片段化森林中檵木幼苗种内叶经济性状变异及其驱动因子
2021-12-02陈春何建平郑诗璐吴雁南王莹徐高福于明坚
陈春,何建平,郑诗璐,吴雁南,王莹,4,徐高福,于明坚*
(1.浙江大学生命科学学院,浙江杭州 310058;2.淳安县新安江生态开发集团有限公司,浙江 淳安 311700;3.西澳大学 生物科学学院,珀斯 6009;4.浙江大学 农业与生物技术学院,浙江 杭州 310058)
面对外界多变的环境,植物在生长与生存间进行权衡[1],为使资源获取和利用最大化,与植物资源利用策略相关的叶功能性状(又称叶经济性状,leaf economic trait)相互关联、协同变化,形成叶经济谱(leaf economics spectrum)[2-4]。叶经济谱对了解植物与环境之间的关系、群落构建和生态系统过程等具有重要作用[5-7]。以往对叶经济谱的研究大多集中于物种水平[1,8-9],但种内性状变异占总变异的25%[10],其对种群延续乃至群落构建的意义不容忽视[11-13]。近期有学者对种间叶经济谱之间的关系是否也存在于种内进行了探讨[14-18]。如LAFOREST‑LAPOINTE 等[15]以欧洲赤松(Pinus sylvestris)为研究对象,发现比叶面积与叶氮含量无关,这与WRIGHT 等[2]发现的基于物种水平的全球叶经济谱间的关系不一致。而MARTIN 等[19]通过研究咖啡(Coffea arabica)的性状指出,咖啡也存在一定的种内叶经济谱,但明显弱于物种水平的叶经济谱。生境片段化是影响生物多样性的重要因素之一,往往会导致种群丧失和适合度下降[20]。全球森林片段化现象十分严重,约有70%的森林群落处于边缘1 km 范围内;我国大多数森林斑块面积较小,小于10 hm2[21-22]。目前针对片段化生境中的种内叶经济谱的研究较少。
种内叶经济谱的形成,主要由环境因子驱动[23]。其中,光照、土壤养分和水分等生境因子的异质性是种内叶经济性状变异的重要来源[17,24-26]。例如,比叶面积随光照强度的增强逐渐降低[27]、随土壤氮元素含量的增加而增加[28]。在生境片段化过程中,面积效应和边缘效应等促使生境条件如光照、土壤性质产生强烈的变化,从而对植物产生影响[29]。种内叶经济性状的变异与协调可通过作用于个体的生长和存活,进而影响植物在片段化生境中的适合度[30],而幼苗期是植物更新的关键阶段,也是植物生活史中对生境变化最为敏感的阶段,在很大程度上决定了植物在群落内的适合度[31]。针对片段化生境中幼苗种内叶经济谱及其机理的研究,对于理解片段化生境中植物的适应和更新策略乃至群落物种多样性维持机制均十分重要。
