斯越桔基生枝叶片、枝条可溶性总糖、淀粉及可溶性蛋白积累的特性;可溶性总糖、淀粉和可溶性蛋白在笃斯越桔不同年龄阶段枝干的分布积累特点;植株有机物质积累量与新梢生长量的相关性。本研究为探索笃斯越桔更新及衰老过程中有机物质变化及笃斯越桔的抚育管理提供参考;提高笃斯越桔利用价值。
1 研究地概况
研究区位于大兴安岭地区,年温差较大,属寒温带大陆性季风气候,海拔445~873 m,土壤腐殖质丰富,土壤呈酸性,pH值为4~5[23]。参照尹德洁等[1]研究的分类标准,选择水湿地、山地、塔头草甸3种立地类型为3个研究区。
2 材料与方法
2.1 样品采集
在笃斯越桔集中分布地区随机设置3个20 m×20 m样地,每个样地中选取四角及中心共5个1 m×1 m的标准样方进行生境记录(表1)及样品采集。
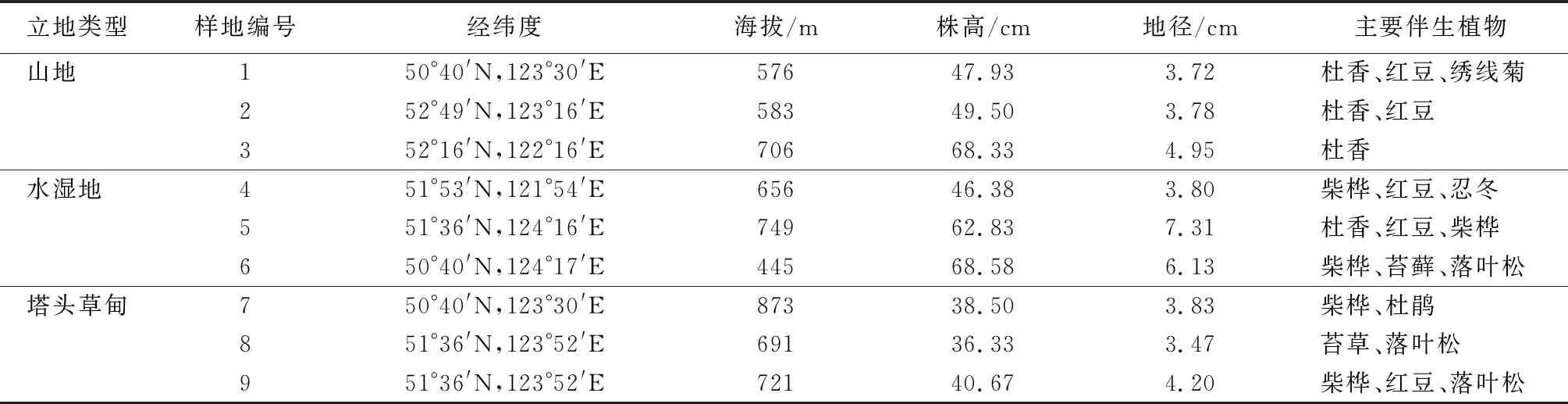
表1 各样地基本情况调查
将笃斯越桔灌丛基生枝分为1~2年(S1)、3年(S2)、4年(S3)、5年(S4)生枝、趋于衰老的多年生基生枝(枝龄>5 a,S5)、枯死枝干(S6)。本试验于2015年8月中旬进行取样,每个标准样方中随机选择20丛长势相对一致的笃斯越桔。对S1~S6不同时期基生枝叶片、枝干(叶片除外的所有枝条)进行分类采集,同一枝龄同一部位样品混合,为待测样品。叶片共采样45份,枝条共采样54份。
2.2 测定指标及方法
采集样品带回实验室后,对54个枝条样品进行新梢生长量测定,具体方法如下:每份样品随机抽取20个枝条,用卷尺分别测量其当年新生枝条的长度,即为新梢生长量,并记录。采用蒽酮比色法测定可溶性总糖质量分数;考马斯亮蓝法测定淀粉、可溶性蛋白质量分数,具体实验步骤参照《植物生理学指导》[24],并修改。
3 结果与分析
3.1 糖类物质
3.1.1 蔗糖
笃斯越桔叶片蔗糖质量分数略低于枝条,且多集中于S2~S4阶段(表2)。塔头草甸立地类型的叶片蔗糖质量分数在S1、S3~S4时期显著低于其余2种立地类型,山地类型、水湿地类型的叶片蔗糖质量分数间差异不显著(P<0.05);S2时期山地类型的叶片蔗糖质量分数显著高于水湿地、塔头草甸类型的叶片蔗糖质量分数;S5时期3种立地类型的叶片蔗糖质量分数无显著差异。3种立地类型的笃斯越桔枝条蔗糖质量分数在S1、S5时期无显著差异;S2~S3、S6时期山地类型的枝条蔗糖质量分数显著高于其余2种立地类型;S4时期山地类型的枝条蔗糖质量分数显著低于塔头草甸类型。
