利用数字图像分析132 份胡枝子种子表型性状遗传多样性
2021-11-22杨正禹陆忠杰张茂董瑞
杨正禹,陆忠杰,张茂,董瑞
(贵州大学动物科学学院,贵州 贵阳550025)
胡枝子属(Lespedeza)为豆科(Leguminosae)多年生草本、灌木或半灌木,全世界共有本属植物100 余种,我国约65 种,在全国各地广泛分布[1−2]。其具有蛋白质含量高、适口性好、抗逆性及水土保持能力强等优点[3−5],富含生物碱、黄酮、萜类、缩合单宁等医用化学物质,其中缩合单宁能够促进反刍动物的消化[6−8]。胡枝子(L. bicolor)还是中国草种子出口量较多的优良饲草之一,每年出口量达50~80 t[9]。其在畜牧业、医药及水土保持方面都得到了广泛的应用。
种质资源是育种工作和物种开发的基础,丰富的种质资源对于物种的收集、保存及改良具有重要意义[10]。物种的表型性状多样性是种质资源遗传变异的外在表现,受环境及遗传综合作用,是最基础、直接和便于掌握的生物学特性[11−13],也是植物遗传多样性研究及衡量物种多样性的重要指标和主要内容,能直观反映植物多样性[14]。前人对同一物种不同种质性状存在的变异进行了广泛深入研究,如红砂(Reaumuria soongorica)[15]、箭筈豌豆(Vicia sativa)[16]、大豆(Glycine max)[17]、苜蓿(Medicago sativa)[18]、小麦(Triticum aestivum)[19]等。
植物种子与其他器官相比,所受的外部压力影响更小,形态更稳定,具有易收集、储存时间长等多种优点,同时对幼苗的生长发育也有很大影响[20−22]。种子的寿命、萌发等也与种子的形态特征存在着密切的联系[23]。因此,种子形态特征在分类学、物种遗传变异等方面均具有重要意义,特别是在物种的鉴定和区分方面[24−25]。Apuan等[26]使用几何形态计量学(geometric morphology,GM)工具进行图像分析,将杂草稻与栽培水稻(Oryza sativa)种子性状进行比较,确定了杂草稻与栽培稻之间的关联和关系。童跃伟等[27]利用种子的数字图像分析了6 个种群红松(Pinus koraiensis)种子表型分化程度和变异规律。
本研究对132 份胡枝子属种质的种子形态学特征、基因型变异进行分析,旨在对胡枝子的种子表型性状进行全面了解,为胡枝子遗传改良提供现实依据。
1 材料与方法
1. 1 试验材料
试验材料为132 份胡枝子属种质种子(表1)。其中131 份种质由美国国家植物种质系统(National Plant Germplasm System of the United States,NPGS)提供,1 份为中国野生种。

表1 胡枝子种质信息表Table 1 Number distribution of Lespedeza germplasm
1. 2 试验方法
本研究于2019 年7 月进行,分别对L. juncea(7份)、L. cuneata(42 份)、L. capitata(51 份)、L. bicolor(10 份)4 个种及全部132 份胡枝子种质进行分析。共测量7 个种子性状,分别为种子长(seed length,SL)、种子宽(seed width,SW)、长宽比(width to length ratio,SL/SW)、种子周长(perimeter,Pe)、种子厚(seed thick,ST)、种脐长(hilum length,HL)、百粒重(100-seed weight,SY)。百粒重为100 粒种子重量,其余性状每份种质测量30 粒种子,3 次重复。
利用扫描仪(EPSON,印度尼西亚)对每份种质种子(每份种子30 粒,3 次重复)进行扫描,获得种质的种子图片,图像分辨率为300 dpi,像素为1024×1024。使用WinSEEDLETM 2011 图像分析系统对种子图像进行分析,得到种子长(SL)、种子宽(SW)、长宽比(SL/SW)、种子周长(Pe)。使用数显游标卡尺(AIRAI SR44,精度0. 01 mm,山东青岛)测量种子厚度(ST)、种脐长度(HL),电子天平(Mettler Toledo-MS204S,精度0. 0000 g,瑞士)称量种子百粒重(SY)。
1. 3 数据分析
数据分析基于:1)利用方差分析来评估胡枝子种质间各表型性状基因型变异的显著性和大小。2)利用模式分析,包含聚类和主成分分析(principal component analysis,PCA)[28],以图形方式来展现所有种质各性状数据矩阵。
在GenStat 7. 1(2003)中使用残差最大似然(residual maximum likelihood,REML)进行方差分量分析。随机线性模型用于分析胡枝子种质间及种间性状变异。利用最佳线性无偏估计值(best linear unbiased predictors,BLUP)[29]分析,来获得各种质种子的性状均值,再利用均值进行主成分和聚类分析。其中方向向量表示各性状之间的相关性。还进行了以胡枝子不同种质和种作为固定效应的混合线性模型分析,以研究胡枝子不同种质和种之间的性状差异。
使用REML 分析中的基因型(σg2)和实验误差(σε2)的方差分量以及nr(重复次数)估算每个种质各性状的平均重复性(R)[30]。用变异系数(CV)表示各性状离散程度。