檵木(Loropetalum chinense)为金缕梅科檵木属植物,广泛分布于北回归线以北地区,常见于我国中部、南部及西南各省[32]。檵木具有很强的适应性,可成为早期群落演替的先锋树种,对生态系统功能维持具有重要作用[33],同时还具有一定的药用价值[34]。浙江千岛湖库区是1959 年因水库大坝建设形成的片段化森林,檵木作为千岛湖库区岛屿和周边大陆马尾松林内的常见种,在千岛湖长期监测样地中其相对多度占40.48%[35]。本研究以千岛湖库区马尾松林内的檵木幼苗为对象,测定了7 种与资源获取、生长和存活紧密相关的叶经济性状,以及幼苗所处的生境条件,探讨其是否存在种内叶经济谱,并比较了大陆、大岛内部、大岛边缘、小岛4 种不同生境下檵木幼苗叶经济性状的差异,以及性状与生境条件之间的关系,旨在揭示片段化生境中檵木幼苗种内叶经济性状的变化机制,为进一步了解片段化生境中常见植物的更新策略提供基础。
1 材料和方法
1.1 研究地区概况
研究地千岛湖(新安江水库)位于我国东部的浙江省杭州市淳安县内(29°22′N~29°50′N,118°34′E~119°15′E;图1),由1959 年新安江水电站修建形成。千岛湖水域面积约540 km²,水位在108 m 时共有1 078 个面积大于0.25 hm2的陆桥岛屿。该地区属于典型的亚热带季风气候,温暖湿润、四季分明,雨热同期。年平均气温为17 ℃,年平均降水量为1 430 mm。土壤类型主要为红壤、黄壤和岩性土。在水库建成前,原有森林遭到砍伐,各岛屿在同一时间开始次生演替,至今已有60 多年,现有植被组成相似,森林覆盖率达88.5%,主要为次生马尾松林,林下多为檵木、短尾越橘(Vaccinium carlesii)、杜鹃(Rhododendron simsii)和格药柃(Eurya muricata)等物种[35]。
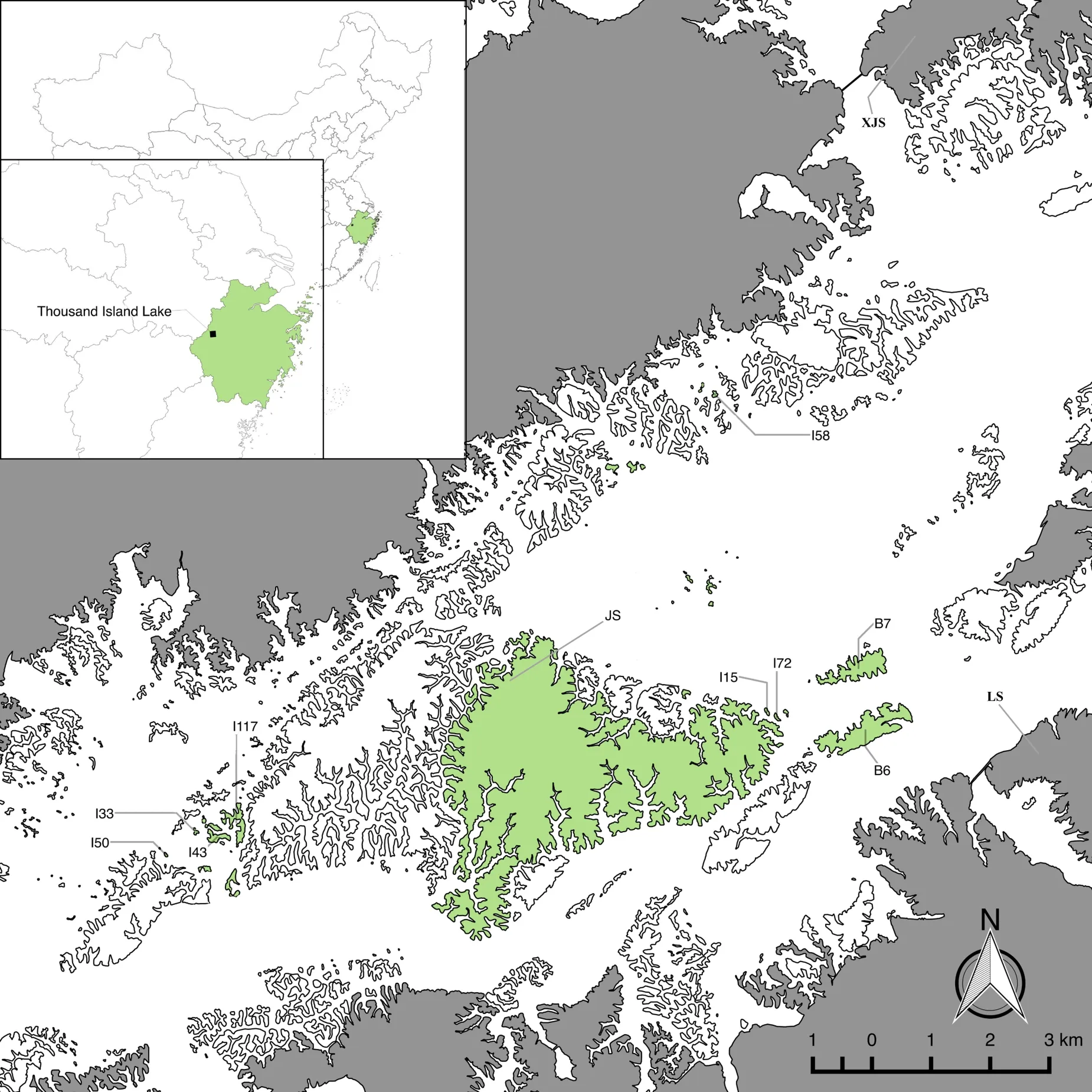
图1 岛屿与大陆样点分布Fig.1 Study islands and mainland in the Thousand Island Lake region
1.2 数据采集
1.2.1 植物样品采集与性状测定