不同年龄的叶片中,山地类型S2时期、水湿地类型S3~S4时期积累蔗糖较多,约为S1时期的1.37~1.62倍;塔头草甸类型的蔗糖质量分数在S2、S5时期较高。不同年龄的枝条中,山地类型枝条的蔗糖质量分数最大值出现在S3时期,为11.04 mg·g-1,高于最低时期(S1)约143.31%;水湿地类型的枝条蔗糖质量分数在S4时期达到最高后,枝条年龄越大,蔗糖质量分数越低;与水湿地类型类似,塔头草甸类型的枝条蔗糖质量分数也在S4时期达到最高。
3.1.2 可溶性总糖
山地、水湿地、塔头草甸3种立地类型中,笃斯越桔不同年龄基生枝的叶片可溶性总糖质量分数明显高于枝条(表2)。S1、S5时期,水湿地、塔头草甸类型叶片可溶性总糖质量分数显著高于山地类型(P<0.05);S2、S3时期,3种立地条件时叶片可溶性总糖质量分数均无显著性差异;S4时期均差异显著。山地类型在S3时期,叶片可溶性总糖质量分数最高,S2与S4时期、S1与S5时期的叶片可溶性总糖质量分数差异不显著。水湿地与塔头草甸立地类型叶片可溶性总糖均在S4时期积累最多。
S3时期,3种立地类型的枝条可溶性总糖质量分数无显著差异;其余时期山地类型的枝条可溶性总糖质量分数均显著低于其余2种立地类型。山地类型的枝条可溶性总糖质量分数与叶片可溶性总糖质量分数分布相似,S3~S4时期积累最多;水湿地类型的枝条可溶性总糖质量分数在S2、S6时期显著低于S1、S3~S5时期;塔头草甸类型的枝条可溶性总糖质量分数在S2时期显著高于其余时期。
3.1.3 淀粉
笃斯越橘叶片淀粉质量分数略高于枝条(表2)。S4时期,叶片淀粉质量分数在3种立地类型中无显著差异;S1、S2、S3、S5时期,水湿地类型的叶片淀粉质量分数均显著高于其余2种立地类型(P<0.05)。S1、S3、S5时期山地、塔头草甸类型的叶片淀粉质量分数间均无显著差异。山地类型叶片淀粉质量分数呈标准正态分布,S3时期最高;水湿地类型叶片淀粉质量分数在营养生长时期(S1)最高,约为最低积累量(S4时期)的1.52倍;塔头草甸类型S4时期的叶片淀粉积累量最高。
