后基因组时代基于选择导入系的水稻设计育种策略
2021-11-22石英尧马赛曾威郝芷圻李思敏王文生黎珉
石英尧, 马赛, 曾威*, 郝芷圻, 李思敏, 王文生, 黎珉*
(1.安徽农业大学农学院, 合肥 230036; 2.中国农业科学院作物科学研究所, 北京 100081)
水稻(OryzasativaL.)是我国乃至世界上较为重要的粮食作物之一,是全世界超过30%以上的人口的主要食物来源。我国人口在未来20年仍将继续增长,对粮食的需求持续增加,但我国耕地面积却在不断减少,未来10年单产必须提高15%以上才能确保我国的粮食安全[1-2],因此,水稻的安全生产对保障我国粮食安全具有重要的战略意义。水稻的两次“绿色革命”使产量取得了两次重大飞跃[3]:20世纪60年代,矮化育种的“绿色革命”解决了大量施肥导致的植株倒伏和减产问题,水稻单产大幅度提升;20世纪70年代,杂交水稻的三系配套将水稻产量又推向了一个更高的水平。传统水稻育种主要分为优异亲本组配获得育种群体、优良后代选择、优良后代培育为新品种[4]。
进入21世纪,水稻生产面临着新的挑战,不断增长的人口与耕地面积和水肥资源的矛盾日益突出。随着全球气候环境不断恶化,自然灾害如干旱、盐渍、高温热害、病害、土壤贫瘠严重影响了粮食安全[5]。传统育种消耗大量的人力、物力,一般需要6~8年才能培育出一个优异品种,难以满足水稻育种的需要。复杂数量性状(如产量和抗逆性)一般由微效多基因控制,同时受遗传背景和环境影响,功能基因组学成果难以直接应用于育种实践中,如何突破传统遗传育种技术的瓶颈,充分利用水稻种质资源的遗传变异,设计不同生态地区的水稻新品种所需要的广适性品种已经成为亟需解决的重大科学问题。
黎志康等[6-7]提出选择导入育种(breeding by selective introgression, BBSI)的理念和策略,用于复杂性状的遗传剖析和新品种改良。BBSI已通过“为亚非脱贫培育推广绿色超级稻”重大国际合作项目广泛实施,有效地将种质资源与基因发掘、目标性状定向改良和品种选育过程结合[8]。其团队以200余份核心种质资源为供体,应用BBSI技术构建了大量优良骨干亲本背景的回交群体和选择导入系,多个新品种通过国交和省级审定的水稻新品种,推广面积超过600万 hm2。同时,获得了大量表现优异的选择导入系(selective introgression lines, SILs),成为培育绿色超级稻的基础材料,为后续分子设计育种奠定了基础[9]。近年来,水稻功能基因和群体基因组的研究使精准育种成为可能[10-11],也称为基因组信息育种,或设计育种(breeding by design, BBD)[12]。随着水稻基因组不断完善,如何将大量的遗传变异信息与育种结合,培育适合不同环境的优良新品种是水稻分子育种面临的新挑战。本文阐述了BBSI研究理念及策略的发展过程及其分子设计育种中的运用,并探讨了如何利用分子数量遗传理论和全基因组变异信息来验证并丰富该策略,包括BBSI对在其它作物育种中实施的可能性。
