华南上泥盆统法门阶哲杜斯贝动物群的埋藏学和居群动力学特征*
2021-12-02袁志伟秦树健杨润宇李辰卿孙元林
李 彤 董 琳 袁志伟 凌 坤 甯 濛 秦树健 杨润宇 刘 威 聂 婷 李辰卿 孙元林
1北京大学地球与空间科学学院,北京 100871 2成都理工大学沉积地质研究院,四川成都 610059
1 概述
哲杜斯贝属(Dzieduszyckia)是古生代小嘴贝类中个体最大的种类之一,以双中槽和粗壮放射状壳线为特征。它具有广泛却不连续的地理分布,在美国、波兰、墨西哥、摩洛哥、南乌拉尔地区、哈萨克斯坦和中国华南地区的法门阶中都有分布(Nalivkin,1947;Rozman,1962;Biernat,1967;Cloud and Boucot,1971;Ageretal., 1976;鲜思远和江宗龙,1978;Nolletal., 1984;Baliński and Biernat,2003;Peckmannetal., 2007;Nieetal., 2016)。哲杜斯贝石通常只发现在由单一属种构成的、个体密集保存的腕足动物群中。Ager等(1976)从其单调的属种组成和个体极为密集的保存状态推断,哲杜斯贝很可能生活在极为特殊的、只适合这种腕足动物生活的环境中。Campell和 Bottjer(1995)则推论,哲杜斯贝可能是生活在与海底热泉喷口(hydrothermal vent)或冷泉(cold-seep)相关的环境中。同时,由于哲杜斯贝在形态结构上与一些被认为生活在海底热泉喷口—冷泉环境中的小嘴贝类(如石炭纪的Ibergirhynchia、三叠纪的Halorella和白垩纪的Peregrinella)十分相似,故推断它们可能属于一类长期栖息在海底热泉喷口或冷泉环境中的、免受大绝灭事件影响的小嘴贝类支系(Campell and Bottjer,1995;Sandy,1995;Gischleretal., 2003;Peckmannetal., 2007)。这一假说得到了来自北美地区哲杜斯贝化石保存的直接证据支持(Nolletal., 1984;Dubé,1988;Torresetal., 2003;Campbell,2006)。此外,Peckmann等(2007)对摩洛哥1个哲杜斯贝化石产地剖面的地球化学研究发现,在哲杜斯贝壳体表面和内部的带状/葡萄状胶结物中存在无机碳碳同位素负异常,由此推测这些带状/葡萄状胶结物的成因可能与甲烷渗漏相关,并结合围岩中丝状藻席的存在,认为该剖面的哲杜斯贝生活在透光带之下、有甲烷冷泉频繁活动的环境中。然而,Baliński和Biernat(2003)的研究则对哲杜斯贝生活环境的海底热泉喷口/冷泉假说提出质疑,他们对波兰和摩洛哥两地化石壳体样品以及附着在化石表面及内部的围岩样品进行了碳氧同位素研究,并未发现指示冷泉环境的同位素信号。Peckmann等(2007)将研究结果上的差异归结为2种可能性: 一是Baliński和Biernat(2003)可能没能采到与甲烷渗漏有关样品;二是Baliński和Biernat(2003)的研究材料可能本身就不存在与甲烷冷泉有关的沉积记录。文献中记载描述的摩洛哥一些化石点的化石产出状态,如成层分布和典型的浅水沉积环境等(Ageretal., 1976;Walliseretal., 2000),亦与冷泉假说相矛盾,推测这类腕足动物的生活环境可能并不局限于有热液活动或甲烷渗漏的环境中,它很可能比大多数仅限于生活在现代甲烷冷泉环境中的生物种类具有更大的生态可塑性(ecological plasticity)。
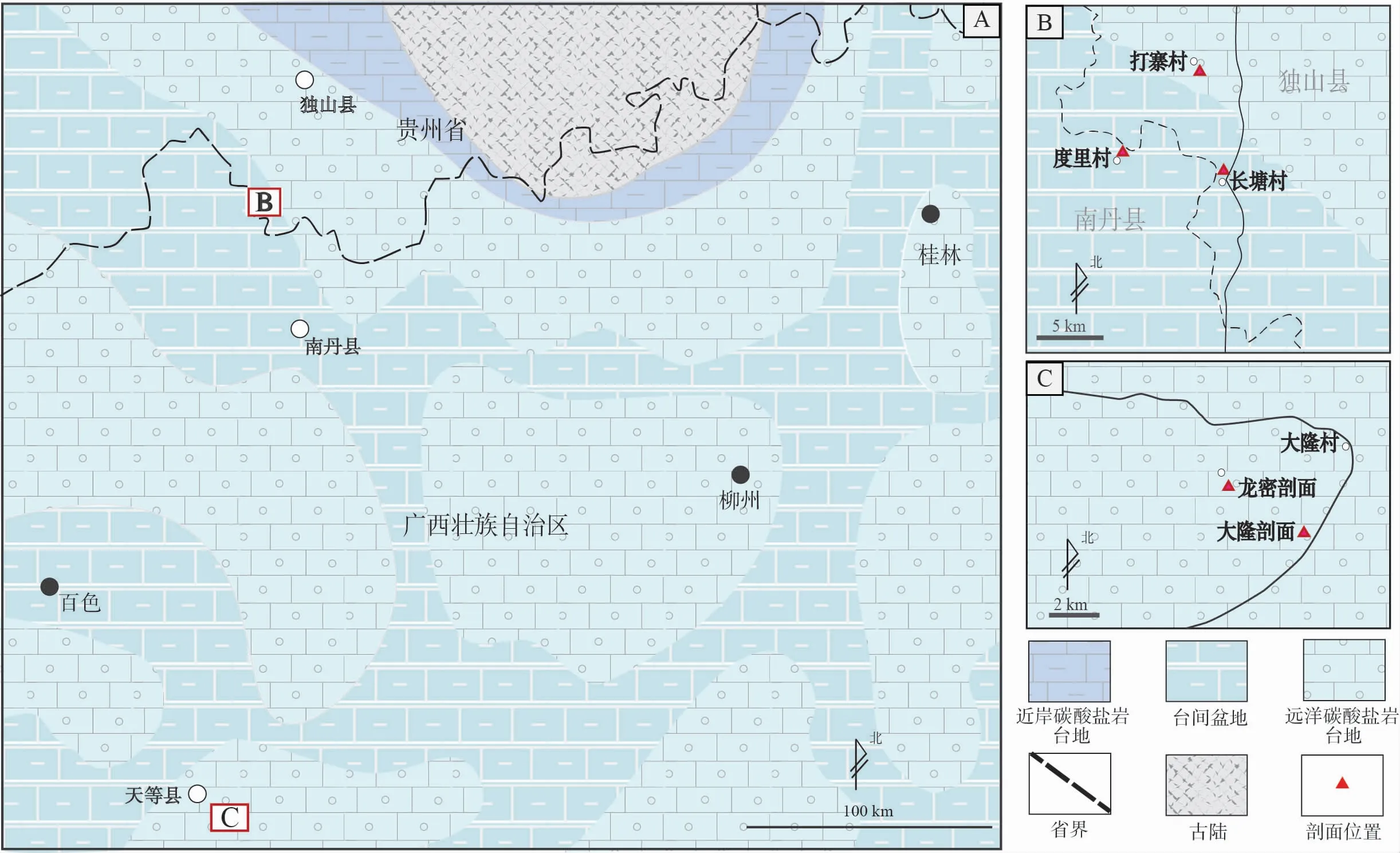
图 1 黔桂地区晚泥盆世古地理略图和研究剖面位置(修改补充自Nie et al., 2016)Fig.1 Sketched palaeogeographic map of the Late Devonianin in Guizhou-Guangxi area and location of study sections(modified from Nie et al., 2016)