选取5 个小岛(0.36~0.84 hm2),5 个大岛(3.70~1 153.88 hm2)和5 个大陆点(图1;附表1),小岛和大陆每个点选10 株幼苗,大岛边缘(距边缘<20 m)和大岛内部(距边缘>40 m)各选10 株,幼苗高度均<1.3 m。2019 年7 月,对每株幼苗采集5~10 片健康向阳叶片,装于密封袋中,冷藏保存,并在采集后12 h 内进行性状测定。在叶性状中,叶面积(leaf area,LA)与水分平衡、光合能力有关;叶干物质质量分数(leaf dry matter content,LDMC)与叶碳氮比(leaf carbon to nitrogen ratio,C∶N)可反映植物对外界的防御能力[36-37];而比叶面积(specific leaf area,SLA)体现植物对光的捕获效率,与叶绿素浓度(chlorophyll concentration,Chl)在一定程度上反映植物的光合能力[38-39];碳是重要的结构物质,也参与新陈代谢[40];氮与植物的最大光合效率紧密相关[36]。本研究将测定上述性状。将采集到的叶片擦拭干净后去除叶柄,称鲜重,置入扫描仪中用万深叶片分析系统(2018 版;www.wseen.com)扫描并计算叶片总面积,即为LA 乘以叶片数。用叶绿素仪(SPAD-502 Plus,Konica Minolta,Japan)避开叶脉测定每张叶片的叶绿素,测3 个点求均值,用其代表该叶片单位面积的叶绿素总浓度。叶片于60 ˚C 烘箱中烘72 h,取出称干重。LDMC 为干重与鲜重之比,SLA 为面积与干重之比,Chl 为叶绿素浓度乘以SLA。用测得的所有叶片性状值的均值表示该个体相应的性状值。各项性状测定完成后,将叶片干燥粉碎过80 目筛,称取50 mg 待测样品,用锡箔纸包裹,用元素分析仪(vario MACRO cube,Elementar,Germany)CN 模式测定叶全碳量(leaf total carbon content,C)、叶全氮量(leaf total nitrogen content,N)。
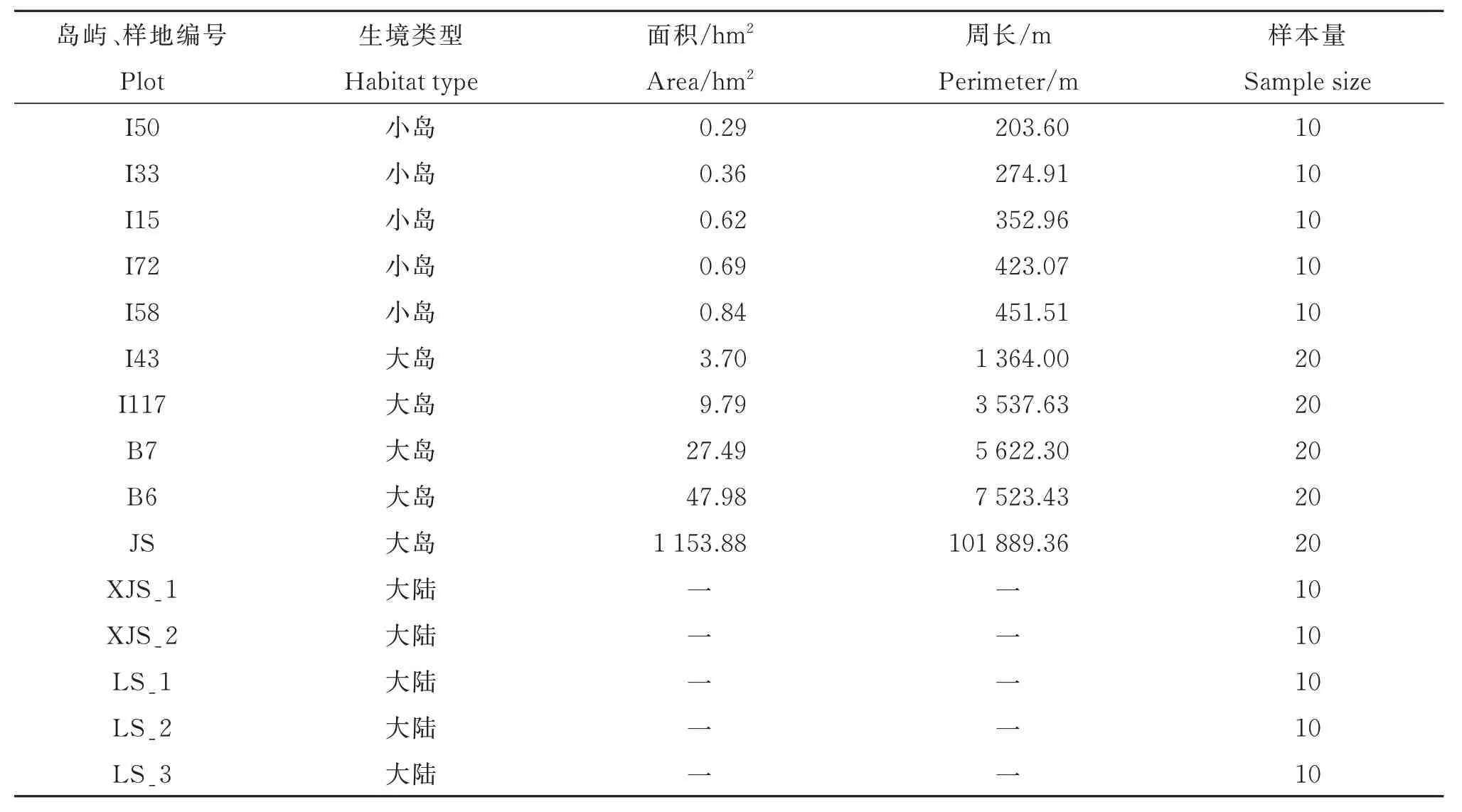
附表1 岛屿和大陆样点基本信息Table S1 The general information of study islands and mainland sites
1.2.2 生境因子测定