水湿地类型的枝条淀粉质量分数显著高于其余2种立地类型。除S2时期外,山地与塔头草甸类型的枝条淀粉质量分数间,均无显著差异。水湿地类型的枝条淀粉积累量在S1、S3、S5时期较高,S2、S4、S6时期较低;山地、塔头草甸类型的枝条淀粉于S2~S4时期积累较多。笃斯越桔植株的淀粉积累量在水湿地类型中最高,山地类型次之,塔头草甸类型最少。笃斯越桔植株的叶片、枝条淀粉积累量受枝干年龄影响,多集中于代谢旺盛的S2~S4时期。
3.1.4 可溶性总糖淀粉质量分数比
可溶性总糖质量分数/淀粉质量分数在一定程度上反应淀粉的转化效率。本研究中,S3时期,塔头草甸类型与山地类型的叶片可溶性总糖质量分数/淀粉质量分数无显著差异(P<0.05),其余时期塔头草甸类型的叶片可溶性总糖质量分数/淀粉质量分数均显著高于其余2种立地类型。其中,山地与水湿地类型叶片可溶性总糖质量分数/淀粉质量分数差异不显著(表2)。枝条可溶性总糖质量分数/淀粉质量分数分布规律与叶片相似,S1~S6时期,塔头草甸类型的枝条可溶性总糖质量分数/淀粉质量分数均显著高于其余2种立地类型。山地、水湿地类型的枝条可溶性总糖质量分数/淀粉质量分数在S1~S5时期无显著差异,S6时期山地类型的枝条可溶性总糖质量分数/淀粉质量分数显著高于水湿地类型。
山地类型的叶片可溶性总糖质量分数/淀粉质量分数在不同时期无显著变化(P<0.05);S4时期,水湿地类型的叶片可溶性总糖质量分数/淀粉质量分数显著高于其余时期;塔头草甸类型叶片可溶性总糖质量分数/淀粉质量分数S2时期达到最高,S1与S4时期、S3与S5时期无显著差异。山地类型枝条可溶性总糖质量分数/淀粉质量分数在S1~S2阶段较低,显著低于S6时期;水湿地类型的枝条可溶性总糖质量分数/淀粉质量分数在S4时期显著高于其余时期;塔头草甸类型与水湿地的枝条可溶性总糖质量分数/淀粉质量分数的分布一致,但塔头草甸类型的枝条可溶性总糖质量分数/淀粉质量分数的最大值出现在S2时期,显著高于其余时期,其余时期的枝条可溶性总糖质量分数/淀粉质量分数间无显著差异。
3.1.5 可溶性蛋白
笃斯越桔的叶片、枝条可溶性蛋白质量分数积累状况可知,枝条的可溶性蛋白质量分数远低于叶片可溶性蛋白质量分数(表2)。S1~S3时期,水湿地类型的叶片可溶性蛋白质量分数显著高于其他2种立地类型(P<0.05);S4~S5时期,山地类型的叶片可溶性蛋白质量分数显著低于水湿地、塔头草甸类型的叶片可溶性蛋白质量分数,且水湿地、塔头草甸类型的叶片可溶性蛋白质量分数间无显著差异。S5时期,3种立地类型的枝条可溶性蛋白质量分数无显著差异,其余时期水湿地类型枝条可溶性蛋白质量分数均显著高于其他2种立地类型。