华南地区保存有非常丰富的哲杜斯贝化石记录。前人曾在贵州南部的贵定大窑、长顺代化和独山甲莲塘等地“南丹型”沉积的上泥盆统代化组(=五指山组)中有过记述(孙云铸和沈耀庭,1965;鲜思远和江宗龙,1978;鲜思远等,1980)。依据独山甲莲塘和贵定大窑的有限材料,鲜思远和江宗龙(1978)命名描述了4个种:DzieduszyckiaoblongaXian、D.elongataXian、D.xiasiensisJiang 和D.acutiplicataXian,并将此属归入到无窗贝目。长顺代化的材料尚未见正式的描述报道,只在文献里以Leiorhnchusbaschkirica或Eoperegrinellabaschkirica或Dzieduszyckiabaschkirica被提及(如孙云铸和沈耀庭,1965;鲜思远等,1980;侯鸿飞等,1985)。在长顺代化剖面上,该类化石产于含典型的法门阶上部菊石Clymenia和Wocklumeria化石层之下10余米处(孙云铸和沈耀庭,1965;阮亦萍,1981)。依据这些化石记录,侯鸿飞和王士涛(1988)认为这一腕足动物群(当时以Eoperegrinella为属名)代表了1个生活在黔桂地区相对远岸深水环境中、属种单调的动物群,时代为法门期晚期(菊石Clymenia带)。最近,Nie 等(2016)报道了在黔桂交界和广西南部4个剖面(贵州独山县打寨剖面和长塘剖面,广西南丹县度里剖面和天等县大隆剖面)(图 1)上都有新的发现。在这些剖面中,均发育多层由哲杜斯贝壳体密集堆积构成的介壳层。基于牙形石生物地层学研究结果和所采集的丰富化石材料,发现哲杜斯贝在华南地区分布于法门阶的下部,而非上部;此外,从沉积环境上来说,哲杜斯贝不但在“南丹型”的相对深水台间盆地相中有分布,在“象州型”的浅水碳酸盐岩台地环境中亦有分布。但是,前人的研究中缺乏有关化石层的沉积学和埋藏学详细信息,因此尚无法判断这2种不同沉积相环境是否都代表哲杜斯贝生存的原生环境。针对这一问题,笔者在Nie 等(2016)所研究的4个剖面和华南1处新发现的剖面上(图 1)开展哲杜斯贝的沉积学和埋藏学研究;同时,也对原地埋藏的哲杜斯贝进行居群生态学的探讨,以期对该属种的生态学、埋藏学有进一步的认识。
2 地质背景和研究剖面
加里东运动导致华南板块整体隆升。泥盆纪初,华南大部分地区主要遭受风化剥蚀作用,只有广西东南地区存在残余钦防海槽。随后,以广西为中心的广大华南地区逐渐进入陆表浅海环境,海相泥盆系广泛发育(侯鸿飞和王士涛,1988;Chenetal., 2001a)。伴随海侵范围的不断扩大,在黔桂地区的海相泥盆系从早泥盆世埃姆斯期中晚期开始出现“象州型”和“南丹型”2种不同的沉积相分异(钟铿等,1992),反映了由同沉积断裂所控制的碳酸盐岩台地和台间盆地相间排列的沉积古地理格局(图 1-A)(Chenetal., 2001a,2001b)。这种沉积古地理格局一直延续到石炭纪初。在广西境内,上泥盆统法门阶远岸碳酸盐岩台地相(“象州型”)沉积主要由厚逾数百米至上千米的厚层白云质鲕粒—似球粒灰岩和藻屑灰岩构成,称作融县组;台间盆地相(“南丹型”)沉积由厚约百米的中薄层泥质条带灰岩和扁豆状灰岩构成,称为五指山组(相当于贵州的代化组+王佑组)。文中研究的5个剖面中,长塘和度里剖面属于台间盆地相,大隆、打寨和龙密剖面属于碳酸盐岩台地相(图 1-A)。
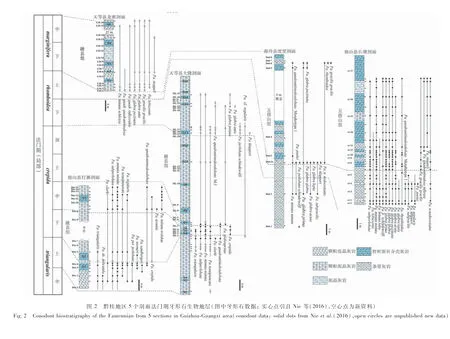
长塘剖面位于贵州省独山县麻尾镇西北约5 km的长塘村内,度里剖面位于长塘剖面以西约9 km的下度里村内(图 1-B),行政上隶属于广西南丹县六寨镇。这2个剖面均发育多层哲杜斯贝介壳层,保存在由中薄层泥质条带灰岩和扁豆状灰岩构成的五指山组中。对长塘剖面的实地踏勘核实,原16层之上的17层仍然是哲杜斯贝介壳层,整个剖面上至少有7层哲杜斯贝介壳层,单层厚度从20~30 cm到3 m不等;度里剖面可见7层20~100 cm厚的哲杜斯贝介壳层(图 2)。据Nie等(2016)的研究,这2个剖面上的哲杜斯贝介壳层均位于法门阶牙形石上rhomboidea带内。