2019 年6 月,用装有鱼眼镜头(Sigma EX DC 4.5 mm)的相机于每个个体所在位置1.3 m 处朝上拍摄,其后用HemiView 软件(Hemiview 2.1 SR4)分析照片,得到群落郁闭度。2019 年10 月,在每个个体下方用环刀取土带回实验室测定土壤最大持水量(maximum soil water retaining capacity,MWRC),测定方法见《森林土壤水分-物理性质的测定》(中华人民共和国林业行业标准LY/T1215-1999)[41]。同时在每个个体附近3个不同方向随机选取3 个位置,去除上层枯落叶后,在土层0~20 cm 内用土钻取样,将3 个土样混合,去除大块石头和粗根系、草等杂物,装入自封袋带回实验室风干,风干后的土样放入95 ℃烘箱烘至恒重,研磨过100 目筛,称取100 mg 待测样品,用锡箔纸包裹,用元素分析仪CN 模式分析测定土壤全碳量(soil total carbon content,SCC)、土壤全氮量(soil total nitrogen content,SNC)。
1.3 数据分析
相关性采用psych 包[42]中的Pearson 相关检验分析。7 种性状标准化后(减平均值,除以标准差)通过Vegan 包[43]进行主成分分析。基于TukeyHSD 检验对4种生境中的檵木幼苗主成分1(PC1)、各叶经济性状、群落郁闭度进行多重比较,并采用独立样本T检验比较群落郁闭度在两两生境间的差异。对生境因子做Pearson 相关性分析后,因土壤全碳量与土壤全氮量之间相关系数大于0.5,故分析时剔除土壤全碳量。剩余自变量经标准化后作为固定效应因子,不同岛屿和大陆点做随机效应,用lme4 包[44]和lmerTest 包[45]拟合线性混合效应模型(最大似然估计)观察生境因子对性状PC1 及其他各性状的影响。通过MuMIn 包[46]完成模型选择。保留模型权重之和达0.95 的子模型,各参数的相对重要性由其出现在子模型的模型权重相加得到。然后进行模型平均得到参数估计值和95%置信区间。此外,线性混合效应模型分析显示,幼苗高度对性状无显著影响,加之高度非本研究的侧重点,故后续部分未涉及幼苗高度。
以上所有分析均于R 3.6.2 软件[47]中完成。
2 结果
2.1 叶经济性状相关性
7 个叶经济性状的相关性分析显示,21 对相关性中有12 对呈显著相关(表1)。其中,Chl 与LDMC、C∶N 极显著负相关,而与SLA、N 极显著正相关。LDMC 与SLA 极显著负相关,与N 显著负相关,且与C∶N 显著正相关。SLA 与N 极显著正相关,与C∶N 极显著负相关。N 与C 极显著正相关,与C∶N 极显著负相关。
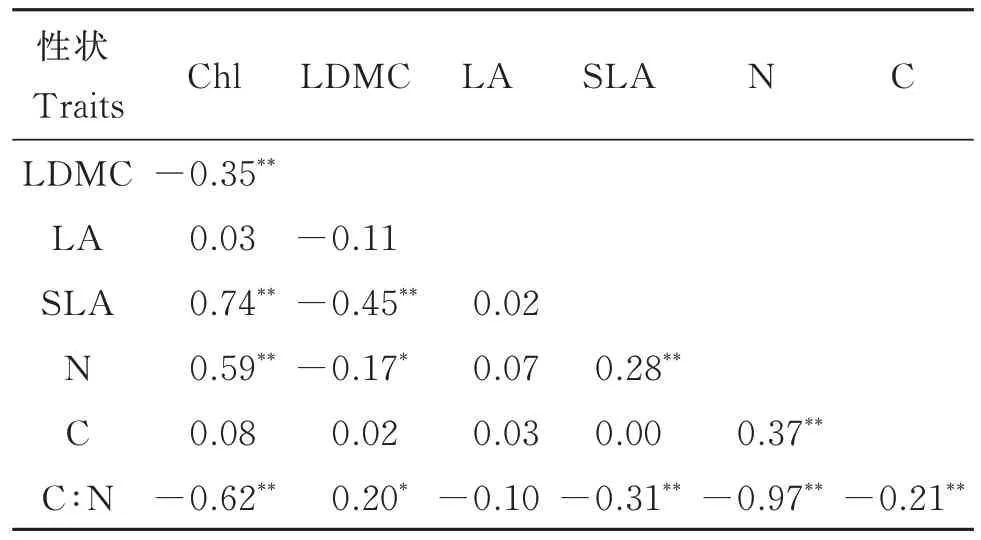
