自生泥晶:过去,现在和未来
2022-06-04沈越峰张乐沈臻欢陈亚婕
沈越峰,张乐,沈臻欢,陈亚婕
1.合肥工业大学资源与环境工程学院,合肥 230009
2.中国地质大学(北京)地球科学与资源学院,北京 100083
0 引言
自生泥晶(autochthonous micrite,简称automicrite),也可被称为原地微晶碳酸钙(in situmicrocrystalline Ca-carbonate)或原地灰泥(in place lime mud),以区分他生泥晶或异地泥晶(allochthonous micrite,简称allomicrite)。自Wolf[1]提出这一概念以以来,国内外众多学者对不同碳酸盐岩环境中自生泥晶的显微特征[2]和地球化学特征[3]、自生泥晶形成过程和来源[4]等问题进行了深入的研究和探讨。伴随着微生物岩[5-15]、碳酸盐灰泥丘[16-17]和生物矿化作用及其衍生的有机矿化作用[18-20]研究的进一步深入,碳酸盐自生泥晶的研究也取得了重要进展。
然而,在自生泥晶的研究过程中,也存在着部分争议。首先,“微生物岩”这一术语[21]已被广泛用于标记各种微晶礁(丘)岩,包括大多数自生泥晶结构。而且,一部分作者将微生物岩等同于自生泥晶[2]。然而,Bourque[22]认为如果没有令人信服的证据证明是主要由底栖微生物群落构建的岩相,微生物岩这个术语就变得无意义。再者,很多生物纹层结构或凝块结构不是微生物岩所特有的,海绵、藻类和其他后生动物也可以形成类似的结构。因此将微生物岩简单地等同于自生泥晶是有争议的。其次,微生物岩(主要是自生泥晶)被广泛用来追踪其成因和来源并指示同时期沉积水体的地球化学特征[3,23],但微生物岩保留地球化学组成的机制和原理尚未清晰,有待于进一步的总结和实践。最后,区别自生泥晶和他生泥晶对碳酸盐岩分类,特别是对碳酸盐岩建造的分类具有显著意义,并且能够更合理地解释地质历史时期碳酸盐岩建造的生物群落组成及其演化[4]。
为此,本文在前人的基础上,对自生泥晶概念的提出和发展进行了概括;对自生泥晶的矿物组成、显微结构、荧光和阴极发光显微特征进行了总结;对自生泥晶的形成过程和来源(包括生物矿化作用、有机矿化作用和无机沉淀三种方式)进行了探讨;对自生泥晶在不同沉积环境碳酸盐岩中的贡献及其地球化学指示进行了论述;对自生泥晶在地质历史时期的分布做了归纳;最后对自生泥晶的后续研究方向提出了一些展望。
1 自生泥晶概念的提出和发展
泥晶(micrite)这一术语在碳酸盐岩研究中已经得到了广泛的应用。它起初是微晶方解石“microcrystalline calcite”的缩写,指的是隐晶至微晶级别的碳酸盐颗粒。泥晶这一概念是由Folk[24]首先提出的,起初是一个指示成因的术语,指的是固结的因机械沉积而形成的灰泥。现在绝大部分学者将其作为一个非成因指示的术语,指的是粒度通常小于4 μm(可能有不同的标准)的细粒碳酸盐基质[25-26]。自生泥晶(autochthonous micrite,简称automicrite)的概念首先是由Wolf[1]在阐述钙藻(钙质微生物)的沉积产物时提出的,指的是原地形成的碳酸钙泥晶基质(matrices formedin situ/in place),用以区分非原地形成的泥晶,即他生泥晶或异地泥晶。Wolf[1]最初介绍的是与钙藻(微生物)相关的自生泥晶(即藻自生泥晶,algal automicrite)。Flügel[27]总结指出,自生泥晶是指在海底或沉积物中由于物理化学、微生物、光合作用和/或生物化学过程而在原地形成的细粒(隐晶—微晶质)自生碳酸钙矿物(方解石和/或文石)。Jameset al.[28]在Origin of Carbonate Sedimentary Rocks一书中对自生泥晶的概念进行了进一步的提炼,指出自生泥晶是指通过无机和/或有机媒介过程作用而沉淀在海底或者沉积物中的文石或方解石胶结物。这一微晶碳酸盐明显不是沉积物,而是同沉积胶结物。自生泥晶具有特定的结构和较为单一的矿物化学组成。作为原地矿化作用的初级产物,自生泥晶与他生泥晶具有不同的成因。他生泥晶指的是非原地形成的泥晶,来源于其他各种碳酸盐颗粒的破碎作用和原始细粒碳酸盐物质的沉积作用而形成的微泥晶基质,其可能包括不同的矿物和化学组成。
自生泥晶的概念在微生物岩[2-3,29-35]和碳酸盐岩礁丘尤其是灰泥丘[4,36-56]研究过程中得到了进一步深化。近几年,自生泥晶的概念得到了进一步扩大,应用到了与甲烷有关的冷泉碳酸盐岩[57-59]和陆相碳酸盐沉积(泉水沉积,泉华,钙华,石灰华,钙质结壳等[28,60-64])当中。
很多研究者将自生泥晶(automicrite)这一术语等 同 于 微 生 物 岩[2,21,65](microbialite)或 有 机 泥晶[29,31,42,66-69](organomicrite)。根据形成模式和形成位置的不同,Reitner[29]又将有机泥晶分成两类,包括堆积有机泥晶和袋状有机泥晶。堆积有机泥晶指的是呈现出凝块石状、叠层石状或呈块状硬底类型的垂直生长构造。袋状有机泥晶是由半封闭微孔洞中的后生动物(大部分是海绵)封闭的降解或腐烂组织中,或者是由钻孔动物钻孔形成的孔洞中发育充填构造而形成的。此外,Leeset al.[70]在灰泥丘的研究中提出了多泥晶结构(polymud fabric)的特征,这一概念在自生泥晶的研究中也被许多作者所沿用[47,56,71]。Keimet al.[72]在研究意大利三叠纪的白云岩陡斜坡沉积中提出了自生泥晶相(automicrite facies,指代具有大量自生泥晶的野外地层)和自生泥晶工厂(automicrite factory)的概念。Pomaret al.[73]重新定义了Schlager[74-75]的底栖“灰泥丘工厂(mud mound factory)”,重命名为“底栖自生泥晶工厂(benthic automicrite factory)”。一些作者又将微生物工厂(microbial factory)等同于底栖自生泥晶工厂[76-77]。