1 选择导入育种策略的提出
回交(backcross, BC)育种是将一个高遗传性状转入优良品种的方法[13]。BC群体的后代在表型上和其轮回亲本(recurrent parent, RP)类似,但携带一个或多个供体的目标性状。供体基因组被随机导入到 RP 背景中,并且 BC 后代中 RP 基因组的比例以[1-(1/2)(t+1)](t是回交的代数)的速率恢复,对一个特定的 BC群体而言,其携带的供体基因组数量可能存在很大差异[14]。传统的 BC 育种需要回交和筛选每一代 BC 的目标性状,其应用存在一定局限性,多用于提高优良品种单显性基因控制的抗病性等质量性状。例如用于提高优良水稻品种Pusa Basmati-1和Kao Dawk Mali 105的抗病性[15-16]。但BC育种作为一种相对保守的方法,较少用于多基因控制的数量性状的改良。这是因为携带显性单基因的BC植株可以在每一代BC中直接选择回交。而当目标性状受隐性基因控制时,育种者必需从每一代BC的BC_F1杂合植株中鉴定携带目标隐性等位基因的植株,而通过表型选择实现隐性基因的导入需要两倍于显性基因的时间。
使用随机BC分离群体进行数量性状位点(quantitative trait loci, QTL)定位可以得出两个主要结论[17-21]:第一,复杂性状一般受多个QTL控制,其效应受遗传背景影响;第二,主效QTL较少,大部分QTL为微效位点,容易受上位性和环境影响[22-24]。这两个结论为分子标记辅助回交(marker-assisted backcross, MABC)育种改良提供了便利[25]。一般来说,当有利基因和不利基因紧密连锁会导致遗传改良的失败,而MABC育种同时进行不利基因的背景选择和目标基因的前景选择时,效率会更高[26-27]。随着高通量测序技术的发展,基因型鉴定成本大幅降低,MABC被广泛应用于稻瘟病[28-29]、细菌性白叶枯病[30-32]、瘿蚊[33]、条纹病毒[34]和褐飞虱[35]等抗性改良,耐淹[36]、耐盐[37]、耐旱[38]、耐低磷[39]等非生物胁迫耐性改良,以及香味[40]等品质性状改良。
为了进一步将QTL挖掘与实际育种结合起来,Tanksley和Nelson提出了将QTL分析与品种选育结合起来的高代回交群体QTL分析方法(advanced backcross QTL, AB-QTL)[41]。该方法在进行QTL定位的同时,还可以将供体中的有利等位基因导入到优良自交系中,目前已广泛应用于水稻产量[42- 43]、品质[44]、生物和非生物胁迫[45-46]等复杂性状的遗传分析。但与典型的基因组作图研究一样,AB-QTL分析需要对随机群体进行基因型和表型的分型,因此很难应用于涉及大量育种群体的育种计划。为了克服AB-QTL方法的缺点和局限性,黎志康等[6-7]提出了BBSI的概念和策略(图1),用于复杂性状的遗传剖析和同步改良。