表1 檵木(Loropetalum chinense)幼苗不同叶经济性状之间的相关性Table 1 Correlations between economic traits of Loropetalum chinense seedlings on the intraspecific level
主成分分析得到的PC1 和PC2 的累计方差贡献率为62.46%(图2)。其中,PC1 的解释度为43.02%,该轴与Chl、SLA、N 呈高度正相关,与C∶N、LDMC 呈高度负相关(表2)。PC1 体现了檵木幼苗的种内叶经济谱,PC1 值越大,植株的资源利用越偏向于快速投资-收益策略。PC2 的解释度为19.44 %(图2),该轴主要与SLA 呈正相关,与LDMC 和C 呈负相关(表2)。
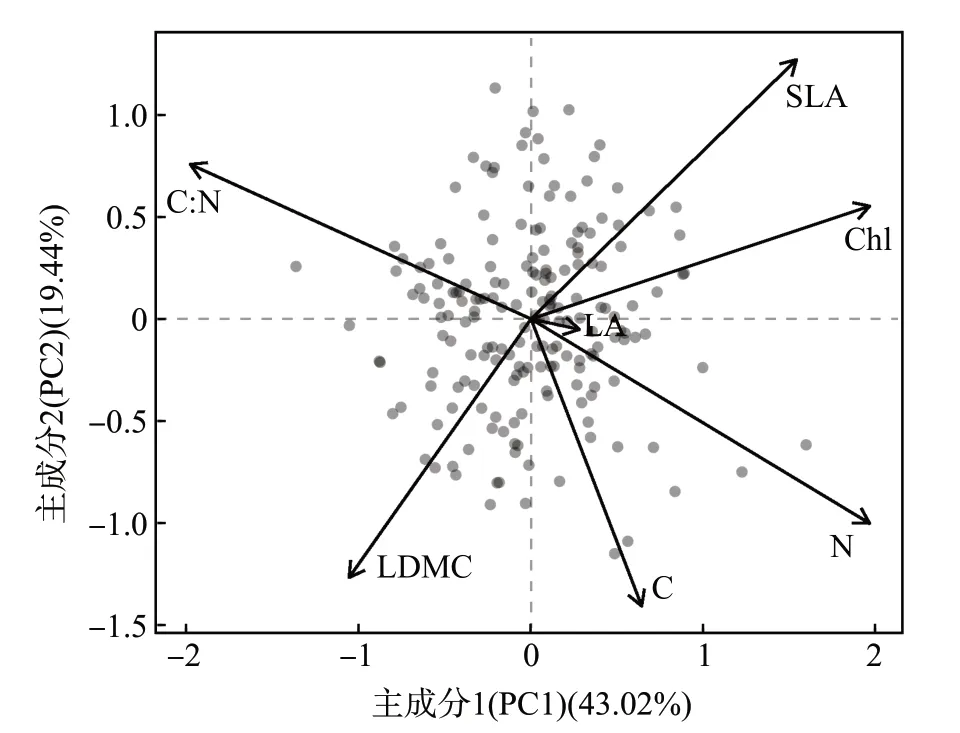
图2 檵木(Loropetalum chinense)幼苗叶经济性状的主成分分析结果Fig.2 Results of principle component analysis of the leaf economic traits of Loropetalum chinense seedlings
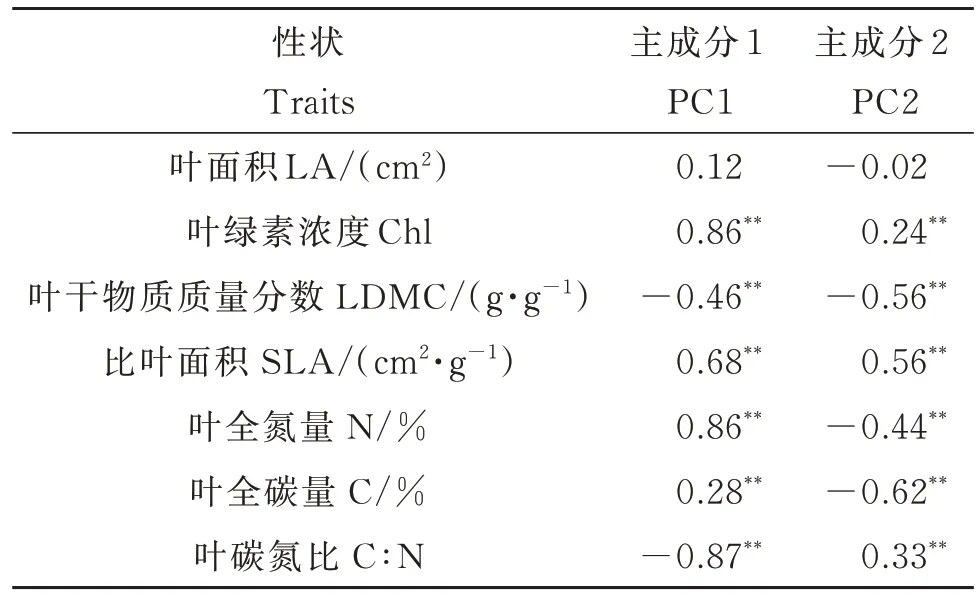
表2 檵木(Loropetalum chinense)幼苗叶经济性状与主成分的相关性Table 2 Correlations between economic traits and the principle components of Loropetalum chinense seedlings
2.2 叶经济性状随生境片段化的变化
多重比较结果(图3)显示,小岛、大岛边缘以及大岛内部的檵木幼苗叶经济性状PC1 显著低于大陆。大岛边缘的檵木幼苗叶经济性状PC1 显著低于大岛内部,而小岛与大岛边缘、大岛内部差异不显著。同时,大陆檵木幼苗的SLA、Chl、N 显著高于大岛内部、大岛边缘、小岛,C∶N 显著低于大岛内部、大岛边缘、小岛;大岛内部的Chl 显著高于大岛边缘,C∶N 略低于大岛边缘(附表2)。
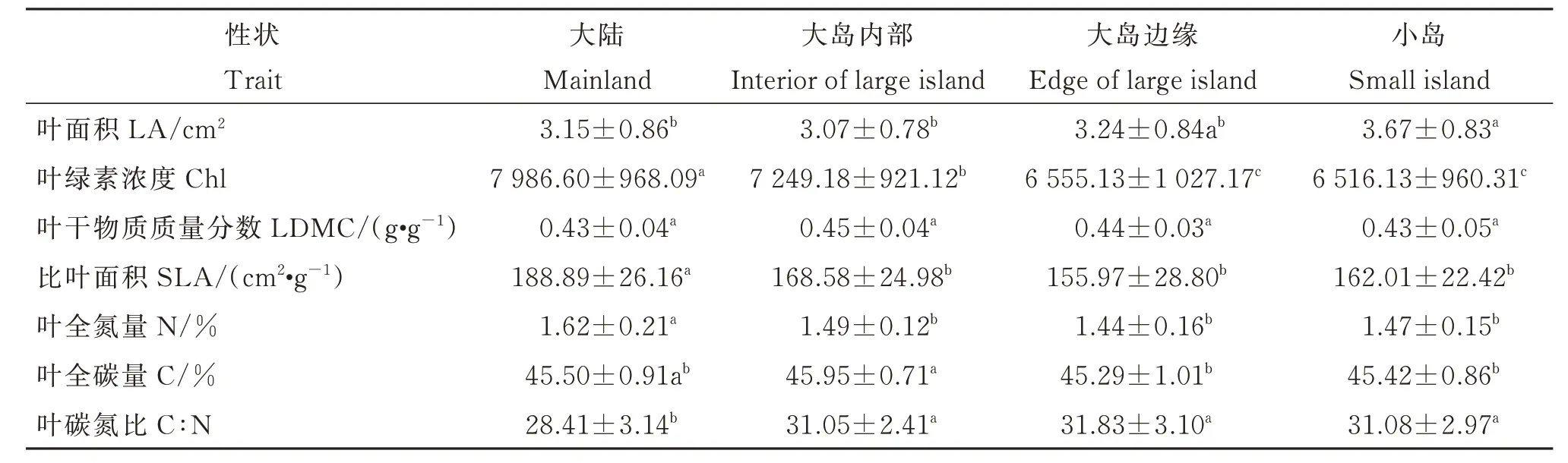
附表2 不同生境中檵木(Loropetalum chinense)幼苗叶经济性状(平均值±标准偏差)差异Table S2 Difference of the leaf economic traits(mean ± SD)of Loropetalum chinense seedlings among different habitats