国内对于自生泥晶的研究相对较少,《沉积地球化学应用》讲座编写组[78]首次提到了自生泥晶一词,并简单介绍了不同来源泥晶的地球化学组成特征。朱士兴[79]总结了第30届国际地质大会题为“钙藻、叠层石和灰泥丘的演化和环境意义”的2~6学科讨论会材料,在国内首次介绍了automicrite的成因、分类、形成环境等领域的国际研究现状和发展趋势。朱士兴[79]将自生泥晶当作一种岩石类型,翻译成“原地微晶灰岩”。进入21世纪,随着国内对碳酸盐岩研究的进一步深入,自生泥晶的概念已被广泛地应用到了碳酸盐岩研究当中。马永生等[80-81]在《碳酸盐岩微相:分析、解释及应用》中也将自生泥晶翻译成了“自生泥晶灰岩”。贾志鑫[82]通过对湖北宜昌九龙湾剖面震旦陡山沱组盖帽碳酸盐岩微相研究,推测本层盖帽碳酸盐岩原本是颗粒和基质均以泥晶形式沉淀的自生泥晶灰岩。梅冥相等[8,83-85]在研究叠层石生物丘(层)和微生物诱导(有机矿化作用)的沉积构造时,说明了这些沉积构造都是底栖自生泥晶工厂[73]的产物,并且强调了自生(原地)泥晶成因的不确定性。杨勇强[86]提到了湖相自生泥晶的碳氧同位素特征。杜翔[87]和杨巍[88]在四川盆地东部晚二叠世海洋底栖生物礁系统研究中,识别出三种台地边缘礁微相类型,强调了自生泥晶在微生物丘格架形成中的重要性。闫振等[16]在介绍灰泥丘时,提到了原地成因的灰泥及其形成方式,并指出灰泥丘的形成机制、时空演化、控制因素及古地理古环境指示意义是将来灰泥丘研究的主要方向。沈越峰等[89]探讨了塔里木盆地奥陶系瓶筐石—寻常海绵碳酸盐岩丘自生泥晶的来源。陈百兵[90]在豫西宜阳地区寒武系馒头组鲕粒灰岩特征及演化的研究中指出了微生物自生泥晶化对鲕粒中泥晶形成的作用[91]。王龙[92]研究了太行山中南部及其邻区寒武系微生物碳酸盐岩主导的生物丘,为以原地自生泥晶构成为主的生物丘提供了较好实例和岩石记录。
本文综合前人论述,按照Wolf[1]以及Reitneret al.[31]的严格定义,并进行了部分扩大,将automicrite这一英文术语理解为自生泥晶,即通过无机和/或有机媒介过程作用在原地形成的泥晶碳酸钙。自生泥晶是碳酸盐岩中一个重要的不具成因指示的结构组分,微生物岩和有机泥晶都应归入自生泥晶的范畴[22,38,75]。
2 自生泥晶的矿物组成和结构特征
自生泥晶有一定的矿物组成,粒度结构、超微结构和外貌形态,并且通常表现为不同的多泥晶结构(polymud fabrics)特征。
2.1 自生泥晶的矿物组成
现代海洋环境中产出的自生泥晶主要是以文石为主,地质历史时期海洋中产出的自生泥晶的矿物组成有可能是文石,亦有可能是方解石,可能与同时期海水地球化学组成(钙离子饱和度,镁钙比等)相关[2,29,58,93-98]。
2.2 自生泥晶的粒度
自生泥晶的粒度根据一般泥晶的概念,是隐晶(在普通偏光显微镜下不可识别)或微晶(在普通偏光显微镜下能识别粒度,但在双目放大镜或手标本中不能识别的)级别的。Folk[24]严格定义了泥晶的粒度,指的是只能在显微镜下或借用扫描电镜才能识别的碳酸盐颗粒,晶体粒度小于4 μm,平均在2~3 μm。然而在碳酸盐岩岩石学分析中,尤其是碳酸盐岩微相分析中,不同泥晶晶体粒度的上限也被应用,包括:10 μm[99]、20 μm[100]和30 μm[25]。 Bosellini[101]还区分了一类泥晶micrite I(晶体粒径<4 μm)和二类泥晶micrite II(晶体粒径为4~30 μm)。Folk[102]提出了微泥晶(minimicrite)的概念,这些微泥晶的颗粒大部分小于1 μm。Bernier[103]还强调了区分非常细粒泥晶(粒径通常小于2 μm)和其他泥晶在岩相学分析中的重要性。由上,本文限定自生泥晶的粒度大小为小于4 μm。
2.3 自生泥晶的超微结构和外貌形态
自生泥晶在扫描电镜(SEM)或冷冻扫描电镜(CryoSEM)下的外貌形态主要呈现出均一的微米或纳米级别的光滑菱面体状、哑铃状、棒状、针状、晶球状、卵圆状或者纳米球状特征[2,29,31,41,55,104-105]。实验研究表明自生泥晶的形状可能带有聚合物的特点。尽管它们形成一定形状的粒状沉积物,但它也可以以球粒状(peloidal)、凝块状(clotted)或者纹层状包覆(laminated coating)骨骼或贴在生物礁丘中的孔隙或洞壁上的形式出现,呈现出抗重力作用的特征。大多数自生泥晶以不同结构类型的多泥晶(polymud fabric[4])组构出现,可以形成生物层状结构、凝块球粒微结构或极细的隐晶质结构[72]。
3 自生泥晶的荧光和阴极发光显微特征
自生泥晶具有特殊的荧光和阴极发光显微特征,是其区别于他生泥晶的一个良好标志,将它们的这两个特征归纳如下。
3.1 荧光显微特征
碳酸盐岩中的荧光性大多可能是由其中所含的有机质残留导致的[67,106-108],有些矿物种类的荧光性可能是由稀土元素激化引起的[106]。碳酸盐矿物中的荧光特性通常可以用来识别隐藏的显微结构[107],研究纹层状灰岩(比如说蓝细菌、藻类、花粉和孢子等)的有机质组成,以及理解生物矿化作用过程。Neuweileret al.[30]将显微荧光技术用于区分自生泥晶和他生泥晶,并取得了较好的效果,指出自生泥晶的荧光显微特征要比他生泥晶的强烈和明亮。随后的研究也证实了其观点,认为由于自生泥晶中大量有机质的赋存,导致其荧光显微特征有别于他生泥晶、骨骼组分和胶结物等[4,38-41,46,109-121](图1a,b)。

图1 自生泥晶的普通偏光、荧光和阴极发光显微特征(a,c)普通偏光显微照片,单偏光;(b)荧光显微照片;(d)阴极发光显微照片。Si=硅质交代;Cc=方解石胶结物;AM1=带有海绵骨针的球粒自生泥晶;AM4=隐晶质自生泥晶;薄片来自于塔里木盆地巴楚地区勒牙伊里塔格露头奥陶系一间房组瓶筐石—海绵礁丘Fig.1 Microphotographs of automicrites in thin sections from Ordovician calathid ̄sponge carbonate mounds in the Leyayilitag outcrop, Yijianfang Formation, Bachu, Tarim Basin, Xinjiang(a,c)under normal light;(b)fluorescent light;(d)cathodoluminescent light.Si=replacive silica;Cc=calcite cement;AM1=automicrite fabric with sponge spicules;AM4=aphanitic automicrite
3.2 阴极发光显微特征
阴极发光显微镜被广泛运用于碳酸盐岩研究中[122-124],尤其是在分析和解释碳酸盐岩成岩作用阶段[125-126],恢复和识别碳酸盐岩原始颗粒、结构和构造[127-129],利用碳酸盐岩胶结物环带研究解释碳酸盐沉积物的埋藏史、成岩孔隙水特征[129-137],以及白云石化作用[138-140]等方面具有非常显著的意义。碳酸盐矿物的阴极发光性质主要受控于锰、某些稀土元素以及铁的相对丰度。Mn2+和三价稀土离子是产生阴极发光最重要的激发剂,而Fe2+则是最主要的淬灭剂[123,129,135]。