式中:S为标准差,X为平均值。
各种子性状之间的表型相关性(rp)使用SPSS 20. 0 分析。
2 结果与分析
2. 1 表型性状多样性分析
对胡枝子属不同种的种子表型性状分析发现,性状SL、SW、ST、HL、SY、Pe 在L. bicolor中最大,分别为3. 156 mm、2. 469 mm、1. 562 mm、0. 697 mm、0. 558 g 和8. 950 mm。性状SL、SW、SY/SW、HL、SY 和Pe 在L.cuneata中最小,分别为1. 747 mm、1. 414 mm、1. 238 mm、0. 562 mm、0. 163 g 和4. 880 mm(表2)。 表明L.bicolor的种子最大,而L. cuneata的种子最小。132 份胡枝子种质间各性状均存在显著(P<0. 05)差异。在L.juncea、L. cuneata、L. capitata和L. bicolor4 个种中,除SL/SW 外,其余性状均具有显著(P<0. 05)差异,表明胡枝子种子表型性状存在丰富的基因型变异。所有种质中除SL/SW 外,其余性状的CV 均较高,其中SY 高达61. 53%。在L. juncea、L. cuneata和L. bicolor中,SY 的CV 最高,分别为61. 95%、20. 60% 和40. 63%。上述结果表明胡枝子种间及种内的种子表型性状多样性均较高。

表2 胡枝子种质估计基因型、实验误差方差分量及相关标准误差评估Table 2 Estimated genotype,experimental error variance component and related standard error evaluation of Lespedeza germplasm
在132 份胡枝子种质间,性状SL、SW、ST、SY 和Pe 具有高的性状平均重复性(R)值,分别为0. 983、0. 984、0. 979、0. 997 和0. 990,性状SL/SW 和HL 具有中等R值,分别为0. 677 和0. 783。在L. juncea中,性状ST、SY 具有高R值,分别为0. 942 和0. 999。在L. cuneata和L. capitata中,仅SY 具有高R值,分别为0. 968 和0. 985。在L. bicolor中,SL、SW、HL、SY 和Pe 具有高R值,分别为0. 965、0. 944、0. 920、0. 999 和0. 980。所有种中性状SY均有高的R值,表明性状SY 遗传性稳定,不易受环境影响,可作为品种选育的重要参考性状。
2. 2 种子形态特征模式分析及表型相关性
在132 份胡枝子的7 个性状中(图1),性状ST、SW、SY、SL 和Pe 之间呈正相关(方向向量间角度<90°),且SW 与SY 呈强正相关。而性状SL/SW与HL、ST 呈负相关(方向向量间角度>90°)。主成分分析显示,主成分1 贡献率为68. 85%,主成分2 贡献率为15. 95%,累计达84. 80%。

图1 132 份胡枝子种质7 个性状的主成分分析及聚类分析Fig. 1 Principle component analysis and cluster analysis for seven traits of 132 Lespedeza germplasm
聚类分析将132 份胡枝子种质分为3 个组群(表3),第1 组包含63 份种质,第2 组包含52 份种质,第3组包含17 份种质。性状SL、SW、ST、SY 和Pe 在第3组中存在最大值,性状SL/SW 在第2 组中存在最大值,7 个性状均在第1 组中存在最小值。