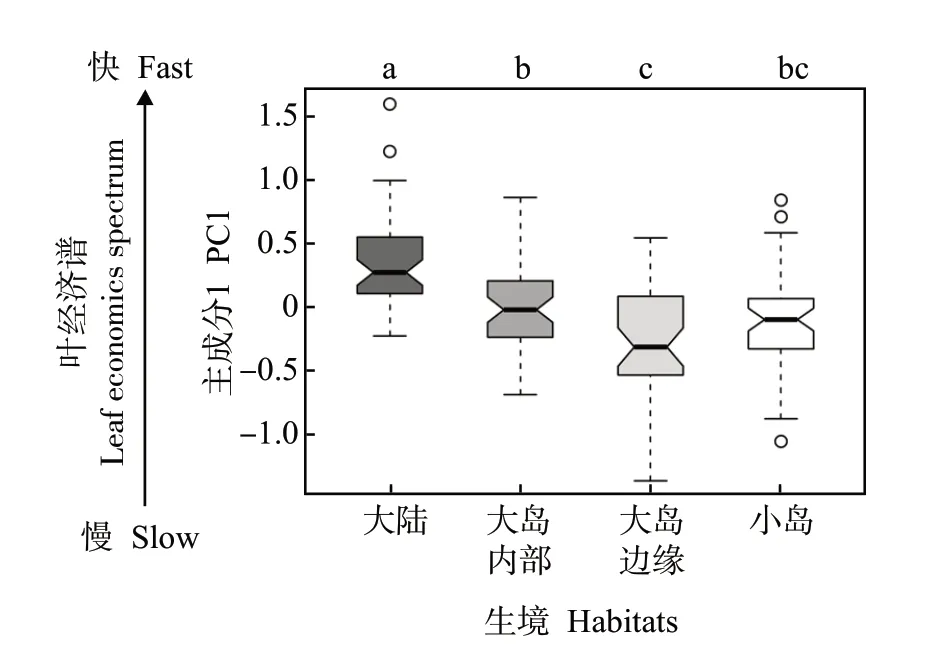
图3 不同生境下檵木(Loropetalum chinense)幼苗叶经济性状主成分1 的差异Fig.3 The difference of PC1 of Loropetalum chinense seedlings among different habitats
2.3 生境因子对叶经济性状的影响
线性混合效应模型分析显示,檵木幼苗性状PC1 受各因数的影响,从大到小依次为群落郁闭度、土壤全氮量、土壤最大持水量(图4)。其中,PC1 受群落郁闭度显著正向影响,受土壤全氮量和土壤最大持水量的影响不显著。生境因子对各叶经济性状的影响主要表现为郁闭度对LA、SLA、Chl 的显著正效应和对LDMC 的显著负效应(附表3)。同时,大陆的群落郁闭度显著高于大岛内部和小岛,而大岛内部、大岛边缘与小岛间的群落郁闭度无显著差异(图5)。T检验结果显示,大陆的群落郁闭度也高于大岛边缘(表3)。
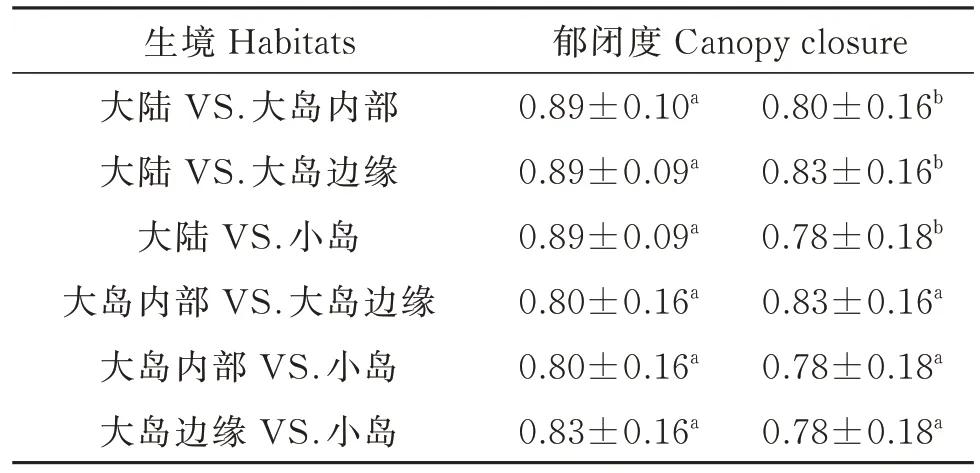
表3 不同生境之间群落郁闭度对比Table 3 The comparison of canopy closure between different habitats
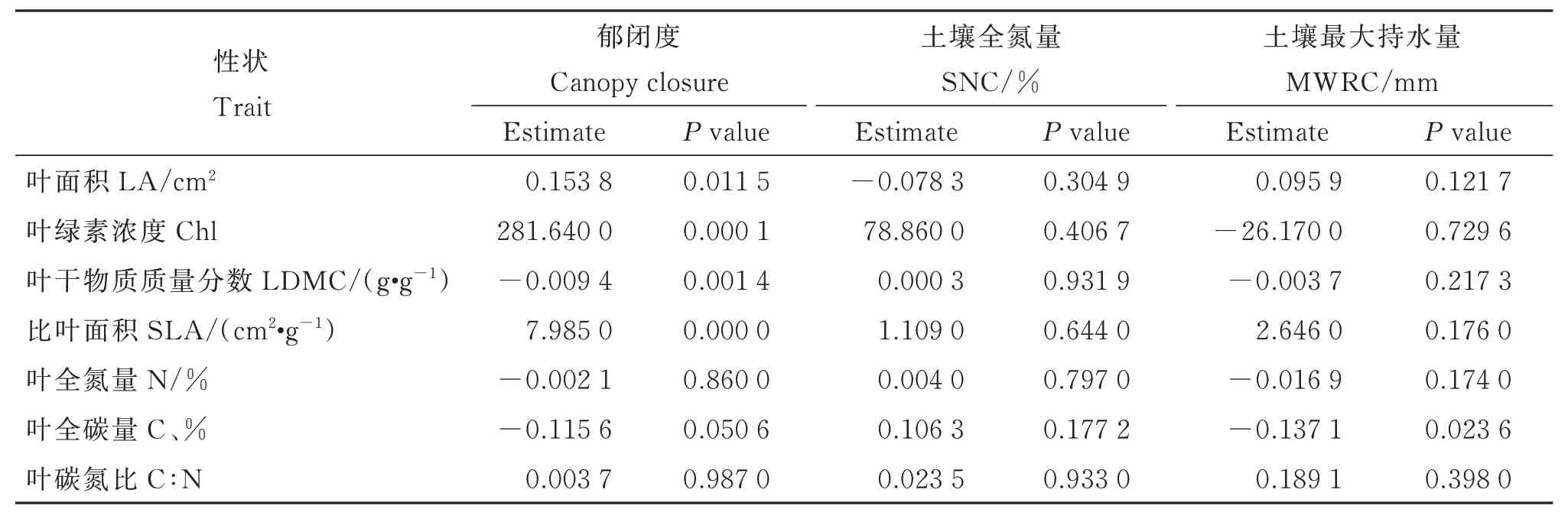
附表3 檵木(Loropetalum chinense)幼苗叶经济性状与生境因子的关联性Table S3 Correlations between leaf economic traits of Loropetalum chinense seedlings and the habitat conditions
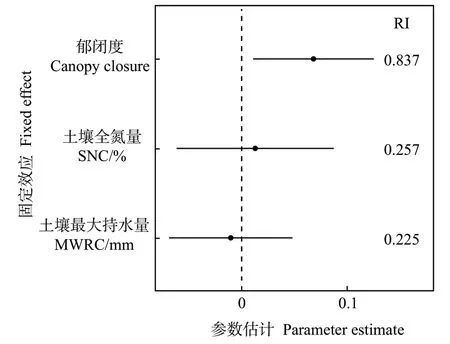
图4 固定效应因子的模型平均参数估计及95%置信区间Fig.4 Model averaged coefficient estimates and 95%confidence intervals for fixed effects explaining PC1
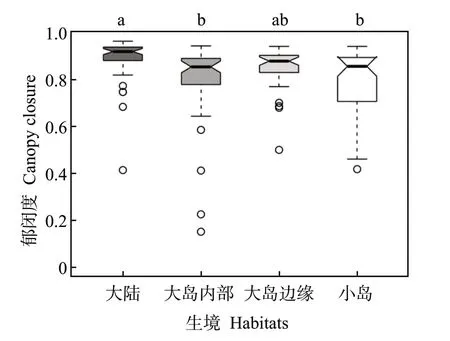
图5 不同生境下群落郁闭度的差异Fig.5 The difference of canopy closure among different habitats(P<0.05)。