虽然前人对自生泥晶的荧光显微特征研究较多,但对它们的阴极发光显微特征研究较少。大部分的研究主要集中于恢复和识别含有自生泥晶的碳酸盐岩非骨骼颗粒(即鲕粒,球粒等[123,128,141-145])。Immenhauseret al.[146]提到基质泥晶并不发光,而重结晶的泥晶会有暗红色发光。Coimbraet al.[147-148]进一步提到了基质泥晶(他生泥晶)呈现出暗色的阴极发光特征,但并没有提及自生泥晶的阴极发光特征,也没有对自生泥晶和他生泥晶进行很好的区别。Floquetet al.[47]首先展示了自生泥晶阴极发光的显微照片,其中M1 呈现出亮红—橙色的阴极发光特征,但对自生泥晶的阴极发光性没有进行详细的描述和解释。Braithwaite[149]讨论了碳酸钙荧光和阴极发光显微特征之间可能存在的联系是通过以有机质为媒介将稀土元素摄取到晶格当中。Shenet al.[4]发现自生泥晶的荧光和阴极发光显微特征具有很好的对应性,进一步指出自生泥晶是通过有机矿化作用形成的(图1c,d)。而且,自生泥晶中的锰含量也相对较高,因此认为相对较高的锰和铈以及其他稀土元素含量可能是自生泥晶产生阴极发光的主要原因[40,45,122,145,150]。自生泥晶的阴极发光性状和其中包含的主微量—稀土元素之间的关系还有待于更进一步的研究。
4 自生泥晶的形成过程和来源
根据自生泥晶的形成方式和过程,可以把自生泥晶的来源分为三类:无机成因,生物控制和诱导矿化作用,以及生物影响矿化作用,也就是物理化学无机沉淀作用、生物矿化作用和有机矿化作用[2,27-28]。
4.1 无机成因
无机成因(非生物)自生泥晶指的是由于盐度和水体温度变化导致的,通过物理化学作用在原地沉淀的微晶碳酸钙胶结物,这一过程没有生物或者有机质的参与。无机成因自生泥晶主要可能包括与甲烷(渗漏)相关的(冷泉)自生碳酸盐(岩)(methanerelated authigenic carbonates[58-59,151-152])以及陆相碳酸盐(岩)(如钙质土壤,泉华,钙华,洞穴沉积物,湖相碳酸盐岩等[2,60,64,86,118,153-156])。但是这些自生泥晶的无机成因较不确定,可能与微生物控制或诱导作用有关[64,118,156],相关基础研究有待于进一步开展。
4.2 生物矿化作用
生物矿化作用[157]形成的自生泥晶是指由生物体通过生物大分子的调控生成自生泥晶的过程,分为两种形式,生物控制的矿化过程而形成的自生泥晶和生物诱导的矿化过程而形成的自生泥晶[2,27-28]。
生物控制矿化作用形成的自生泥晶是指生物(光养蓝细菌和藻类)在不受外界环境影响的条件下,通过生命活动和新陈代谢过程,将二氧化碳还原为有机碳化合物,从而将溶解度平衡向碳酸盐沉淀转移,促使自生泥晶的沉淀[2,27-28]。蓝细菌产生的自生泥晶在现代潮间带和高盐度湖相环境中被观察到[158-162]。蓝细菌泥晶通常发育为微球粒晶体大小<2μm。丝状蓝细菌在泥晶形成中的重要性通过底栖蓝细菌的钙化作用和保存作用潜力得到了总结[163]。在丝状蓝细菌死亡和有机质降解之后,在底栖丝状蓝细菌的胞外聚合物(EPS)有机层中的钙离子被释放出来,可能是导致碳酸盐灰泥形成的原因。Kaźmierczaket al.[164-165]和Kempeet al.[166]认为许多侏罗纪泥晶和球粒灰岩都是底栖球状蓝细菌席不同程度的密集钙化的产物,并且从古代生物礁中描述的许多微晶胶结物可能是体内钙化蓝细菌的产物。底栖球状蓝细菌席的原地钙化作用,比如在土耳其的高碱性苏打湖Van Lake,可能代表了蓝细菌控制泥晶或球粒泥晶形成的一种模式[165 ̄166]。球状蓝细菌席通过体内以及在围绕着细胞的黏液层上或黏液层内的泥晶文石的沉淀而进行矿化作用。受碳酸钙饱和度控制球状蓝细菌席钙化强度的变化产生了均匀的泥晶和似球粒体,产生了球粒泥晶结构。在高盐海相环境中形成的蓝细菌席中,常出现类似于高盐湖Van Lake 的泥晶球粒颗粒结构(Baffin Bay,Texas[158];Laguna Mormona,California[159];Shark Bay,Australia[160];Gulf of Aqaba[160-161])。
生物诱导矿化作用形成的自生泥晶是指生物(异养和化养细菌及其他微生物)的生命活动和新陈代谢过程引起细菌周围微环境的变化,并诱导微晶碳酸钙沉淀[2,27-28]。有机质的细菌降解可能是沉积物早期成岩作用的主要能量来源。众多研究者已经证明并提出,细菌的生命过程和细菌对有机物的分解作用会导致其周围微环境的物理化学变化,从而促使微晶碳酸钙的沉淀,这些现象不仅得到了实验室的证实,在现代的各种碳酸盐岩环境中也能够被观察到,包括土壤、淡水和海洋环境,尤其是潟湖环境中[167]。
4.3 有机矿化作用
有机矿化作用[168]形成的自生泥晶是指以活体生物产生的有机质(Organic matter,OM;螯合钙离子的有机大分子)为载体,而又独立于活体生物生命活动和新陈代谢作用形成的自生泥晶(图2),也叫有机泥晶[29,31]。这些有机化合物可能是活体生物的排泄产物或者是它们的分离部分,又或者是死亡的生物残留物或者副产品,被释放到水体或者被纳入到土壤,沉积物或岩石中[31,120]。
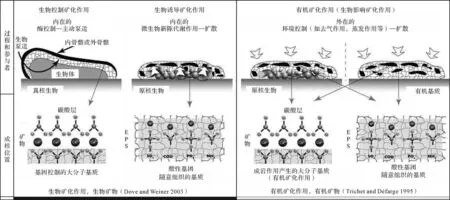
图2 生物矿化作用和有机矿化作用形成自生泥晶的模式(修改自Dupraz et al.[2])注意:本文有机矿化作用的定义沿用Trichet et al.[168]的狭义有机矿化作用定义Fig.2 Models of biomineralization and organomineralization in automicrite production (modified from Dupraz et al.[2])Note:The term‘organomineralization’in this study follows the definition by Trichet et al.[168]
基于与酸性有机大分子(AOM)的相互作用的碳酸盐矿化作用概念最初用于解释生物的骨架结构[157,169-171],并在自生泥晶形成的背景下进一步发展。Reitner[29]在大堡礁蜥蜴岛附近礁洞里结壳微生物岩中研究了与周围海水同位素平衡条件下沉淀并受特定有机物质组成控制的现代镁方解石自生泥晶的形成。这些微生物岩是在黑暗环境中形成的,光养生物是不存在的。控制有机泥晶形成的最重要因素是存在大量具有固定和结合二价阳离子(主要是Ca2+和Mg2+)能力的特定有机大分子。形成钙晶体所需碱度的增加是由氨化作用(生物比如海绵的微生物腐烂降解)、硫酸盐还原作用(在分层水体的厌氧层中)或通过碳酸盐和硅酸盐风化作用[31]。Neuweileret al.[39]讨论了溶解的腐殖质对原地微晶碳酸钙沉淀的控制作用。