表3 132 份胡枝子种质聚类分析分组信息Table 3 Cluster analysis grouping information of 132 Lespedeza accessions
表4 中132 份胡枝子种质各性状间的表型相关性(rp)系数进一步证明了图1 中ST、SW、SY、SL 和Pe之间呈正相关,且差异极显著(P<0. 01)。正rp的最小值在SL/SW 与SY 间,为0. 058。 最大值在SL 与Pe 间,为0. 991。
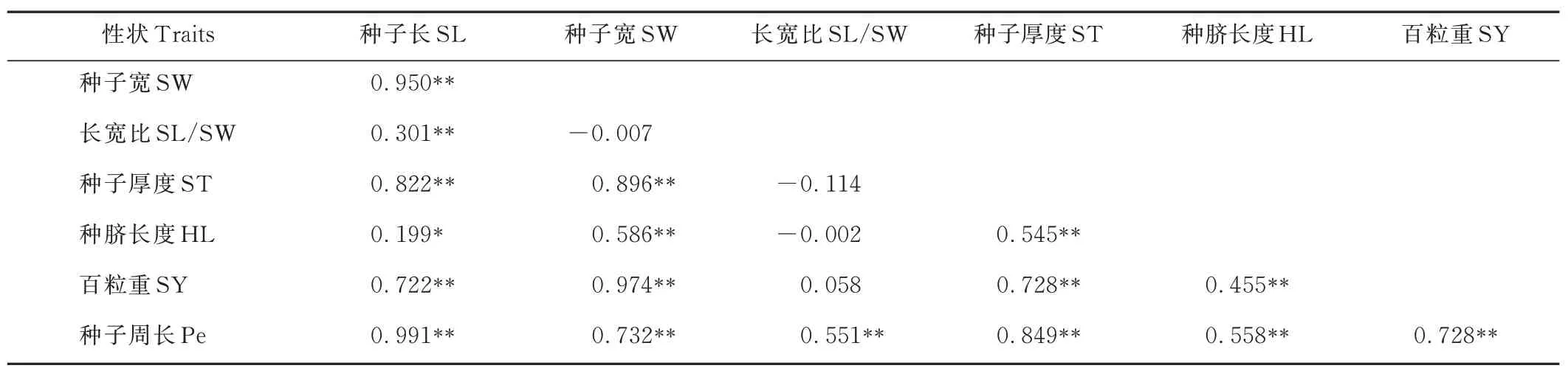
表4 132 份胡枝子种质各性状间表型相关性系数Table 4 Phenotypic(rp)correlation coefficients among traits based on the 132 Lespedeza accessions
在L. juncea中(图2),性状ST 与HL,Pe 与SL 均呈强正相关。SY 与ST、HL 呈正相关(方向向量间角度<90°)。性状SW 与SL/SW,SL/SW 与Pe、SL 均呈负相关(方向向量间角度>90°)。主成分分析显示,主成分1 贡献率为43. 25%,主成分2 贡献率为36. 57%,累计达79. 82%。
对L. juncea进行聚类分析,可将其分为2 个组群(表5)。第1 组包含3 个种质,第2 组包含4 个种质。第1 组各性状均值除ST 外,其余均大于第2 组。

表5 L. juncea 聚类分析分组信息Table 5 Cluster analysis grouping information of L. juncea
相关性系数(rp)进一步验证了图2 所示的性状Pe与SL,ST 与HL,SW 与Pe 均在显著(P<0. 05)水平下呈正相关(表6)。正rp的最小值在SW 与SY 间,为0. 082,最大值在SL 与Pe 间,为0. 871。

表6 7 份L. juncea 种质各性状之间的表型相关性系数Table 6 Phenotypic(rp)correlation coefficients among traits based on the 7 L. juncea accessions

图2 L. juncea 7 个性状主成分分析及聚类分析Fig. 2 Principle component analysis and cluster analysis for seven traits of L. juncea
在L. cuneata中(图3),性状HL 与SL/SW,ST与SY 间均呈强正相关(方向向量间角度<90°),而性状SW 与HL、SL/SW、SY、ST 呈负相关。主成分分析结果显示,主成分1 贡献率为42. 94%,主成分2 贡献率为23. 69%,累计达66. 63%。

图3 L. cuneata 7 个性状的主成分分析及聚类分析Fig. 3 Principle component analysis and cluster analysis for seven traits of L. cuneata
聚类分析将42 份L. cuneata分为2 个组群(表7),第1 组包含23 个种质,第2 组包含19 个种质。由性状均值可看出,第2 组除性状SY 外,其余各性状均值均大于第1 组。