山地、水湿地类型的叶片可溶性蛋白质量分数与枝条年龄呈正相关,均在S5时期达到最大值,显著高于对应S1时期的32.13%;塔头草甸类型的叶片可溶性蛋白质量分数在S4~S5时期显著高于其他时期,且在S4时期达到最大值。笃斯越橘枝条可溶性蛋白质量分数在植物代谢旺盛时期(S1~S4时期)较高。山地、塔头草甸类型枝干可溶性蛋白质量分数在S6时期最低;水湿地类型枝干可溶性蛋白在营养生长期(S1)积累最多,显著高于其他时期,在S6时期最低。笃斯越桔中可溶性蛋白积累多集中于营养生长期(S1)及大量结果期(S2~S4),且叶片可溶性蛋白质量分数远高于枝干。立地类型对笃斯越橘可溶性蛋白质量分数也有所影响,水湿地类型可溶性蛋白积累较多,塔头草甸类型次之,山地类型最少。

表2 不同立地类型、年龄的枝条及叶片有机营养物质分析
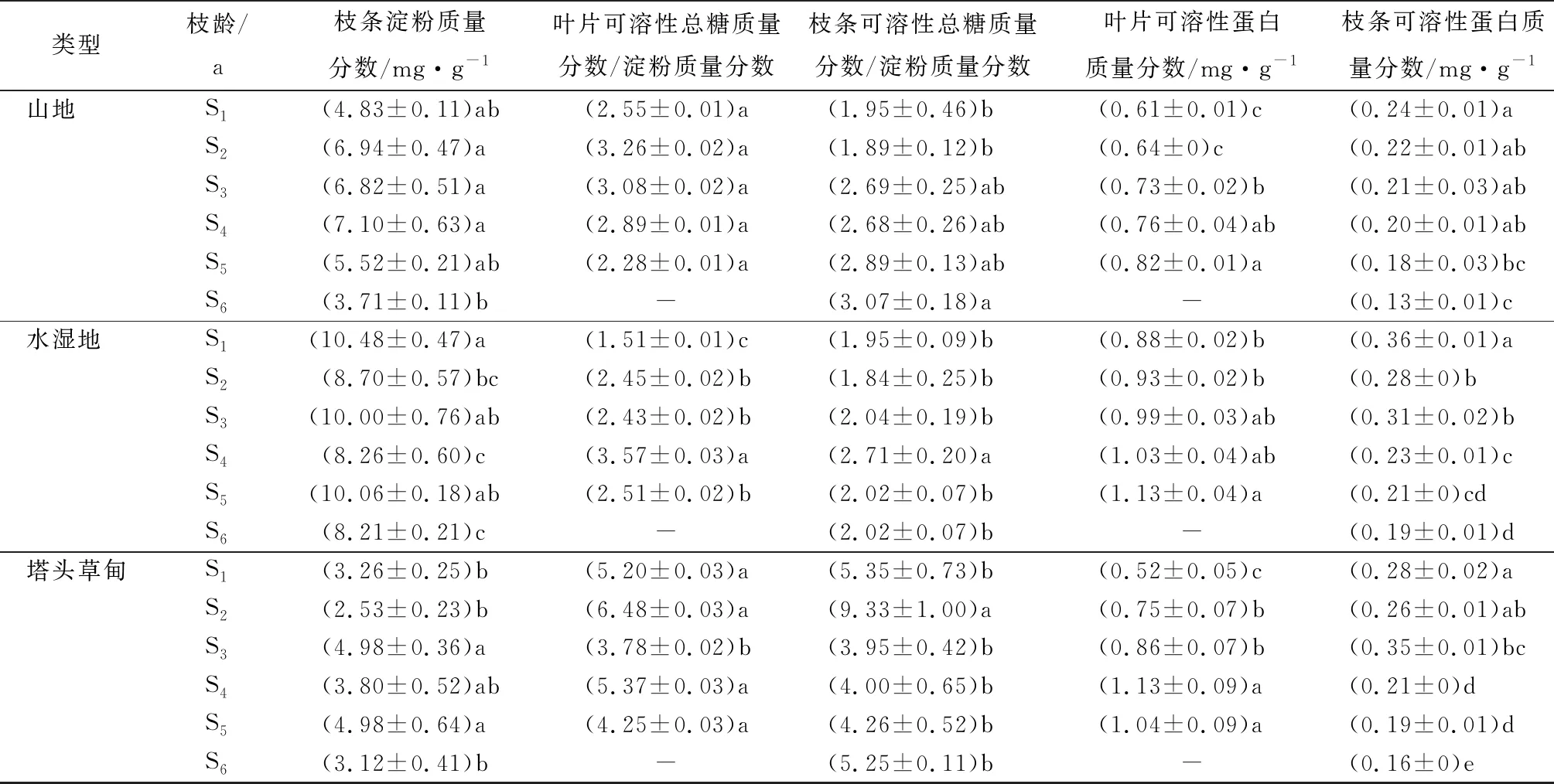
类型枝龄/a枝条淀粉质量分数/mg·g-1叶片可溶性总糖质量分数/淀粉质量分数枝条可溶性总糖质量分数/淀粉质量分数叶片可溶性蛋白质量分数/mg·g-1枝条可溶性蛋白质量分数/mg·g-1山地S1(4.83±0.11)ab(2.55±0.01)a(1.95±0.46)b (0.61±0.01)c (0.24±0.01)a S2(6.94±0.47)a(3.26±0.02)a(1.89±0.12)b(0.64±0)c(0.22±0.01)abS3(6.82±0.51)a(3.08±0.02)a(2.69±0.25)ab(0.73±0.02)b(0.21±0.03)abS4(7.10±0.63)a(2.89±0.01)a(2.68±0.26)ab(0.76±0.04)ab(0.20±0.01)abS5(5.52±0.21)ab(2.28±0.01)a(2.89±0.13)ab(0.82±0.01)a(0.18±0.03)bcS6(3.71±0.11)b -(3.07±0.18)a -(0.13±0.01)c水湿地S1(10.48±0.47)a(1.51±0.01)c(1.95±0.09)b(0.88±0.02)b(0.36±0.01)aS2(8.70±0.57)bc(2.45±0.02)b(1.84±0.25)b(0.93±0.02)b(0.28±0)bS3(10.00±0.76)ab(2.43±0.02)b(2.04±0.19)b(0.99±0.03)ab(0.31±0.02)bS4(8.26±0.60)c(3.57±0.03)a(2.71±0.20)a(1.03±0.04)ab(0.23±0.01)cS5(10.06±0.18)ab(2.51±0.02)b(2.02±0.07)b(1.13±0.04)a(0.21±0)cdS6(8.21±0.21)c -(2.02±0.07)b -(0.19±0.01)d塔头草甸S1(3.26±0.25)b(5.20±0.03)a(5.35±0.73)b(0.52±0.05)c(0.28±0.02)aS2(2.53±0.23)b(6.48±0.03)a(9.33±1.00)a(0.75±0.07)b(0.26±0.01)abS3(4.98±0.36)a(3.78±0.02)b(3.95±0.42)b(0.86±0.07)b(0.35±0.01)bcS4(3.80±0.52)ab(5.37±0.03)a(4.00±0.65)b(1.13±0.09)a(0.21±0)dS5(4.98±0.64)a(4.25±0.03)a(4.26±0.52)b(1.04±0.09)a(0.19±0.01)dS6(3.12±0.41)b -(5.25±0.11)b -(0.16±0)e
3.2 有机营养与新梢生长量的相关性
简单相关分析结果显示(表3),笃斯越桔新梢生长量变化与叶片糖类物质、可溶性蛋白呈不显著负相关、与枝条蔗糖呈显著负相关(r=-0.517*)、与枝条可溶性蛋白呈极显著正相关(r=0.699*);叶片可溶性总糖、淀粉、可溶性总糖质量分数/淀粉质量分数与枝条对应指标均呈显著或极显著正相关;叶片可溶性总糖与叶片淀粉、可溶性蛋白呈显著正相关。

表3 笃斯越桔叶片、枝条有机营养与新梢生长量的相关性
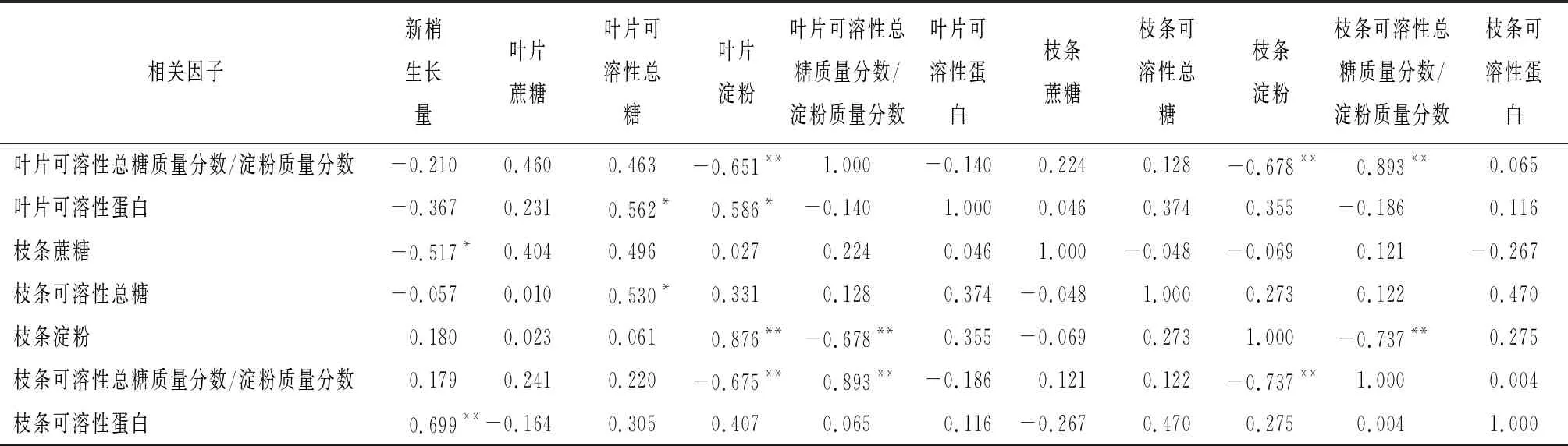
续(表3)
3.3 立地条件对笃斯越桔有机物质积累的影响
立地是植物的生长发育所需营养物质的直接来源。单变量线性模型分析显示:立地类型对笃斯越桔叶片淀粉、叶片可溶性蛋白、枝条蔗糖、枝条淀粉、枝条可溶性蛋白在不同年龄基生枝条上的分布,均有显著或极显著影响(表4)。