大隆剖面位于广西南部天等县城东南约7 km的大隆村附近,龙密剖面为新发现的剖面,位于大隆剖面以西3.5 km处(图 1-C)。大隆和龙密剖面上的融县组均主要发育厚层鲕粒灰岩和藻团块亮晶灰岩。在大隆剖面上,笔者在Nie等(2016)所记述的3层化石层之下和之上又新发现了多层哲杜斯贝介壳层,共计有16层。介壳层单层厚度从20 cm到2 m不等,含哲杜斯贝介壳层的层段出露厚度约150 m(图 2)。据Nie等(2016)和笔者新的牙形石分析结果,化石层位于牙形石上triangularis带至rhomboidea带。龙密剖面含哲杜斯贝介壳层的层段出露厚约100 m,中间约有25 m厚的地表植被掩盖,未出露良好地层。共发育5层哲杜斯贝介壳层(图 2),介壳层单层厚度从1 m到3 m不等。牙形石分析结果表明,化石层位于牙形石下marginifera带内(图 2)。
3 研究方法和材料
采用野外地质锤破碎和室内台钳挤压相结合的方法采集腕足化石标本。使用气针和修理针对标本进行修理,剔除标本上残留的围岩,得到共计1272枚哲杜斯贝标本,分别是从大隆剖面S4-S6层介壳层中获得的298枚、打寨剖面的上化石层获得的536枚和龙密剖面第4层化石层S4获得的438枚较为完整的标本。采用游标卡尺(精度0.02 mm)对腕足标本进行度量,并对度量数据进行统计分析。
选取典型标本,抛光制作光面并拍照,观察沉积特征和哲杜斯贝化石埋藏特征。在哲杜斯贝介壳层和非介壳层选取典型位置制作薄厚片,在Nikon eclipse LV100POL偏光显微镜下进行薄片观察和拍照,记录其岩性、沉积相和化石组合等特征。
4 沉积和埋藏特征
4.1 台间盆地相剖面
4.1.1 沉积特征
长塘和度里剖面位于独山和南丹县交界处,在晚泥盆世法门期处于台间盆地环境(图 1)。哲杜斯贝介壳层发育在中薄层泥质条带灰岩和扁豆状灰岩构成的五指山组中。在长塘和度里剖面中,哲杜斯贝介壳灰岩与非介壳层灰岩互层产出。其中,长塘剖面发育哲杜斯贝介壳层至少7层,单层最厚可达3 m;度里剖面发育7层哲杜斯贝介壳层(在第4-5层之间有近4 m的掩盖)。2个剖面上含哲杜斯贝介壳层的地层总厚度均在10 m左右(图 2)。在介壳层内部,靠近中部的介壳密集堆积,靠近底部和顶部位置的哲杜斯贝介壳相对稀疏,介壳呈现出由紧密排列到逐渐稀疏、再过渡到围岩的特征。哲杜斯贝壳体之间填充泥晶灰岩,少量泥晶灰岩发生重结晶变成亮晶灰岩。非介壳层主要以泥晶灰岩为主。介壳层围岩与非介壳层泥晶灰岩的岩性特征一致。
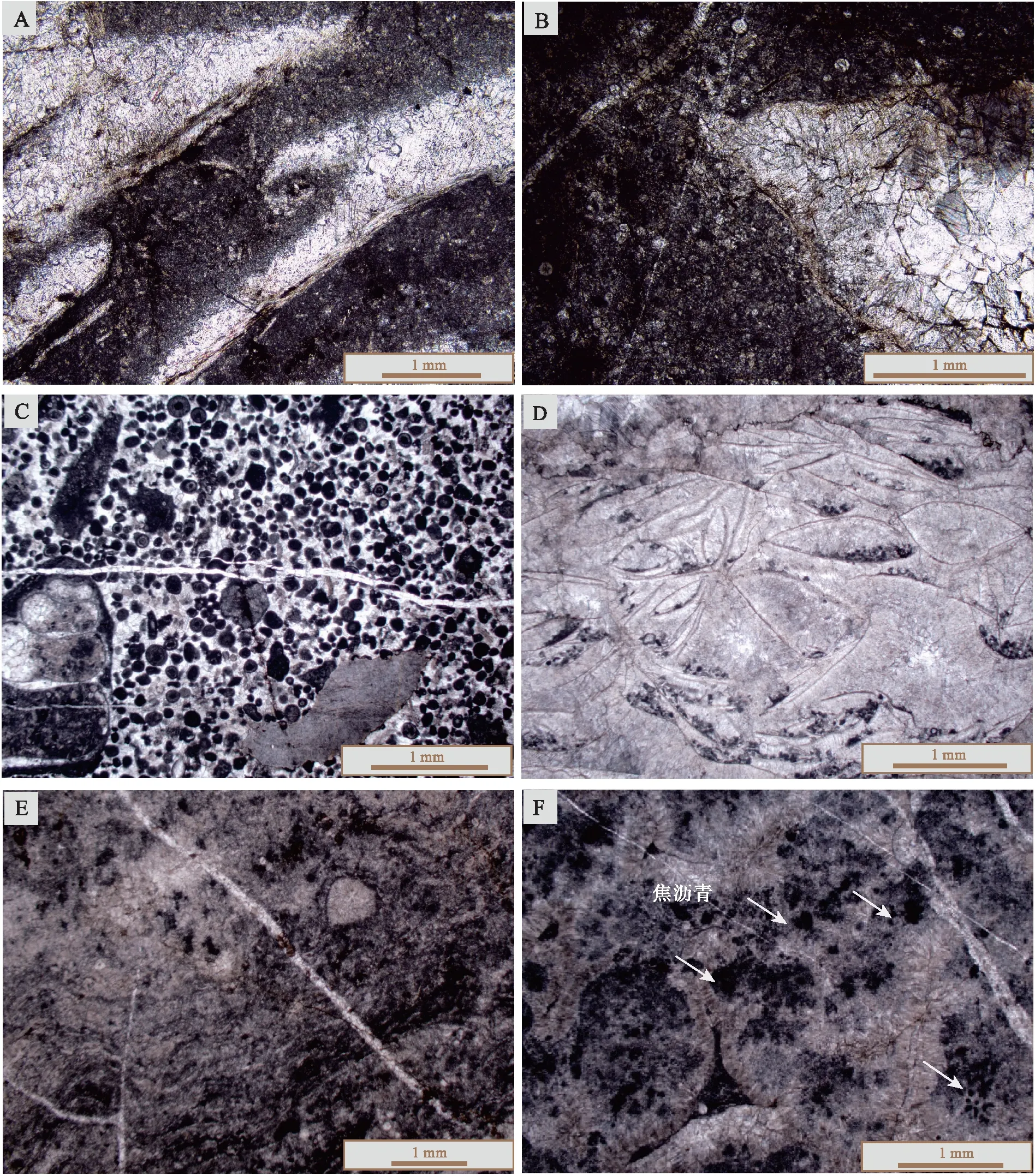
2个剖面沉积微相观察的结果基本相同,均表现出如下的特征: 哲杜斯贝介壳的钙质单瓣壳壳体在成岩的过程中被亮晶方解石交代(图 3-A,3-B)。壳体内部的有机质在哲杜斯贝死亡后被分解,形成空腔。亮晶方解石世代胶结,填充空腔(图 3-B,3-D)。介壳层中哲杜斯贝壳体之间为暗色泥晶灰岩所填充(图 3-A,3-B,3-D,3-E)。部分泥晶灰岩围岩沿单瓣壳一侧重结晶形成簇状亮晶方解石(图 3-D,3-E)。介壳层的泥晶灰岩围岩中无哲杜斯贝壳体以外的生物碎屑。
非介壳灰岩层均以泥晶灰岩为主,长塘剖面上生屑含量为5%左右(图 3-C),生屑类型包括腕足动物、棘皮动物和介形类等。度里剖面上的生屑含量不超过3%,包括介形类、腕足、腹足和极少量珊瑚碎屑。非介壳层泥晶灰岩与介壳层围岩中的泥晶灰岩十分相似,只是非介壳层泥晶灰岩中局部层位出现黄铁矿颗粒,少量发生白云岩化(图 3-F)。
4.1.2 哲杜斯贝埋藏保存特征