表7 L. cuneata 聚类分析信息Table 7 Cluster analysis grouping information of L. cuneata
L. cuneata各性状间表型(rp)相关性系数进一步证明了性状ST 与SY、HL 与SL/SW 均在极显著(P<0. 05)水平下呈正相关(表8)。性状Pe 与SL、ST 之间也存在极显著(P<0. 01)正相关。HL 与SL/SW 间rp最大,为0. 893,最小rp在HL 与ST 之间,为0. 002。
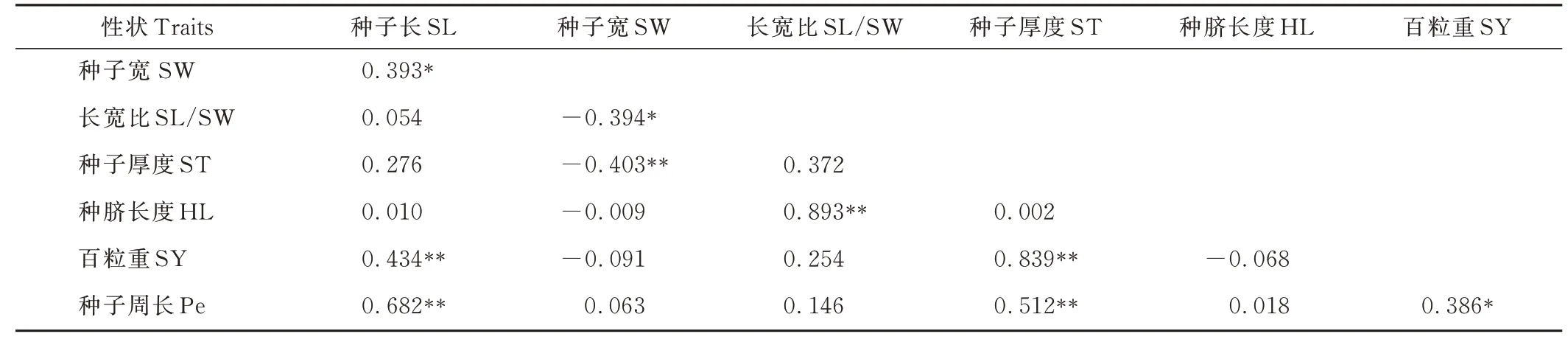
表8 42 份L. cuneata 种质各性状之间的表型相关性系数Table 8 Phenotypic(rp)correlation coefficients among traits based on the 42 L. cuneata accessions
在L. capitata中(图4),性状SY 与SW 呈强正相关(方向向量间角度<90°),HL 与SY、SW 间呈正相关。性状ST 与SY、SW、HL 呈负相关(方向向量间角度>90°)。主成分分析显示,主成分1 贡献率为43. 98%,主成分2贡献率为23. 63%,累计达67. 61%。

图4 L. capitata 7 个性状的主成分分析及聚类分析Fig. 4 Principle component analysis and cluster analysis for seven traits of L. capitata
聚类分析将51 份L. capitata分为2 个组群(表9)。第1 组包含28 份种质,第2 组包含23 份种质。性状HL 与SY 在2 个组群中的均值相等,第1 组除性状SL/SW 外,其余性状均值均小于第2 组。

表9 L. capitata 聚类分析信息Table 9 Cluster analysis grouping information of L. capitata
L. capitata各性状之间的表型(rp)相关性系数进一步证明了SY 与SW 在极显著(P<0. 05)水平下正相关(表10)。 性状SL 与SW、SL/SW、Pe,SW 与HL、SY,SL/SW 与Pe,ST 与Pe 间均存在极显著(P<0. 01)正相关。 性状SW 与ST 在极显著(P<0. 01)水平下呈负相关。正rp最小值在SL/SW 与SY之间,为0. 050,最大值在SY 与SW 之间,为0. 945。

表10 51 份L. capitata 种质各性状之间的表型相关性系数Table 10 Phenotypic(rp)correlation coefficients among traits based on the 51 L. capitata accessions
在L. bicolor中(图5),性状SL、SL/SW、Pe、ST 和HL 之间呈正相关(方向向量间角度<90°)。性状SY 与SW、SL/SW 呈负相关(方向向量间角度>90°)。主成分分析显示,主成分1 贡献率为60. 53%,主成分2 贡献率为19. 80%,累计达80. 33%。

图5 L. bicolor 7 个性状的主成分分析及聚类分析Fig. 5 Principle component analysis and cluster analysis for seven traits of L. bicolor
聚类分析将10 份L. bicolor分为2 个组群(表11)。第1 组包含4 份种质,第2 组包含6 份种质。两个组群的各性状均值除HL 相等外,其余性状均值第2组均大于第1 组。