3 讨论
3.1 种内叶经济谱
具有高SLA 的檵木幼苗光捕获面积大,因而具有较强的捕获光资源能力[48],Chl 和N 均为光合作用的重要物质,其中Chl 是植物光合作用的主要光合色素,可在一定程度上反映植物的光合能力[39],N是光合酶不可或缺的成分,通过影响酶含量和酶活性等影响光合作用[49],高的光合速率通常伴随高的SLA、Chl 和N[2,50],三者呈正相关关系,出现在叶经济谱的一端,代表较强的光合能力,即更侧重于生长。而高的光合速率往往伴随短的叶片寿命,在一定程度上降低了植物对植食昆虫和物理胁迫的耐受能力。相反,在经济谱的另一端,具有较高的LDMC、C∶N。LDMC 高的叶片相对坚韧,对植食动物的适口性较低,且对物理胁迫的抵抗力更强[51]。叶片中C∶N 较高表明其具有较多的防御物质(如纤维、固醇类)和较少的营养成分(如蛋白质等)。有研究表明,资源利用策略较保守的植物,通常具有较长的叶片寿命和较低的光合速率,即更侧重于生存[1]。
檵木幼苗种内叶经济性状变异间的显著协同性,主要呈现为PC1,表明存在一定程度的种内叶经济谱。具体地,PC1 与各性状的关系表明,PC1 越大,则SLA 大、N 和Chl 高,而LDMC 和C∶N 减少,植物将资源更多地用于光合生产,以使其快速生长,即具有更大的最大光合效率,更倾向于快速投资-收 益的资源利用策略;PC1 越小,则SLA 小、N 和Chl 低,而LDMC 和C∶N 增大,更有利于幼苗叶片对外界的防御,可能也具有更长的叶寿命,倾向于缓慢投资-收益的资源利用策略[2,23,52]。与以往研究相比[15,19],本研究检测出了种内叶经济性状间相对较强的关联性,这可能与生境片段化带来的较大的生境异质性有关。
3.2 片段化对生境条件及种内叶经济性状的影响
千岛湖库区的生境片段化,对马尾松林内生境条件和檵木幼苗叶经济性状具有一定影响。片段化可能通过影响生境条件进而影响幼苗性状。相对于大陆森林,岛屿森林内的檵木幼苗更倾向于采取缓慢投资-收益的资源利用策略。群落郁闭度对檵木幼苗最大光合效率具有显著正效应,大陆森林的林下光照较少,檵木幼苗具有较大的SLA 和Chl 等性状特征,以提高对光照的利用效率,达到更高的同化率,这与以往的研究结果一致[27]。相反,岛屿森林的林下光照更充足,可能也伴随较大的温度浮动等不利生境条件,在此情况下,较小的SLA(较厚的叶片)有利于减少水分散失等以抵御高温干旱胁迫。这一系列的性状特征改变,导致岛屿生境中的檵木幼苗形成了较为保守的资源利用策略,叶片可能较为结实耐久[38],进而更好地适应片段化环境,在经济谱中偏向于缓慢投资-收益的资源利用策略。