度里和长塘剖面介壳灰岩中,哲杜斯贝壳体密集保存(图 4-A;图 5-A,5-B),壳体在介壳层中占比达80%。哲杜斯贝壳体之间围岩为泥晶灰岩,保存的壳体以单瓣壳和壳体碎片为主。长塘剖面介壳层中单瓣壳含量约占壳体总量的80%以上,两壳铰合的完整个体含量不足20%;度里剖面介壳层单瓣壳含量占壳体总量的60%~70%,两壳铰合的完整个体占30%~40%(图 5-A,5-B),指示两地大多数壳体经历了搬运作用并遭到破坏,从而以单瓣壳保存为主。完整的个体随机散布在介壳层内,个体和个体之间不紧密接触,壳喙指向随机,平躺或直立(图 5-A,5-B),显示出壳体很可能经历了快速搬运并埋藏,并无经流水搬运后定向的特征。此外,度里剖面介壳层样品的纵切光面上局部可见明显的沉积韵律特征, 即具有壳体大小分选递变的层和只含有零星分布壳体的泥晶灰岩层互层(图 5-A),这种类似于鲍马序列(A+E)的沉积特征印证了重力流搬运的方式。因此,在华南地区台间盆地相中出现的哲杜斯贝介壳层属于异地埋藏成因。

A,B—长塘剖面,保存在泥晶灰岩中的哲杜斯贝,多为单瓣壳,样品ct-6和ct-14;C—长塘剖面,非介壳层泥晶灰岩,样品ct-4; D,E—度里剖面,保存在泥晶灰岩中的哲杜斯贝,多为单瓣壳,样品dul-3a和dul-3;F—度里剖面,非介壳层泥晶灰岩,样品dul-6。 所有照片均为单偏光图 3 黔桂地区长塘和度里剖面法门阶哲杜斯贝介壳层和围岩的镜下沉积特征Fig.3 Microscopic sedimantary features of the Famennian Dzieduszyckia coquinas and surrounding limestone from Changtang and Duli sections in Guizhou-Guangxi area

A—度里剖面,示密集堆积的介壳;B—长塘剖面,示密集堆积的介壳;C—打寨剖面,介壳层与非介壳层分界,上部为介壳层, 壳体密集堆集;D—打寨剖面,介壳层,示沿层面分布的完整壳体,壳体密集堆集,部分壳体因挤压而变形图 4 黔桂地区长塘和打寨剖面法门阶哲杜斯贝介壳层宏观沉积特征Fig.4 Macroscopic sedimentary features of the Famennian Dzieduszyckia coquinas from Changtang and Duli sections in Guizhou-Guangxi area

A—度里剖面,泥晶灰岩,壳体呈层状韵律性堆积,dul-5;B—长塘剖面,壳体紧密堆积,可见鸟眼构造,ct-17;C—大隆剖面, 壳体紧密堆积,围岩为泥晶灰岩,DL-3;D—打寨剖面,壳体完整并呈紧密堆积,壳体内部重结晶,壳体之间呈亮晶胶结,Dtz-S2图 5 黔桂地区法门阶哲杜斯贝介壳灰岩的纵切光面特征Fig.5 Vertically polished slabs features of the Famennian Dzieduszyckia coquina in Guizhou-Guangxi area
4.2 台地相剖面
4.2.1 沉积特征
打寨、大隆和龙密剖面均位于浅水碳酸盐岩台地之上(图 1),介壳灰岩与非介壳灰岩互层产出(图 4-C)。风化面和光面显示,浅水台地相的部分哲杜斯贝介壳灰岩与非介壳灰岩之间截然分界,两者间不存在过渡变化特征(图 4-C)。其中,打寨剖面上出露巨厚介壳层(厚度约2.5 m),偶夹厚2~3 cm的薄层灰岩,灰岩的出现可能指示短期环境突变或扰动,致使哲杜斯贝的生长埋藏暂时中止,之后继续繁盛。
打寨、大隆和龙密3个剖面上的介壳灰岩普遍呈层状产出。以最为典型的打寨剖面为例: 第1、2介壳灰岩层在横向上广泛延展,至少有20 m呈现稳定层状出露;第3介壳灰岩层在横向上延展,至少有50 m呈层状产出。大隆剖面和龙密剖面介壳灰岩同样表现为层状产出,但受露头出露条件所限,未能追踪到其完整的分布范围。3个剖面中,哲杜斯贝壳体可占层状产出的介壳灰岩总体积的95%(图 4-C,4-D)。介壳灰岩层底部5~10 cm主要由破碎的腕足类壳体组成,少量完整壳体散布其间,腕足类碎片的长轴多平行层面分布。介壳灰岩层的其余部分几乎完全由完整的腕足壳体紧密堆积而成。壳体含量近90%,无明显的定向性,多以腹壳在下、背壳在上的方式保存,腹喙紧靠其下相邻的个体(图 4-D),反映了正常的以肉茎固着生活状态。哲杜斯贝壳体之间多由亮晶方解石胶结,偶有泥晶灰岩充填。层状介壳灰岩中,除哲杜斯贝外,未见其他后生生物的化石。部分介壳层中,如大隆剖面第8介壳层和龙密剖面第3介壳层,可观察到哲杜斯贝集群呈现透镜状产出,透镜体最厚部分可达2~3 m,侧向延伸2 m以上。透镜体中心化石密集,哲杜斯贝壳体含量可达85%~95%,透镜体边缘化石则逐渐稀少至消失。透镜状介壳层的出现可能受到幼虫固着的影响,在这一时期透镜体中心的幼虫密度较高,固着率增强,在平面上的展布不均匀。相比于台间盆地相剖面,台地相剖面中介壳层壳体含量更高,完整壳体含量也更高(对比图 5-A,5-B与图 5-C,5-D)。