有机泥晶的形成受有机大分子的控制。有机物的特点是有大量的特定化合物,这些化合物来自微生物、游离有机物和腐烂的生物体。利用色谱和荧光显微镜在现代和古代的自生泥晶中发现了酸性有机大分子(AOM)螯合钙离子的证据[31,67]。荧光显微镜在测定残留有机物中具有特别的作用[107-108]。酸性有机大分子有两种来源:1)真核生物(海绵,珊瑚,苔藓虫,有孔虫,藻类等[39])和原核生物(细菌[2])通过代谢酶和腐烂过程产生AOM;2)外部来源的AOM,作为开放水体中溶解有机物的一部分。这些大分子在未成熟的微生物岩中或在水/成熟沉积物间隙中富集。它们由生物体(如生物膜)产生和/或固定,或被圈闭在微洞穴和囊袋中。这些化合物组成的差异可能导致形成不同的碳酸盐岩组构:酸性氨基酸(Asp+Glu)的富集可能与叠层石和凝块岩组构的形成有关;有限空间(如微洞穴)内的蛋白质物质可能与球粒囊袋,球粒包壳和球粒叠层石的形成相关;以及富含细菌的软海绵组织的降解,导致微球粒结构的形成,恰好反映了之前海绵体的某些部分。许多微生物岩(叠层石、凝块石、树枝石等)、非骨骼颗粒(球粒、鲕粒、核形石及包壳颗粒等)和碳酸盐岩礁丘的形成都与有机矿化作用形成的自生泥晶有关。
综上,自生泥晶的形成方式多种多样,导致自生泥晶也有不同的来源。在碳酸盐岩微相分析中,区分不同类型的自生泥晶,对探讨自生泥晶的特征及其来源,恢复碳酸盐岩沉积古环境具有重要的指示意义[172]。因此,不能简单将自生泥晶归类为微生物成因的,需要综合自生泥晶不同的特点来分析其成因及形成方式。
5 自生泥晶的地球化学指示
泥晶(灰岩)以组分较为单一、低孔低渗透性为特征,故而许多学者认为泥晶(灰岩)受到的成岩作用影响较弱,可能较好地反映古海水地球化学组成,因而能够较好地评估古环境条件[173-175]。然而,近期研究表明,不同成因、不同环境的泥晶所反映的古海水地球化学特征具有较大的不同[55,146,176-178]。多数学者认为自生泥晶相对于他生泥晶,其地球化学特征能够较好地反映古海水地球化学组成[23,55,146,179-180]。但是,也有学者认为不同成因,不同环境中的自生泥晶,其地球化学特特亦有所不同[45,146]。下文对自生泥晶的地球化学指示意义进行了归纳和总结。
5.1 元素地球化学
自生泥晶在元素地球化学方面的应用主要包括两个方面:一方面,大多数自生泥晶包含了其形成时期沉淀流体的元素地球化学特征(主、微量元素,稀土元素等),因而元素地球化学分析可以用于识别自生泥晶形成的(微)环境信息(沉淀水体);另一方面,自生泥晶的形成过程使多种金属元素富集,这个富集的过程可能是选择性的也可能是非选择性的,而这些元素的富集保存可能会提供不同的生物(有机)标志,进而有助于确定和区别自生泥晶的形成过程和来源[3]。近年来,对自生泥晶元素地球化学的研究主要集中在识别现代和地质历史时期微生物岩、钙质微生物岩、鲕粒和海绵礁丘等含自生泥晶组构的(微)环境信息方面[3,23,40,44-45,55,110,119,156,181-206]。
首先,自生泥晶可能有利于识别同时期海水元素地球化学特征,因而能够反映古海水的氧化还原条件及其演化。自Webb et al.[23]测定大堡礁Heron Reef浅水礁格架洞穴中全新世微生物岩稀土和钇元素(REE+Y),发现微生物岩的REE+Y 地球化学特征与周围充分氧化的浅海海水地球化学特征相一致以来,尤其是伴随新的原位微区元素分析技术的发展(激光剥蚀—电感耦合等离子体质谱法,LA-ICP-MS)微生物来源的自生泥晶(即微生物岩,主要是叠层石、凝块石、微生物席、生物膜或钙质微生物)被认为是反映同时期海水稀土元素地球化学组成(REE+Y)的最佳指示剂(见Webb et al.[3]总结)。随后,许多作者对不同地质历史时期微生物岩的稀土元素进行了测量和分析,例如:Nothdurft et al.[182]测定了澳大利亚西部坎宁盆地Lennard Shelf 地区晚泥盆世钙质微生物岩中的钙质微生物Renalcis 样品;Olivier et al.[184]测定了晚侏罗世珊瑚—海绵—微生物礁中的微生物岩样品;Allwood et al.[188]测定了来自澳大利亚西部Pilbara Craton 的具有34.5 亿年历史的太古宙Strelley Pool Formation 中叠层石样品;Matyszkiewicz et al.[192]测定了波兰南部Kraków ̄Częstochowa Upland 南部晚侏罗世Oxfordian 碳酸盐岩建造中微生物岩样品;Kamber et al.[194]分析、整理和总结了几个不同构造区和不同地质年龄的太古宙叠层石样品;Della Porta et al.[197]测定了摩洛哥High Atlas 早侏罗世Sinemurian 硅质海绵微生物丘中的微生物岩;van Smeerdijk Hood et al.[198]测定了澳大利亚南部新元古代晚成冰期碳酸盐岩礁中的微生物岩;Tang et al.[199]测定了华北中元古代微生物岩;Kalvoda et al.[205]测定了捷克晚泥盆世Hangenberg时期的薄层纹层状微生物岩;Hüeter et al.[55]测定了克罗地亚特提斯中部白垩纪Aptian 时期大洋缺氧事件OAE1a 的浅水碳酸盐自生泥晶。此外,针对二叠三叠边界的微生物岩,Loope et al.[193],Collin et al.[196],Eltom et al.[201]也进行了相关的稀土元素分析。以上所有研究基本上认为微生物岩能够很好地反映同时期海水稀土元素地球化学特征。此外,Li et al.[202-203]利用原位LA-ICP-MS 对现代和下三叠统鲕粒中的自生泥晶组构进行了稀土元素地球化学分析,进一步证实了鲕粒中的自生泥晶组构能够很好指示特定时期海水稀土元素组成。
然而,对于动物来源(海绵的有机矿化作用而形成)的自生泥晶,它们蕴含的稀土元素地球化学特征可能完全保留、或部分保留、或不保留同时期海水稀土元素地球化学特征。Neuweiler et al.[40]分析了下白垩统深水碳酸盐岩丘中的自生泥晶的稀土元素,发现自生泥晶具有明显的Ce 正异常特征,而海水胶结物是明显的Ce 负异常。Desrochers et al.[45]分析了加拿大Anticosti Island 下志留统苔藓虫—海绵碳酸盐岩建造中的自生泥晶,也发现了同样的问题。而Neuweiler et al.[44]对瑞士南部下侏罗统Sinemurian 晚期的平底晶洞灰岩中的自生泥晶的稀土元素进行了测定,发现自生泥晶与海水胶结物同时表现出Ce 负异常,但他们对出现的Ce负异常做出了一定的解释。Larmagnat et al.[50]测定了加拿大魁北克地区Trenton Group 的奥陶纪海绵碳酸盐岩丘中的自生泥晶的稀土元素,也发现自生泥晶的稀土元素特征与海水胶结物的稀土元素特征是一致的。