表4 不同立地条件下笃斯越桔有机物质积累的单变量方差分析
3.4 枝条新梢生长量的影响因子
不同立地类型枝条新梢生长量的影响因子不同(表5)。山地类型枝条新梢生长量主要受枝条年龄、叶片蔗糖质量分数、叶片可溶性总糖质量分数/淀粉质量分数、枝条可溶性蛋白质量分数的影响;水湿地类型枝条新梢生长量主要受枝条年龄、枝条可溶性总糖质量分数、叶片淀粉质量分数的影响;塔头草甸类型枝条新梢生长量主要受枝条年龄、叶片可溶性总糖质量分数/淀粉质量分数、枝条可溶性总糖质量分数的影响。由表5可知,枝条年龄对新梢生长量的影响最大,呈负相关。不同立地条件下,叶片可溶性总糖质量分数/淀粉质量分数均对新梢生长量产生影响。山地类型的叶片可溶性总糖质量分数/淀粉质量分数与新梢生长量呈负相关;水湿地、塔头草甸类型的叶片可溶性总糖质量分数/淀粉质量分数与新梢生长量呈正相关。

表5 影响枝条新梢生长量的因子筛选及回归方程的建立
4 结论与讨论
4.1 不同年龄枝条有机营养的积累特性
8月上旬处于笃斯越桔果实成熟期,此时新梢增长基本停止,功能叶片同化水平高,各部位器官的同化物分配达到较均衡的状态[25]。以该时期笃斯越桔糖类物质及可溶性蛋白作为参考对象,估测笃斯越桔不同年龄阶段枝条的代谢水平。
糖是植物生长发育必不可少的物质之一,蔗糖是叶片光合产物向其他器官运输的主要非还原性糖形态[26]。本研究中笃斯越桔叶片蔗糖质量分数略低于枝干,原因是果实成熟过程中,植株营养生长趋于停止,生殖生长加强,此时叶片光合作用产生的蔗糖会优先供应至代谢旺盛部位[27]。S2~S4阶段笃斯越桔枝干叶片蔗糖质量分数高于S1、S5时期(除塔头草甸类型外),推测是由于S1时期枝干主要进行营养生长,枝干上着生叶片光合作用产生的碳水化合物以非蔗糖形式运输至植株其他部位;S5时期枝干已趋于老化,枝干着生叶片的光合作用有所下降。
可溶性总糖与可溶性蛋白一定程度反映植物对环境的适应性[16,28-29],可溶性蛋白质量分数变化是植物生长过程中对环境适应性的体现[30]。本研究中,塔头草甸类型笃斯越桔叶片、枝干的可溶性总糖质量分数最高,水湿地类型次之,山地类型最少。塔头草甸立地类型多分布于沼泽地,含水量极高[1],而笃斯越桔仍能正常生长,说明该立地条件下笃斯越桔耐涝能力高于其他立地。