打寨、大隆和龙密3个剖面沉积微相类似,介壳层为哲杜斯贝介壳灰岩。哲杜斯贝壳体内部充填亮晶方解石,簇状方解石从壳体向内生长,中心为片状方解石,晶体颗粒大(图 6-B)。约20%的哲杜斯贝壳体内部有粒泥灰岩残留,充填在壳腔的下部。粒泥灰岩中藻屑和藻团块含量为10%~30%,藻屑主要为直径0.05 mm左右的钙球,钙球内部有重结晶现象。介壳层围岩为泥晶灰岩或亮晶灰岩,哲杜斯贝壳体之间充填泥晶,部分泥晶发生重结晶。围岩内部未发现其他种类的后生底栖动物碎片(图 6-A,6-B)。部分壳体中心出现焦沥青(图 6-F)。
非介壳层以颗粒泥晶灰岩和颗粒亮晶灰岩(图 6-C)为主。部分层位颗粒泥晶灰岩中的泥晶重结晶,成为亮晶方解石,形成颗粒亮晶灰岩(图 6-C)。颗粒含量为20%~45%,类型包括粒径0.5~0.8 mm的鲕粒(同心鲕、复鲕、椭形鲕)、1 mm的藻团块、内碎屑(砂屑、鲕粒灰岩)、生物碎屑(0.3~0.5 mm的枝状或肾状藻类碎片和直径0.1 mm的钙球、0.5~1 mm的腕足碎片、0.3~1 mm的介形虫、0.5~0.7 mm的腹足碎片,其中钙球、介形虫类、腹足类内部都有重结晶现象)。非介壳层中极少的生物碎屑含量表明,这一时期除哲杜斯贝外,底栖后生生物极少。
3个台地相剖面也有不同之处,大隆剖面非介壳层的颗粒泥晶灰岩受重结晶影响最弱,介壳层壳体内部粒泥灰岩残留相较打寨和龙密剖面为高,其非介壳层顶部出现枝状藻类碎屑并且局部遭受白云石化作用改造。打寨剖面非介壳层灰岩中,鲕粒灰岩和藻团块灰岩发育(图 6-C),局部层位遭受轻微白云石化作用改造。龙密剖面非介壳层粒泥灰岩受重结晶改造最为明显,顶部局部位置受白云石化作用影响最为明显,且具有较多的枝状藻类碎片。龙密剖面的S3与S4哲杜斯贝介壳层之间的非介壳灰岩为典型的微生物岩(图 6-E)。微生物岩往往在后生动物多样性极弱的环境下沉积(Riding and Liang,2005),因此,龙密剖面的微生物岩指示了这一时期较弱的后生动物活动性。
A—大隆剖面介壳层S9边缘,示簇状方解石沿壳体生长,DL-10;B—大隆剖面S10介壳层,壳体内部重结晶,簇状方解石沿壳体向内生长,壳体之间充填泥粒灰岩,DL-11;C—打寨剖面非介壳层,似球粒亮晶颗粒灰岩,Dtz-6;D—打寨剖面S3介壳层,Dtz-13;E—龙密剖面非介壳层微生物岩,LM-12;F—龙密剖面S3介壳灰岩,内部残留沥青质有机物,LM-8。所有照片均为单偏光图 6 黔桂地区法门阶台地相剖面介壳灰岩和围岩镜下沉积特征Fig.6 Microscopic sedimentary features of the Famennian brachiopod coquina and surrounding limestone from carbonate platform sections in Guizhou-Guangxi area
4.2.2 哲杜斯贝埋藏特征
大隆、打寨和龙密3个台地相剖面中的哲杜斯贝介壳层具有以下共同的埋藏特征: (1)介壳层中,80%以上的介壳为两壳铰合完整保存的壳体(图 5-C,5-D);(2)介壳层中,单瓣或破碎壳体含量不足20%,主要分布在介壳层的最底部,壳瓣破碎明显,壳瓣和壳体碎片的长轴多与层面平行,相互叠覆;(3)完整个体相互紧密接触,富集成层(图 4-C,4-D),保持原位生长的状态(图 4-D),由于壳体的排列过于密集,甚至在手标本中也很难观察到壳体与壳体之间的围岩(图 5-C,5-D);(4)在对分离出的单个保存完整的壳体进行统计时发现,有超过60%的壳体出现生长畸形,表现为壳体形态不对称、不同部位出现凹坑、某一位置壳体变薄等。