在痕量或微量过渡金属元素或生命元素方面,Kamber et al.[185]发现了钙质微生物Renalcis 中V,Sn,Cu和Zn等生命元素的大量富集。Collin et al.[196]分析了二叠纪末期生物大灭绝事件后微生物岩的U,V和Mo 元素,发现微生物岩中并未富集指示还原条件的U 和Mo 元素。Tang et al.[199]分析了中元古代微生物岩中的U,V和Mo元素,发现这些元素在微生物岩中较为富集,说明中元古代海水一直处于缺氧环境。Kalvoda et al.[205]分析了晚泥盆世纹层岩中的Al,Zr,Ti,Th,U 和Mo 等元素,发现了纹层岩在沉积过程中从海底氧化条件到弱氧化条件间歇性变化特征。Hüeter et al.[55]分析了OAE1a 时期特提斯海中部氧化还原敏感的微量元素As,V,Mo 和U,发现氧化还原敏感指标的变化与自生泥晶壳的扩展相吻合。
其次,自生泥晶元素地球化学特征有助于确定和区别自生泥晶的形成过程和来源。Sanchez-Beristain et al.[189]通过对意大利北部白云岩地区St Cassian组微生物岩的REE+Y 和主微量元素分析,来区分微生物岩和他生泥晶。Guido et al.[110]通过对Messinian Calcare di Base(CDB)的微生物岩REE+Y 的分析,认为CDB微生物岩是生物诱导的碳酸盐沉淀,而并非是蒸发环境下的非生物沉淀。Corkeron et al.[191]利用LAICP-MS 测量了澳洲中部新元古代Bitter Springs 组生物层中的叠层石和叠层石间隙的痕量元素特征,发现痕量元素(REE+Y,Th)地球化学可以区分这些叠层石中的细粒碳酸盐与同期非叠层石碳酸盐沉积物,并证明采样的叠层石主要是由原位沉淀形成的,大概是在微生物席/生物膜中形成的,而不是通过捕获和绑结周围的沉积物。Tanget al.[119]利用元素地球化学分析,证明了中国北方中元古代雾迷山组巨鲕的有机矿化成因。Liet al.[202]利用原位LA-ICP-MS结合岩石学和同位素地球化学技术,在华南下三叠统的海洋鲕粒中鉴定出指示古海水稀土元素(REE)组成和微生物活动的信号。Neuweileret al.[40]结合荧光能谱和稀土元素地球化学分析,发现海相方解石自生泥晶是以海水中的腐殖酸类似的有机聚合物为媒介或催化剂的,在次氧化条件下经过腐殖化作用即有机矿化作用在早期成岩阶段形成的。Desrocherset al.[45]利用沉积岩中REE+Y的分布模式特征(坡度,形状等)讨论有机聚合体的再矿化、腐殖化合物的螯合作用以及早成岩作用中铁循环的影响等,以区分自生泥晶的不同来源和沉积成岩环境。Larmagnatet al.[50]也通过稀土元素的分配模式和剖面形状来区分自生泥晶和他生泥晶。
综上所述,自生泥晶到底能不能反映同时期海水的元素地球化学特征有待于进一步的研究和验证,而自生泥晶所赋含的元素地球化学特征能否反映自生泥晶的形成过程和来源,尤其是动物来源和微生物来源的自生泥晶的区分还有待进一步研究。其实解决这些问题的关键,一方面我们需要认清自生泥晶沉淀流体的来源问题,另一方面就是自生泥晶元素地球化学赋存的微环境效应。Webbet al.[3]全面总结了微生物岩中保留的微量元素可能的四个来源,也可以作为自生泥晶中微量元素的来源:1)与生物体相关的有机富集(即生物矿化作用或有机矿化作用过程中元素的富集);2)周围水体;3)捕获和束缚的环境沉积物,包括羟基氧化物;4)后期的成岩流体。事实上,自生泥晶的形成在不同程度上反映的是微环境的效应,该种微环境可能反映也可能不反映与其形成的周围水体相同的地球化学性质。自生泥晶代表了广泛的微环境中的矿物沉淀,包括:在开阔的底栖环境中被EPS 鞘丝或ECM 包裹,分层的微生物席或生物膜,半封闭洞穴体系内的密闭环境,以及高度密闭的岩石内部环境等。所以,自生泥晶的地球化学特征所反映的环境水化学的程度也可能相应地变化(见Webbet al.[3]的总结)。当然,伴随着激光剥蚀—电感耦合等离子体质谱技术的兴起,原位微区地球化学分析成为可能,高分辨率地球化学分析更有助于区分自生泥晶的来源以及示踪周围水体地球化学的特征。
5.2 同位素地球化学
5.2.1 非金属稳定同位素
碳酸盐岩无机碳、氧同位素组成对古气候和古环境以及成岩作用过程具有较好的表征作用。碳同位素组成的变化可以通过追溯地质历史时期全球碳循环,较好地反映温室效应与冰室效应的变化,也可以充分反映有机碳产率和埋藏率的变化。而氧同位素是一种天然的同位素温度计,更容易受气候或成岩作用过程中埋藏温度的控制。此外,利用Keithet al.[206]提出的经验公式可以用来区分海相和淡水相的碳酸盐岩。前人对自生泥晶碳、氧同位素的研究较为广泛,主要是为了示踪自生泥晶的来源及其对古气候古环境的指示。无论是现代还是古代的海相微生物岩自生泥晶,它们的无机碳同位素值一般与同时期同环境的海水的无机碳同位素值是较为一致的[29-31,146,207-211]。然而,对于某些钙质微生物岩中的部分组成,可能由于在其形成自生泥晶的过程中存在异养细菌活动,可能会出现比其他微生物岩低的碳同位素值[212]。现代湖相微生物岩自生泥晶的无机碳同位素值可能受到陆源物质以及地下水等的影响,与所在水体的无机碳同位素值有些差异[21,118]。海相碳酸盐岩生物礁或灰泥丘中非微生物来源的自生泥晶(钙化动物软组织等)显示出了与同时期海水较为一致的无机碳同位素值[38,40,42,44-45,47,50,213-214]。海相鲕粒自生泥晶的无机碳同位素值也可能与同时期海水无机碳同位素值有较好的对应关系[119,179,203],而与甲烷渗漏或冷泉相关的自生泥晶的无机碳同位素值普遍偏负[58-59,113-114,142,151-152,215-216],这是与甲烷渗漏和化学自养生物的活动有关[217]。细菌硫酸盐还原反应产生的碳酸盐岩无机碳同位素值要比自生泥晶的稍微低一些,但不是很大[218]。由于受后期成岩作用改造的影响,自生泥晶的氧同位素值通常变化幅度较大,不能很好地示踪当时水体或者气候地变化。