表11 L. bicolor 聚类分析信息Table 11 Cluster analysis grouping information of L. bicolor
L. bicolor各性状之间的表型(rp)相关性系数进一步证明了性状Pe 与ST、SL,SL 与ST 在极显著(P<0. 01)水平下呈正相关,SL/SW 与HL、SL 与SW 在显著(P<0. 05)水平下呈正相关(表12)。正rp的最小值在SL 与SY之间,为0. 049,最大值在SL 与Pe 之间,为0. 997。

表12 10 份L. bicolor 种质各性状之间的表型相关性系数Table 12 Phenotypic(rp)correlation coefficients among traits based on the 10 L. bicolor accessions
3 讨论
自然资源的遗传变异是经过长期自然选择形成,从而获得了相对稳定且丰富的表型变异特征[31]。本研究对胡枝子属132 份种质及4 个种L. juncea、L. capitata、L. bicolor和L. cuneata的7 个种子性状进行分析。结果表明,除性状SL/SW 外,其余种子性状都存在显著(P<0. 05)的基因型变异,这些种子性状的基因型变异和R值表明,在各种质内存在潜在可用的遗传变异。张吉宇[32]对14 个胡枝子属野生居群植株形态特征进行了遗传多样性研究,发现胡枝子的表型性状丰富。Bécquer 等[33]和Ocampo 等[25]也从不同植物种子数字图像中提取了长度、宽度、周长以及种子形状等数据来分析其遗传多样性和物种分类。本研究中各种子表型性状的CV 均较高(>5%)。有研究表明,性状的CV 超过5% 可视为产生了较大变异[34],而CV 越大说明植物对于环境的适应能力越强,能适应的环境范围也越广[35]。本研究中132 份胡枝子种质CV 均较高,这是由于132 份胡枝子种质来自全球不同地区,经过了长期的地理隔离及不同海拔和环境气候的影响。而环境条件是种质资源遗传变异的主要因素之一。
根据R值可判断各表型性状的遗传稳定性。L. juncea中性状SL、SL/SW、Pe、HL、SY 和ST 具有较高(R>0. 7)的R值;L. cuneata中性状SW、Pe、HL、ST 和SY 具有较高的R值;L. capitata中性状SL、SW、Pe、SY 和ST具有较高的R值;L. bicolor中性状SL、SW、Pe、HL、SY 和ST 具有较高的R值。性状的R值越高表明其受遗传因素的影响越大,越不易受环境因素影响,遗传更具稳定性。L. bicolor各性状的R值均高于其他种,表明L.bicolor与其他种相比遗传更稳定。在4 个种中,性状SY、ST、Pe 的R值均较高(R>0. 9),相比其他性状,其遗传稳定性较好,不易被环境因素所改变,能够在后期的繁育过程中稳定遗传,保持优良性状。
种子表型相关性分析表明,性状SL/SW 与SY、Pe 之间相关性较弱,而种子的SL/SW 主要用来描述种子的形状,与种子的大小性状无关,因此胡枝子种子大小与种子形状独立遗传,这与对青稞(Hordeum vulgare)[36]、小麦[37]的研究结果相似。主成分分析结果表明,在132 份胡枝子种质中,第1 主成分解释了总变异的68. 85%,第2主成分解释了15. 95%,累计达84. 80%,说明这两个主成分可反映大部分信息。第1 主成分包括SL、SW、ST、SY、Pe,第2 主成分包括HL、SL/SW。表明第1主成分主要描述种子大小,第2 主成分主要描述种子形状。 杨生华等[38]和侯维海等[36]分别对蚕豆(Vicia faba)和青稞种子表型性状进行的主成分分析都表明第1 主成分涵盖了籽粒大小信息,第2 主成分涵盖了籽粒形状信息。在特定环境中物种种子大小对其幼苗的建植和存活具有显著影响。因此,植物种子性状遗传变异的信息可为新品种选育提供重要的数据,尤其是对种子产量的提高以及幼苗建植和存活情况的改良具有十分重要的意义[39]。在本研究中,胡枝子属各种质间和种内种子性状均具有显著的(P<0. 05)基因型变异,这些性状可用于胡枝子属植物分类学研究。
4 结论
种子丰富的表型性状信息对于优良品种的选育及保存具有重要意义,且种子的大小对植物幼苗的建植和存活具有显著影响。本研究结果表明,胡枝子种质及种间均存在显著的基因型变异,表明胡枝子各种质种子遗传多样性丰富。性状SY、ST 和Pe 遗传稳定,可作为后期品种选育的参考指标。这不仅为胡枝子种质资源的收集、保存提供基础数据,还有利于胡枝子改良育种工作的开展。