结合对原位保存的哲杜斯贝的观察可以发现,这种生长畸形并未展现出一定的方向性,所以排除了埋藏后受一个方向应力的影响,更可能反映了生长发育空间狭窄、生活拥挤的特征;(5)无明显水流搬运分选特征,这在野外露头风化面上和室内观察的光面上均有验证(图 4-C,4-D;图 5-C,5-D):哲杜斯贝的完整壳体呈现密集排布,世代与世代之间呈现直接叠置的产出状态,同时,介壳层的横截面(图 4-D)上哲杜斯贝的壳体大小均一,壳长均大于0.6 cm。介壳层的横截面基本上可以代表哲杜斯贝的一个世代,同一个世代的哲杜斯贝中未出现小个体,有2种可能性, 第1种可能是绝大多数小个体没有固着就被水流冲走,第2种可能是所有小个体都成功长大。如果是前一种情况,小个体无法成功固着,被水流冲走,还是会有一定的概率使得较小的个体保留在同一世代的介壳层中,然而研究层位并未在横截面上观察到任何小个体(图 4-D)。并且,如果所有小个体都被水流搬运走,就无法形成如此密集排列的哲杜斯贝介壳层了。因此,小个体全部被水流搬运走的概率极低,更可能的原因是绝大多数的哲杜斯贝正常生长至成年期。这些特征充分说明,华南地区台地相剖面的哲杜斯贝介壳层除最底部有流水和波浪作用的影响和改造外,主体部分呈现了典型的原地埋藏保存特征。
5 讨论
5.1 哲杜斯贝原生生存环境和生活习性
虽然侯鸿飞和王士涛(1988)依据在贵州南部一些地方南丹型沉积代化组(相当于文中的五指山组)中发现的哲杜斯贝化石,认为这是生活在黔桂交界一带深水台间盆地相中的一个腕足动物群,但笔者对2种沉积相类型的5个剖面上多层哲杜斯贝介壳层的沉积学和埋藏学的研究则表明,在台间盆地相分布的哲杜斯贝属于异地搬运而来,深水型的台间盆地相并非哲杜斯贝的原生生活环境,相反,浅水碳酸盐岩台地相剖面上分布的哲杜斯贝则呈现原地埋藏的特点,表明碳酸盐岩台地相为当时哲杜斯贝生活的原生环境。打寨、大隆和龙密剖面上的厚层—巨厚层哲杜斯贝介壳灰岩系由哲杜斯贝在碳酸盐岩台地上原地生长、多个世代相互叠加而成。
对腕足动物而言,由多个个体(少则几个,多则几十至上百成千)聚集在海底形成块状或层状的密集居群现象,无论是在现代还是化石记录中都是比较普遍的。这是由于孵化出的浮游幼虫,由于没有卵黄营养,需要在很短的时间内寻找到合适的基底固着才能继续存活下来,而之前定居的个体则是良好的栖息场所,所以容易形成聚集的现象。但是像哲杜斯贝这样形成规模庞大、由单一物种组成的巨厚介壳层是非常罕见的。前人认为哲杜斯贝只能密集生活在在某些极端条件下,如富含甲烷的冷泉或海底热泉喷口等(Nolletal., 1984;Dubé,1988;Campell and Bottjer,1995;Torresetal., 2003;Peckmannetal., 2007),但从Nie等(2016)和笔者的研究结果来看,华南地区的哲杜斯贝可以生活在透光带之上的浅水碳酸盐岩台地,且没有发现与冷泉活动有关的证据,说明哲杜斯贝并非必须要生活在与冷泉活动有关的环境中,而是具有更大的生态可塑性。哲杜斯贝动物群中缺乏其他后生底栖动物,说明要么是哲杜斯贝居群具有排他性,如释放某种气体或排泄物影响其他后生底栖动物的生存,要么是其生活的环境仍然有特殊性,不适合其他后生底栖动物生存。从华南地区的情况看,前一种可能性目前尚无可靠的证据支持,后一种的可能性更大。在华南地区,哲杜斯贝是生活在不受陆源输入影响的远岸碳酸盐岩台地环境中。F/F事件(弗拉期—法门期之交生物灭绝事件)之后,在华南地区远岸的碳酸盐岩台地相中普遍发育以微生物岩为特征的巨厚碳酸盐岩沉积(融县组),缺乏除哲杜斯贝之外的其他后生底栖动物化石。与之形成鲜明对比的是在受陆源输入影响明显的近岸滨浅海区域的同期地层中,如湘中的锡矿山组和鄂西地区的写经寺组,以云南贝和小云南贝为代表的底栖动物群则十分繁盛(Tien,1938;许寿永,1984;Maetal., 2016)。因此,比较合理的解释可能是远岸的碳酸盐岩台地环境中缺乏其他后生底栖动物生长发育所需的某种营养元素,或在此生活的微生物会释放某种毒素,抑制其他后生底栖动物的生长发育。而哲杜斯贝可能在生理和生化功能上具有某种优势,可在这种环境中正常生长发育。