生物活动以一种特殊的方式分离氮同位素(δ15N),使其成为可靠的生物标志和准确的古环境指标。氮所记录到的大量同位素变化与特定的新陈代谢变化有关,这些变化是对主要环境压力的直接反应,如大气中氧气的上升和海洋中硝化细菌和反硝化细菌的进化。这些同位素变化不是唯一的,而是与碳和铁的同位素变化密切相关,这表明氮可以成功地用于模拟地球历史上微生物代谢变化的相互作用,并将其与精确的环境变化联系起来[219]。自生泥晶氮同位素研究相对较少。Diazet al.[220]测试了巴哈马滩鲕粒灰岩全岩、晶间、晶内有机质的氮同位素,发现这些有几组分的氮同位素组成显示出与显示出与固氮一致的窄范围,说明微生物优先使用NO3作为电子受体。这些有机组分的氮同位素分析反应了鲕粒中微生物活动的地球化学特征。这些发现表明,氧化还原依赖的微生物群可能以鲕粒增生、胶结和泥晶化的形式影响着碳酸钙沉淀。此外,还推断出鲕粒沉积物不是反映古气候的合适指标,因为鲕粒在其一生中都受到一系列复杂的非生物和生物过程的影响,可能会导致较大的地球化学偏移。硫同位素常被用作示踪地质尺度内海洋环境中硫的生物地球化学循环过程、反演古海水的氧化还原条件变化和海洋循环模式[221-233]。碳酸盐伴生硫酸盐(CAS)的硫同位素组成通常作为古海水硫酸盐组成的指标进行测量,并为地球表面的氧化还原平衡提供信息[229,234]。绝大部分作者用他生泥晶或者泥晶基质来测试硫同位素[235-236],这是因为浅水碳酸盐岩中常见的他生泥晶中黄铁矿的比例要比自生泥晶高[236]。这种增加可能是由于化石分解过程中有机质的涌入,通过增加不稳定有机碳的可用性来促进微生物硫酸盐还原作用(MSR)237-238]。Diazet al.[220]测试了巴哈马滩鲕粒灰岩中鲕粒不同部位(外层氧化区vs.微钻孔/斑区、EPS基质内部和内核层缺氧/微需氧区)的碳酸盐伴生硫酸盐(CAS)的硫同位素组成,推断出鲕粒沉积物不是反映古气候的合适指标,因为鲕粒在其一生中都受到一系列复杂的非生物和生物过程的影响,可能会导致较大的地球化学偏移。
5.2.2 金属同位素
对自生泥晶金属同位素特征开展的研究到目前为止还较少,涉及的金属稳定同位素主要包括锶、铬和铀同位素等。放射性同位素主要是包括铅—铅和铀铅的定年。碳酸盐岩(包括自生泥晶在内的)锶同位素(87Sr/86Sr)应用的相关研究较多,也比较成熟,应用方面主要包括海相地层的定年[239-240],示踪物源[241-243]和反映原始的海相沉积环境[187-188,191,244-245]。铬是一种重要的氧化还原敏感元素,铬同位素作为反映古环境氧化还原状态的指标,对它的研究虽然不多,但已经取得了一些重要的成果[246-248]。碳酸盐岩铬同位素(δ53Cr)组成能否反映同时期海水氧化还原状态还在研究中,存在的争议也比较多[249-256]。自生泥晶铬同位素研究开展的相对较少,仅有Bonnandet al.[249]对巴哈马和尤卡坦浅滩的鲕粒灰岩的铬同位素有一些研究,Bonnandet al.[249]认为它们的铬同位素值与阿根廷盆地海水的铬同位素值一致。事实上,用碳酸盐岩铬同位素来指示海洋氧化还原状态还有诸多问题亟待解决。铀同位素体系不但广泛应用于地质年代学研究,在反演古海洋沉积环境方面的研究也得到了较大的发展[257]。根据铀的氧化还原敏感性特征,沉积物中记录的铀丰度和其同位素组成(238U/235U)可作为重建海水氧化还原模式的指标[258-261]。古碳酸盐岩中的铀同位素变化可能受海水pH、pCO2、Ca2+或Mg2+浓 度 的 变 化 控 制[262]。van Smeerdijk Hoodet al.[178]研究表明,不同的碳酸盐组构显示出238U值的高可变性,即使在同一个样本中也是如此。Hüeteret al.[55]测试了白垩纪大洋缺氧时间OAE1a时期中特特斯域Adriatic微板块碳酸盐岩台地不同组构(包括自生泥晶)的铀同位素值,认为微结壳相碳酸盐能够较好反映海水铀同位素值,并且海水铀(VI)的238U 值随着底层水缺氧空间范围的增大而减小。在放射性金属同位素方面,Kamberet al.[244]测定了津巴布韦晚太古宇Mushandike 叠层石的Pb-Pb 年龄,发现测出的年龄与锆石年龄不符。沈安江等[263]利用激光铀铅同位素定年技术测定了塔里木盆地肖尔布拉克组不同碳酸盐组构(包括藻纹层或藻叠层白云岩)的年龄,但测得的年龄与地层实际年龄有一定偏差,可能反映了成岩作用产物的年龄。此外,自生泥晶中可能赋存的其他非传统金属稳定同位素比如铁、钼、钕、钐、锂、镁、钙等在反映古环境方面的研究工作还没有得到很好的开展和应用,在可能赋存的放射性金属同位素定年等研究也亟需进一步的深入。
5.3 有机地球化学
自生泥晶中赋存有大量有机质,因而导致了其荧光显微特性。前人对自生泥晶的有机地球化学研究主要涉及自生泥晶的有机碳含量和生物标志化合物等。
自生泥晶中赋存有大量有机质,其总体有机质含量不是很高,但比其他碳酸盐岩的有机质含量要高。Neuweileret al.[38]测定了西班牙北部早白垩世(阿尔布期)碳酸盐岩灰泥丘的有机质含量,大约在0.22%~0.72%之间。Guidoet al.[109-110]测定了意大利卡拉布里亚北部晚中新世墨西拿期微生物岩的有机质含量,大约为0.06%~0.19%。
生物标志化合物的研究主要是为了示踪自生泥晶的来源及其形成过程。Keuppet al.[207]分析了德国南部晚侏罗世早启莫里期的海绵石灰岩中的生物标志化合物,分别测得了代表不同生物有机残留的标志化合物,然而不同碳酸盐岩组构(海绵残体,凝块岩结壳和碎屑沉积物)的生物标志物趋向协同,因而不能通过生物标志化合物来区别不同碳酸盐岩相。他们指出,虽然不能区别厌氧和好氧细菌以及蓝细菌在自生泥晶形成过程中的不同作用,但是从生物标志化合物中测得不同的五环烃(藿烷类)是源自细菌庚烷醇经成岩作用而形成的,意味着在有机质沉积中或沉积后真细菌活动的影响。而正构烷烃C17和C18则是代表了典型的蓝细菌来源。可以推断姥鲛烷和植烷来自叶绿素(植醇),而姥鲛烷在有氧条件下和植烷在缺氧条件下衍生。另一方面,不同的植烷/姥鲛烷关系证明海绵残体形成的缺氧条件,而凝块岩和碎屑沉积物形成条件的含氧量更高。同样,烃类角鲨烷和番茄红素似乎证明了结壳和沉积物中的厌氧古细菌活性。Reitneret al.[264]分析了美国犹他州大盐湖鲕粒和微生物岩的生物标志化合物,发现微生物岩以链长最高达21个碳原子的短链正构烷烃为主,在正庚烷显示最高峰值。随着链长的增加,正构烷烃浓度迅速降低,但在nC29周围区域的奇数同系物处表现出次峰。烃馏分的其他重要成分是短链单甲基烷烃和各种正庚烷异构体。通常在接收到大量来自光养微生物的有机物的沉积物中观察到正庚烷(nC17)和/或正庚烷类的优势。这就说明了微生物岩的蓝细菌的来源。而共同存在低分子量的中链支链单甲基烷烃是蓝细菌起源的有力支持,它们被认为是这些生物的特征性生物标记。与微生物岩相反,鲕粒砂和纯鲕粒的烃类特征主要由高分子量化合物组成。主要成分是长链正构烷烃,显示出奇数碳编号同系物的明显优势。