以上结果符合叶经济谱中的环境因子主导假说[23]。
岛屿中,大岛内部的最大光合效率更高,表明大岛内部的檵木幼苗更倾向于快速投资-收益的资源利用策略,而大岛边缘更倾向于缓慢投资-收益的资源利用策略。这可能与檵木幼苗对昆虫植食的防御有关。GONZALEZ 等[53]在探究片段化森林中边缘效应对节肢动物的影响时指出,边缘的植食昆虫 丰富度更高;GUIMARAES 等[54]通过整合分析片段化对植食昆虫多度、丰富度和植食的作用指出,森林边缘具有较高的植食昆虫多度和丰富度,且边缘植物比内部植物的植食率高70%;而MALDONADO-LOPEZ 等[55]也发现,森林边缘的瘿蜂具有较高的多度和丰富度。大岛边缘的檵木幼苗可能更多地将资源分配到防御性状上,为更好地存活而采取缓慢投资-收益的资源利用策略。小岛与大岛间檵木幼苗叶经济性状差异不显著,表明边缘效应可能较面积效应对植物叶经济性状的影响更大。
与以往研究不同[25,56],土壤全碳量、土壤全氮量和水分条件对千岛湖片段化岛屿及周边连续森林中的檵木幼苗性状无显著影响,这可能与千岛湖片段化景观基质是水有关,使水分不再是限制性因子。而HAN 等[57]通过对753 种植物的氮磷比研究指出,我国陆地植物生长主要受土壤磷的限制,这可能是导致檵木幼苗叶经济性状不受土壤全碳量、全氮量影响的原因。
4 结论
檵木幼苗种内叶经济性状并不独立存在,面对片段化森林中的多变生境,檵木在更新阶段形成了一套能适应特定环境的叶经济性状组合,从而调整其资源利用策略,形成了种内叶经济谱。片段化可能通过影响生境条件进而影响植物的叶经济性状,而随着生境片段化的加剧,檵木幼苗叶逐渐倾向于缓慢投资-收益策略。本研究一定程度上丰富了植物叶经济谱的理论,在开展基于功能性状的物种共存研究时对是否需要考虑种内变异具有一定的指导作用。也提示在研究片段化生境中群落构建和生物多样性维持机制时应考虑种内叶经济谱的影响。
后续研究需同时测定叶脉密度、气孔密度等水力性状以确定主成分2 是否与植物的水分利用策略等相关,且需同时考虑生物因子对叶经济性状的影响;并在条件允许的情况下尽可能增大取样量,以使结果更有说服力。
贵州师范大学金毅,浙江大学张爱英、巫东豪对论文提出了修改建议,淳安县新安江生态开发集团有限公司在项目实施过程中给予配合与支持,当地村民对野外工作给予帮助,在此一并致以诚挚的谢意!