哲杜斯贝在华南地区生活在浅水的碳酸盐岩台地环境,而非相对深水的台间盆地相环境,可能与这2种沉积相水岩界面附近的氧逸度差异有关。Wang等(2019)对华南晚泥盆世不同沉积相的碳酸盐岩中铁锰含量(Fecarb和Mncarb)的研究表明,位于浅水台地的打寨剖面碳酸盐岩不同组分(泥晶、亮晶、腕足壳)均具有较低的铁锰含量(Fecarb为57.0~60.2 μg/g,Mncarb为7.7~12.6 μg/g),反映了由于沉积环境中的氧化还原界面远低于水岩界面之下,在水岩界面附近具有较高的氧逸度;而位于台间盆地的长塘和度里剖面的碳酸盐岩具有较高的铁锰含量(Fecarb为389.0~1180.5μg/g,Mncarb为41.2~188.0μg/g),反映了沉积环境中的氧化还原界面非常接近水岩界面,受还原性孔隙水的影响,在水岩界面附近具有较低的氧逸度。因此,笔者认为台间盆地相水岩界面附近较低的氧逸度可能不利于哲杜斯贝浮游幼虫在海底固着生存,因而不能形成原生的居群。
5.2 居群生态学
大隆、打寨和龙密剖面上的哲杜斯贝显示了典型的原地埋藏特征,满足居群生态学研究的前提。因此,通过野外剖面和室内光面的定性观察,并定量分析壳长和壳体对称性等形态特征指标(图 7),对这3个哲杜斯贝居群的生态学特征进行探讨并横向比较。
大隆、打寨和龙密剖面上的3个哲杜斯贝居群最显著的共性之一是: 介壳层与非介壳层分界截然,介壳层中的壳体堆叠密集,厚度大、呈层状产出,丰富度极高(图 4-C,4-D)。相当数量的壳体呈现出由于密集生长导致生存空间有限、壳体生长畸形(malformation)的现象,壳体的生长发育不对称。如前文台地相哲杜斯贝埋藏特点部分中所描述的,壳体畸形现象在大隆、打寨和龙密剖面的哲杜斯贝中极为普遍,畸形壳体占总壳体的比例分别高达57%、67%和84%(n=294,n=521,n=438)。类似的生长畸形现象亦常见于其他地区发现的哲杜斯贝动物群中(Biernat,1967;Cloud and Boucot,1971;Nolletal., 1984;Baliński and Biernat,2003),表明在合适的环境条件下,哲杜斯贝倾向于形成个体密集的生活群落。
大隆、打寨和龙密剖面上3个哲杜斯贝居群最显著的共性之二是: 哲杜斯贝所生活的群落物种多样性极低。一方面,在哲杜斯贝介壳层中,群落由单一属构成(monotonous community),未观察到其他种类的底栖后生生物与之共存。另一方面,在非介壳层中,生物化石极少,只存在约1%的腕足类、介形类、腹足类和藻类等碎片,生态系统极其单调(图 6)。介壳层和非介壳层的特点均表明,这一地区与哲杜斯贝同时的环境并不适合其他底栖后生生物的生存。在全世界其他地区发现的哲杜斯贝也具有类似的高丰度、低分异度的种群特征(Biernat,1967;Cloud and Boucot,1971;Ageretal., 1976;Nolletal., 1984;Baliński and Biernat,2003;Nieetal., 2016)。关注哲杜斯贝的居群结构发现,3个剖面中哲杜斯贝动物群样本的壳长分别介于11~37 mm、5~37 mm和7~27 mm,壳长均值分别为20.0 mm、14.0 mm和16.9 mm。3个居群的壳体大小—频率分布图均呈现左偏态(left skewed distribution)(图 7-A1,7-B1,7-C1)。为避免在名称上将小个体与营漂浮生活的、未固着的幼虫相混淆,具体在本研究中,下文中提到的“幼年期”哲杜斯贝将表示已经有硬体的小个体哲杜斯贝。左偏特征的大小—频率分布表明,3个居群中的个体均以成年期或接近成年的个体为主,壳长小于10 mm的壳体鲜有出现。同时,生存曲线呈现典型的上凸特征,表明3个哲杜斯贝居群具有极低的幼年期死亡率。其中,大隆居群的哲杜斯贝的生存曲线在壳长为12.8 mm时才开始出现微弱下降,打寨居群的哲杜斯贝生存曲线在壳长为10.2 mm时出现较为显著的下降,龙密居群的哲杜斯贝生存曲线在壳长达到13.4 mm时出现下降。