尤其是具有27、29和31个碳原子的化合物是维管植物表皮蜡的主要成分。因此,它们在沉积物中的存在被认为是陆地衍生有机物质贡献的指示。鲕粒砂和纯鲕粒的另一个显著方面是存在高相对数量的碳链范围从C26至C35的长链单甲基烷烃。Neuweileret al.[38]分析了早白垩世(阿尔布期)碳酸盐岩灰泥丘丘核中自生泥晶的生物标志化合物,发现化合物的分布以短链正构烷烃的狭窄范围为主导,正构烷烃在nC18(正十八烷)达到最大值。这些化合物伴随有C19和C20类异戊二烯姥鲛烷和植烷。此外,在C18~C20范围内观察到大量的甲基支链烷烃和烷基环己烷。缺乏典型陆源生物标记,表明所观察到的有机物是海洋生物的来源。未分解复杂混合物(UCM)的明显峰形表明有机物异养分解(生物降解)的强烈烙印。Delecatet al.[42]分析了德国中部晚侏罗世早启莫里期牡蛎点礁中生物标志化合物,观察到的模态正构烷烃模式具有有限的生物标志物意义,因为它可能是由于有机物的热成熟而不是特定的生物来源输入产生的,但在C21至C25范围内明显出现了一组不寻常的支链烷烃。因此认为这些化合物具有微生物来源。藿烷类是沉积有机物的常见成分,是由细菌生物合成的。碳原子数超过30的烷类烃被认为是细菌细胞膜的组成成分C35细菌类六醇或C35复合生物类化合物的成岩产物。因此,沉积物或沉积岩中藿烷烃化合物的发生与细菌生物量的输入有关。从前人的研究可知,生物标志化合物分析可知自生泥晶的形成与微生物活动相关,但是对于自生泥晶的来源和形成过程的指示作用还需进一步深入研究。
6 自生泥晶在地质历史时期的分布
自生泥晶在许多古代礁建造当中广泛存在而在现在的生物礁中相对稀少。由自生泥晶和同沉积胶结物组成的叠层石曾是前寒武时期唯一的造礁生物,但在新元古代时期为凝块石和钙质微生物所补充。侏罗纪以来海水地球化学组成(钙饱和度,钙镁比)的巨大变化可能在很大程度上解释了钙化微生物、自生泥晶和平底晶洞在同一时期的急剧减少或消失的原因[265](图3)。

图3 自生泥晶在不同地质历史时期发育情况与碳酸盐岩建造组成、全球海平面变化、方解石海文石海的交替、气候变化以及生物大灭绝事件之间的关系(修改自James et al.[265];注意:*说明时间尺度发生了变化)Fig.3 Development of automicrites and their relationship to components of carbonate buildups, global sea−level changes, shift of calcite sea and aragonite sea, climate change and mass extinctions in Earth’s history (modified from James et al.[265]; Note: * indi ̄cates change of scale)
7 展望与结论
7.1 展望
正确认识自生泥晶的性质,形成过程、来源及地质历史时期的演化和分布对碳酸盐岩的结构成因分类,碳酸盐岩建造的成因分类、生物群落组成和演化,古气候和古环境的地球化学示踪,现代工程建造和环境修复,探寻地外生命和油气勘探等方面都将产生深远的影响。
(1)传统碳酸盐岩分类方法,不论是Folk[24,266],还是Dunham[100],及他们的延伸分类方法[25,267-278],都不曾将自生泥晶与他生泥晶区分开,因此很难区分真正的泥晶基质和微生物岩或其他碳酸盐岩建造中的原位泥晶。近期,Lokieret al.[172]提出了新的碳酸盐岩相分类模式,将自生泥晶和他生泥晶区分开,形成了他生(异地)碳酸盐岩(即无证据显示原始组分在沉积时是通过有机黏结作用形成的)和自生(原位)碳酸盐岩(即原始组分在沉积时是由有机黏结作用形成的)两种类型。其中,自生(原位)碳酸盐岩又根据支撑结构类型,分为黏结灰岩(生物黏结原先存在的基底)、格架灰岩(生物建造坚硬的格架)和绑结灰岩(不可识别黏结模式)三类。因此,在今后碳酸盐岩分类中需要注意区分自生泥晶和他生泥晶。
(2)自生泥晶在碳酸盐岩建造中广泛存在,在古代和现代碳酸盐岩建造中其局部贡献率可高达80%[279],但是它们却又极容易被忽视,它们的成因和来源对碳酸盐岩建造的分类,生物群落组成和演化具有重要意义[4]。前人对碳酸盐岩建造的分类和命名进行了多种尝试,大致可分为两种分类方法,一种是按照组成碳酸盐岩建造的端元组分进行的结构分类[43],另一种是按照形成碳酸盐岩建造的动力学机制进行的成因分类[28,71,265]。两种分类方法之间的最大区别就是对碳酸盐岩建造中的基质,也就是对自生泥晶和他生泥晶的有无进行区分。Bosenceet al.[280]将碳酸盐岩建造分为三种类型:格架礁、微生物灰泥丘和生物碎屑灰泥丘。Riding[43]基于碳酸盐岩建造的三个基本组构:基质(M)、(原地)骨架(S)和孔隙/胶结物(C)作为分类单元,将碳酸盐岩建造划分为基质支撑、骨架支撑和胶结物支撑三种结构。利用这个MSC 三端元组分,他又将生物礁和碳酸盐岩灰泥丘细分为:聚合微生物礁、簇状礁、节状礁、格架礁、胶结礁和灰泥丘。Jameset al.[281]以是否具抗波浪性为标准,将碳酸盐岩建造划分为礁(具抗波浪性)和丘(不具抗波浪性)。Bourqueet al.[71]强调了自生泥晶在碳酸盐岩建造中的重要性,并以碳酸盐岩建造的成因和动力学机制,即底栖碳酸盐生产的三种基本模式生物矿化作用、有机矿化作用和胶结作用为标准,将碳酸盐岩建造分为礁和生物丘体系、灰泥丘体系和胶结碳酸盐岩建造体系。Jameset al.[265]进一步强调了自生泥晶、钙质微生物、同沉积胶结物在碳酸盐岩建造中的重要性,将碳酸盐岩礁分为骨骼礁、骨骼—微生物礁、微生物礁和灰泥丘。Jameset al.[28]在碳酸盐岩礁的分类中强调了古代生物礁和现代生物礁的主要区别:1)进化生物学—现代造礁生物与化石造礁者无关,古代无脊椎动物是混合营养生物还是异养生物尚不清楚;2)钙质微生物在现代生物礁中并不重要,但在许多化石生物礁中却十分丰富;3)同沉积胶结物在岩石记录中比现在丰富得多;4)自生泥晶在许多古老的生物礁结构中普遍存在,但在今天相对罕见。因此,他们根据大于1 cm 的生物骨骼的富集程度,将碳酸盐岩礁划分为礁和礁丘两大类。其中礁包括骨骼礁、骨骼—(钙质)微生物礁和微生物礁;而礁丘则包括骨骼—钙质微生物礁丘、钙藻礁丘和灰泥丘等。在此基础上,Ridinget al.[282]根据原位碳酸盐岩的形成机理,即非生物、生物诱导和生物控制三种类型,也就是在非生物(A),微生物(M)和骨骼(S)三个端元组分的基础上,建立了碳酸盐岩建造的混合碳酸盐岩划分类型,其中单端元碳酸盐岩建造包括非生物岩、微生物岩和骨架岩,二端元碳酸盐岩建造包括非生物—微生物岩、微生物—骨骼岩和非生物—骨骼岩,部分三端元碳酸盐岩建造包括非生物—微生物和少量骨骼岩、微生物—骨骼和少量非生物岩、非生物—骨骼和少量微生物岩,完全三端元碳酸盐岩建造就是非生物—微生物—骨骼岩。但这一分类同样忽视了自生泥晶的成因和来源,并且混淆了生物控制和生物诱导碳酸盐的产物。此外,Shenet al.[4]认为识别自生泥晶的多种结构类型并正确区分不同结构类型自生泥晶的成因和来源(尤其是微生物和动物来源的自生泥晶),对研究地质历史时期碳酸盐岩建造的生物群落组成和演化,尤其是珊瑚、层孔虫海绵等群集性造礁生物出现前的前寒武—早古生代时期生命辐射和演化具有重要意义。