克热曼等[31]研究不同立地下葡萄的果实品质时也发现:相比其他立地,含水量高的壤土果园植物的枝条可溶性总糖质量分数要高于其余2种含水量较低的土壤。一定时期内,叶片、枝干可溶性总糖质量分数随枝条年龄增加而增加。有研究表明,大叶女贞枝条可溶性总糖质量分数随树龄增加而增加;油茶在一定树龄内,抗逆性随树龄增加而增加,100 a树龄油茶抗逆性有所下降[5,32]。笃斯越桔在代谢旺盛时期,体内有机营养质量分数较高,植株整体状态较好。
淀粉属于储藏类多糖,由光合作用直接合成或通过蔗糖转化[33]。笃斯越桔水湿地类型的叶片、枝条淀粉质量分数均高于其他立地类型,山地类型次之,塔头草甸类型最少。推测立地类型影响淀粉在笃斯越桔树体内的积累。王丹等[34]发现低温胁迫可促使结缕草体内淀粉水解,导致体内可溶性总糖增加,淀粉减少。可溶性总糖质量分数/淀粉质量分数可作为植物抗逆性的参考指标[29]。本研究中,相同气候条件下,塔头草甸类型的可溶性总糖质量分数/淀粉质量分数显著高于其他立地类型,说明相比其他立地类型,塔头草甸类型的笃斯越桔生长状态更好。
4.2 影响新梢生长量的因素
立地土壤环境因素是影响植物营养物质质量分数的重要因素之一[35]。已有研究表明,立地类型明显影响笃斯越桔生长发育[20]、树体矿质元素质量分数[21]、果实品质及内生菌种类[36]。本研究中,不同立地条件的新梢生长量均与可溶性总糖质量分数/淀粉质量分数有关,在山地类型中呈负相关,其余2种立地类型均呈正相关。有研究表明,水湿地类型的新梢生长量较高,山地类型相对较低[37]。山地类型的笃斯越桔矿质元素,影响果实品质的总酚、类黄酮、可溶性总糖等均相对较低[38],推测山地条件对笃斯越桔生长影响比较特殊。
与矿质元素来源不同,有机物质来自于植物本身的同化,是植物的生长发育所必备的能源物质,两者均优先供应生长中心[26]。碳水化合物为植物体内的能源物质,在叶片中通过光合作用产生,继而经由韧皮部运送至枝条、根部,代谢旺盛的部位是同化产物的优先分配点[26]。本研究中,笃斯越桔S1~S4阶段新梢生长量变化与叶片、枝条中糖类物质及可溶性蛋白变化的差异是因为物种自身的遗传因素;在结果盛期,笃斯越桔叶片同化产物糖类、蛋白类物质在供应新梢生长的同时还会就近运输至果实,用于果实的生长、代谢。随着枝条不断生长,生长中心逐渐转移,枝条所能利用的能源物质减少,在本研究中是引发新梢生长量减少的原因之一。