10 mm的壳长可以作为华南哲杜斯贝幼年期和成年期的分界。生存曲线指示哲杜斯贝到成年期才开始有较高的死亡率,反映了绝大多数的个体都可以生活至成年阶段。然而,对比已有的腕足大小—频率分布和生存曲线(Richards and Bambach,1975;Bitner,2001;Tomašovch,2004,2006;Zuschinetal., 2004,2005;Websteretal., 2008;Panetal., 2012;Skawina,2013;Lü and Ma,2017;Huangetal., 2018;Yuanetal., 2019)发现,包括腕足在内的、正常生活在在硬质基底上的底栖动物居群通常具有较高的幼年期死亡率和较低的成年期死亡率,即通常有相当一部分数量的腕足动物在幼年期夭折。因此,哲杜斯贝与常见的生活在硬质基底上腕足动物群的居群动力学特征有显著不同。
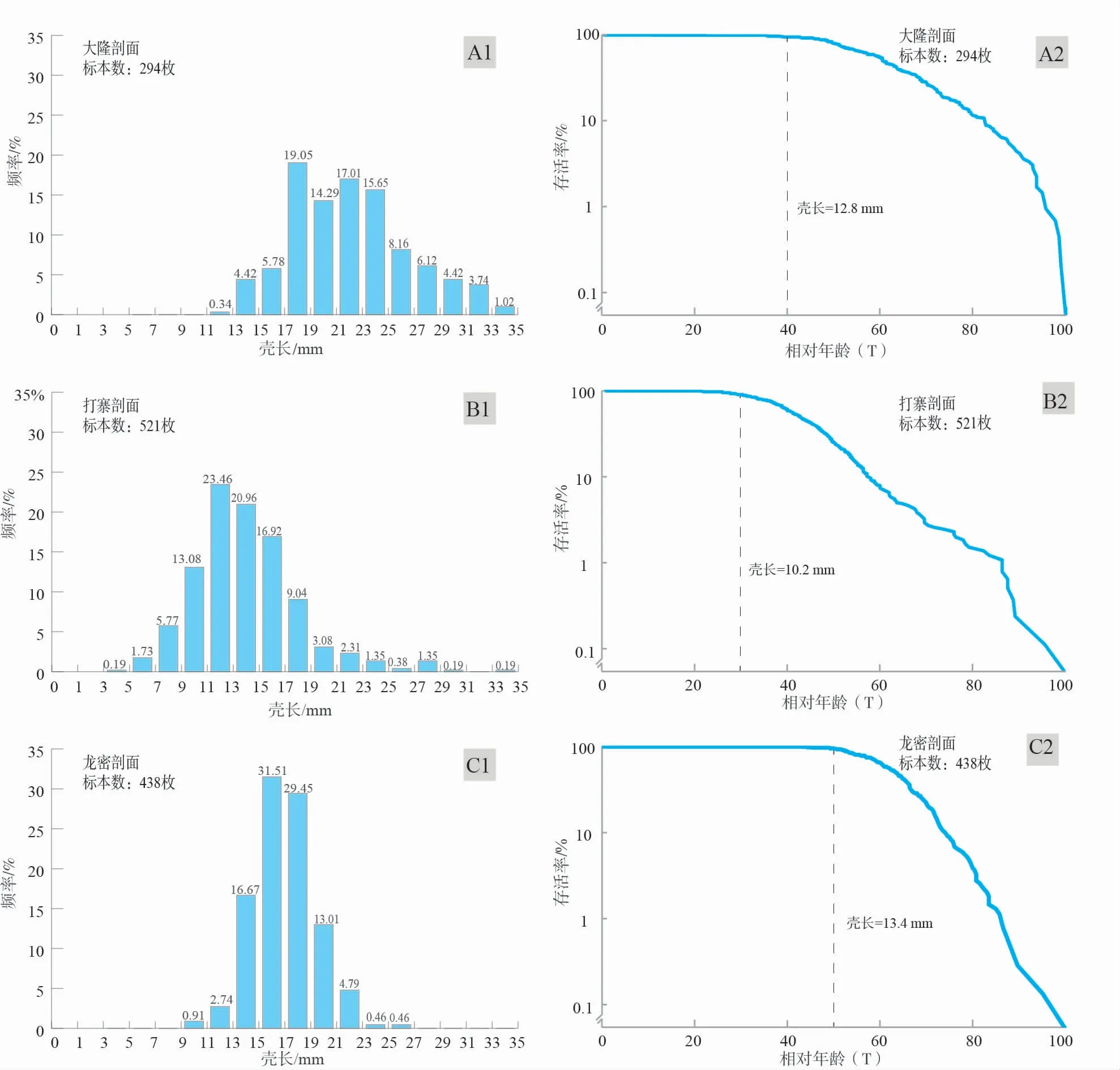
图 7 黔桂地区台地相剖面哲杜斯贝壳体大小频率分布(A1,B1,C1)和种群生存曲线(A2,B2,C2)Fig.7 Size-frequency histograms(A1,B1,C1)and survivorship curves(A2,B2,C2)of the Dzieduszyckia from carbonate platform sections in Guizhou-Guangxi area
对大隆、打寨和龙密3个剖面上发育的厚层介壳灰岩断面观察结果也支持壳体大小频率分布和生存曲线所反映的居群动力学特征。在多个世代连续生长的厚层至巨厚介壳层(如打寨的第1、2、3介壳层;大隆的第7、8、12、15介壳层;龙密的第3、4介壳层)中,无论是同一世代(同一水平面分布),还是多个世代(不同水平面分布),哲杜斯贝个体的大小均无明显变化,多数接近或超过壳体长度的均值(图 4-D),表明同一世代的个体多数可生活至成年,除非是靠近群落边缘生活的个体,或受环境变化影响明显的世代,个体大小才会普遍低于均值。
结合哲杜斯贝高丰度、低物种分异度的居群特征,以及异于普通腕足动物生存曲线的特征,推测哲杜斯贝异于普通腕足而特有的密集生长的生活方式可能有利于克服外界环境的压力。哲杜斯贝的密集生长,很有可能区域性地改变局部生活环境,比如有可能有利于哲杜斯贝的滤食、抗击较强的水动力环境等。这种密集生长的居群生活方式,对于提高小个体存活率可能起到了至关重要的作用。
6 结论
1)对华南地区2种沉积相区5个剖面哲杜斯贝介壳层的沉积学和埋藏学特征的比较研究证实,华南地区法门阶中的哲杜斯贝动物群原生生活环境是远岸的浅水碳酸盐岩台地环境,而非前人所认为的较深水台间盆地环境。哲杜斯贝在远岸浅水碳酸盐岩台地上形成密集的居群,偏好氧逸度较高的环境,与北美和摩洛哥等地的哲杜斯贝生活在有冷泉信号的环境不同,表明哲杜斯贝具有较强的生态可塑性。
2)从华南和世界其他地方已经发现的化石记录看,哲杜斯贝总是以群居的方式产出,形成以其占优势的属种单调的动物群,表明喜欢群居是哲杜斯贝的一个重要生活习性。在华南地区,哲杜斯贝生活在受陆源输入影响微弱、微生物岩发育的沉积环境中,缺乏其他后生底栖生物;而以云南贝和小云南贝等为代表的同期其他后生底栖生物则在受陆源输入影响明显的近岸滨浅海区域十分繁盛,表明哲杜斯贝动物群生活环境中缺乏其他后生底栖生物的存在,这不是哲杜斯贝排斥其他后生底栖生物的结果,而是其生活的环境不适合其他后生底栖生物的生存。推测在F/F事件之后,华南地区远岸碳酸盐岩台地环境可能较为特殊,抑制其他后生底栖动物的生长发育,而哲杜斯贝可能在生理和生化功能上具有某种优势,可在这种环境中正常生长发育。
3)壳体大小—频率分布和生存曲线所揭示的华南地区哲杜斯贝原生居群种群动力学特征表明,哲杜斯贝小个体存活率高,成年期死亡率上升,与一般腕足下凹的生存曲线有很大差异。独特的密集群居生活方式可能有利于哲杜斯贝更好地克服外界环境的压力、改变局部生境条件,从而提高个体存活率。