(3)自生泥晶,尤其是微生物岩自生泥晶,其蕴含的地球化学特征通常能够较好地反映其发育水体的地球化学组成,从而是古环境或者古气候示踪的标志[3]。然而,追寻自生泥晶在形成过程中元素富集效应和同位素分馏效应,以及自生泥晶的生物标志化合物特征,对我们厘清自生泥晶的形成机制和来源,示踪古环境和古气候具有指导作用[55]。也就是说,一方面需要认清自生泥晶沉淀流体的来源问题,另一方面就是自生泥晶地球化学赋存的微环境效应。此外,自生泥晶其他地球化学方面的应用,包括自生泥晶的原位微区元素地球化学特征,原位微区放射性同位素定年技术,非常规金属稳定同位素,高分辨率显微微区非金属稳定同位素,以及自生泥晶的生物标志化合物和脂类物质组成等方面都需要进一步加强研究和实践。
(4)自生泥晶在现代建筑、工程和生态环境修复方面也扮演着重要角色,主要是通过微生物诱导碳酸 钙 沉 淀[283](microbial induced calcium carbonate precipitation,MICP)这一形式实现的。事实上,MICP(微生物诱导碳酸盐沉淀)技术是现今生物矿化领域的研究热点之一,其主要机理是在环境中有Ca2+等金属离子以及其他底物存在时,微生物通过自身代谢或产生的EPS 降解产物来调节体系环境,形成以方解石为主的碳酸盐晶体的过程,这一过程实质上是自生泥晶形成过程的一种形式[284]。MICP 技术一方面用于建筑和工程研究领域,包括制作生物水泥、加固地基、固化土壤、防风固沙、修复裂缝、混凝土自修复和石质文物修复等[285-289],另一方面则用于环境修复包括固定土壤和水体中的重金属[290-293](Cu、Pb、Zn、Cd、Cr、As 等)。因此,认识自生泥晶(尤其是受微生物控制或诱导的碳酸盐沉淀)的形成机理和机制,对于自生泥晶的沉淀在工程和环境领域中的应用将会起到进一步促进作用,这还需要大量实验研究和证明。
(5)通过生物衍生有机质的介导形成的有机矿物(自生泥晶)可以作为生命标志,在地质岩石记录中以及在没有化石或生物建造的外星体上寻找生命的证据。Westallet al.[294]强调矿化的EPS 可作为地球或地外物质中细菌存在的标志。Reitner[295]实验表明,从Murchinson CM2 陨石中提取的有机质(很可能是非生物的)能够沉淀出与陆地微生物沉积物中已知的有机矿物相似的碳酸钙有机矿物。更普遍地说,有机分子被证明能够在无菌条件下沉淀类似生物诱导矿物的碳酸钙或磷酸盐矿物产品,尤其是那些被解释为化石的“纳米细菌”[295-296]。寻找生命起源的天体生物学研究应该在自然界中探索潜在的前生物、非生物或生物衍生有机矿物。
(6)富含自生泥晶的碳酸盐岩建造,具有一定的油气潜力[13-14]。一方面自生泥晶本身富含较多的有机质,尤其是相对深水环境的灰泥丘(例如沃尔索灰泥丘,Waulsortian mud mounds[297])以及与海底热液、油气和甲烷渗漏或冷泉相关(例如摩洛哥与热液相关的Kess-Kess 灰泥丘[298-299];墨西哥湾与冷泉石油渗漏相关的灰泥丘[300];以及其他地质历史时期的同类型灰泥丘[301])的自生泥晶,其有机质含量更高,可能为碳酸盐岩油气圈闭的形成提供良好的烃源岩[302]。另一方面,自生泥晶作为一种同沉积期成岩作用的产物,一般会降低碳酸盐岩建造的孔隙度[302]。然而,由于自生泥晶的早期固结作用会在碳酸盐岩建造中形成孔隙或格架,而且在活性生物膜的下方,细菌降解导致pH 值变化,从而导致二氧化硅和碳酸盐溶解,这样就导致了原生孔隙的扩大以及次生溶孔的发育和原生基质的局部塌陷。这一过程主要的成岩作用是海相胶结物的沉淀,而溶蚀作为生物降解的副产品,可能是为碳酸盐岩建造形成次生孔隙的一个重要过程[303]。微生物岩也能形成良好的油气储层,比如及美国阿拉巴马州阿普尔顿Little Cedar Creek 油田[304]等,但是它们原生孔隙不甚发育,孔隙度也不高,主要为由于成岩作用改造而形成的组构选择性孔隙和裂缝[305]。
7.2 结论
自生泥晶指的是原地形成的泥晶碳酸钙,它是碳酸盐岩中一个重要的不具成因指示的结构组分,微生物岩和有机泥晶都属于自生泥晶的范畴,下面总结了几个关于自生泥晶的粗略结论。
(1)现代自生泥晶以及球粒和微晶碳酸盐胶结物由高镁方解石和文石组成。自生泥晶一般是隐晶质或微晶质的。自生泥晶超微结构外貌形态主要呈现出均一的微米或纳米级别的光滑菱面体状、哑铃状、棒状、针状、晶球状、卵圆状或者纳米球状特征,它们也可以以球粒状、凝块状或者纹层状出现,形成生物层状结构、凝块球粒微结构或极细的隐晶质结构。大多数自生泥晶可以显示出不同结构类型的多泥晶组构。
(2)自生泥晶具有特殊的荧光和阴极发光显微特征,是其区别于他生泥晶的一个良好标志,但自生泥晶的这些发光特性是由什么机理决定的还有待于进一步详细研究。
(3)根据自生泥晶的形成方式和过程,可以把自生泥晶的形成机制分为无机物理化学沉淀作用、生物矿化作用和有机矿化作用三类。这些机制的作用过程以及对环境变化的响应需要在实验室和地质样品中进一步开展分析和实践。
(4)自生泥晶的地球化学特征一方面能够指示其形成机制和来源,一方面又保留了古海水地球化学组成,因而能够示踪古环境古气候变化。但是,对于自生泥晶所蕴含的元素地球化学特征、同位素分馏机理、微环境效应以及生物标志化合物的识别等需要进行更为深入的研究。
(5)自生泥晶在许多古代礁建造当中广泛存在而在现在的生物礁中相对稀少,可能与地质历史时期海水地球化学组成(钙饱和度,钙镁比)变化有关。
(6)自生泥晶在微生物岩和碳酸盐岩礁丘尤其是灰泥丘中广泛分布,也存在于与海底热液以及甲烷渗漏有关的冷泉碳酸盐岩和包括湖相沉积,泉水沉积,泉华,钙华,石灰华,钙质结壳、等陆相碳酸盐岩沉积当中。
致谢 论文在撰写过程中得到了加拿大拉瓦尔大学Fritz Neuweiler 教授和中国地质大学(北京)于炳松教授的指导。谨以此文纪念加拿大拉瓦尔大学地质与地质工程系教授Pierre-André Bourque 逝世15周年!