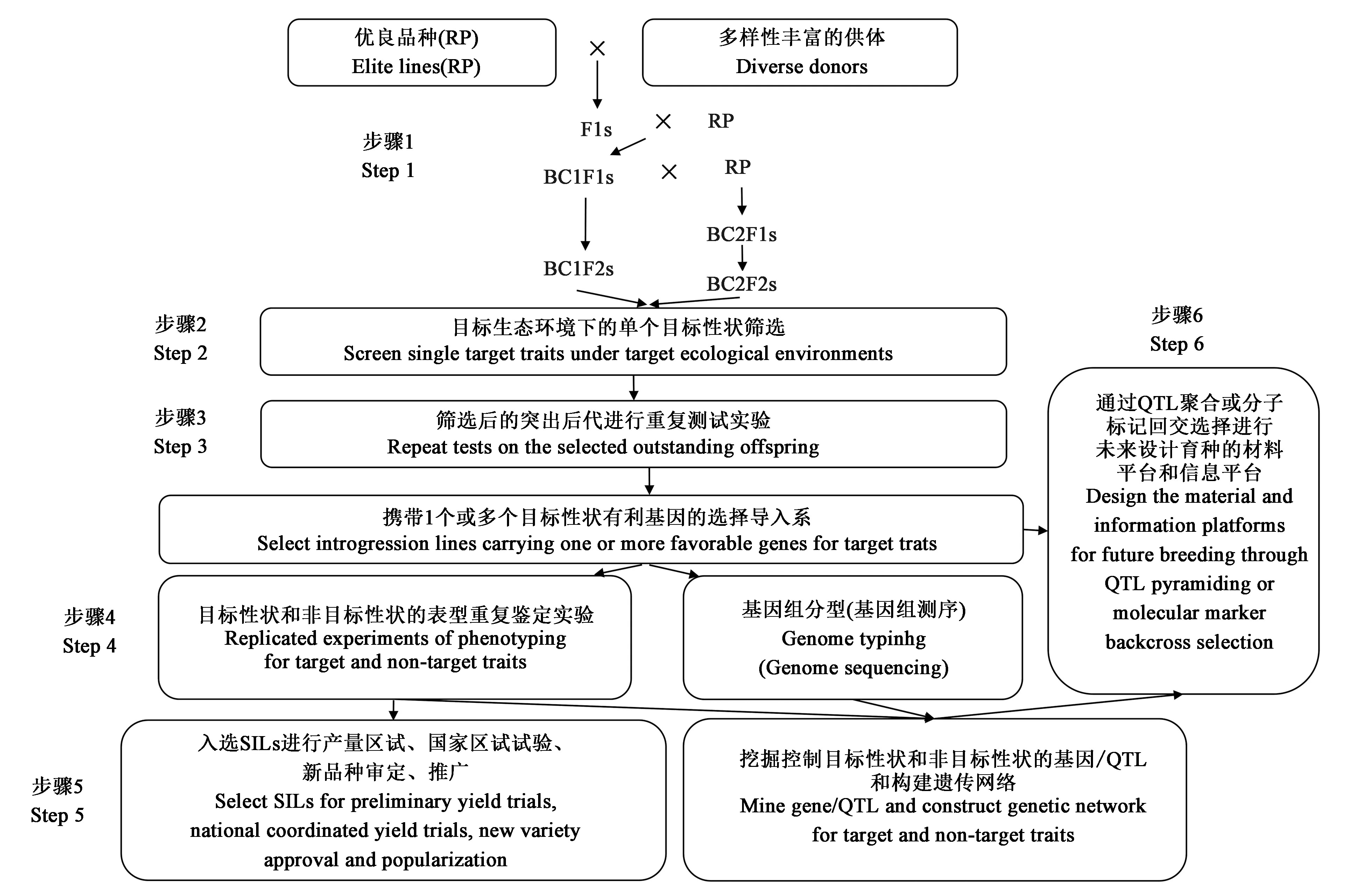
图1 选择导入育种策略
BBSI的整体策略主要为六个步骤:①利用一个优良品种作为RP与一组不同来源背景的供体进行杂交选育出BC1F2或BC2F2群体,进一步通过自交混收形成单一的混收BC1F2或BC2F2群体。②对每个BC1F2或BC2F2群体进行不同目标性状筛选,并与RP进行比较,筛选在目标性状上表现明显优于RP的单株(2%~5%)。③为单个(多个)目标性状选择的所有BC1F2或BC2F2植株在相同的筛选条件下对所有目标性状进行后代测试,所有入选单株在不同筛选条件下进行后代测试以选择第二个目标性状,并根据附加目标性状的数量和附加目标性状的剩余遗传变异量重复后代测试2~3次。到步骤③结束时,从单个 BC 群体中选择出的所有株系都被确认携带改良的目标性状,并在表型上与RP相似,形成一组在RP背景下的性状特异性的SILs。④使用分子标记分析选定的SILs,在目标环境中对目标性状进行精准表型分析,以挖掘影响目标性状的基因/QTL,以及通过连锁作图或选择导入(selective introgre-ssion, SI)等方法鉴定影响非目标性状的基因和QTL。⑤根据步骤④的结果,携带目标性状并在正常条件下优于RP的有希望纯合的SILs进入区试试验并逐步成为新品种,同时发掘目标/非目标性状基因/QTL及构建遗传网络。⑥上述优良RP遗传背景中选育的SILs及其与目标和非目标性状相关遗传(网络)信息,构成了通过设计聚合QTL(designed QTL pyramiding, DQP)或BBD进一步改良多个复杂性状的材料和信息平台。
2 选择导入育种策略的实践
2.1 BBSI在改良复杂性状上的显著作用
BBSI策略已在中国及菲律宾、越南、老挝、缅甸等多个国家实施,对提升当地优良品种对非生物/生物胁迫的耐受性或抗性以及正常条件下的高产等发挥了显著效果[47-54]。本文总结了BBSI的一部分结果,首先,几乎所有的复杂性状都存在大量有用的遗传变异,这在以往的水稻遗传改良中却未能得到充分利用,可能是由于这些变异在很大程度上被隐藏了。例如利用BBSI进行的耐冷性(cold tolerance,CT)[54]和耐淹性(submer-gence tolerance, SUT)[55]研究中,3种籼稻类型的供体都表现出较差的CT,但导入粳稻背景下的SIL却表现出更强的CT,说明这些供体虽然本身CT较差,但仍可用作改良目标品种耐冷性的优良供体;籼稻TKM9和Khazar两种供体对淹水高度敏感,但导入3个优良遗传背景的BC后代中都产生了更强的SUT。显然,耐冷和耐淹的基因/等位基因都隐藏在这些供体中。其次,BC育种加上适当的表型选择(在适当的压力下与 RP 直接比较)是有效发掘这种“隐蔽基因”的有利方式。最后,与RP关系较远的供体往往在BC后代中贡献更多的超亲分离(表1)[56-57]。
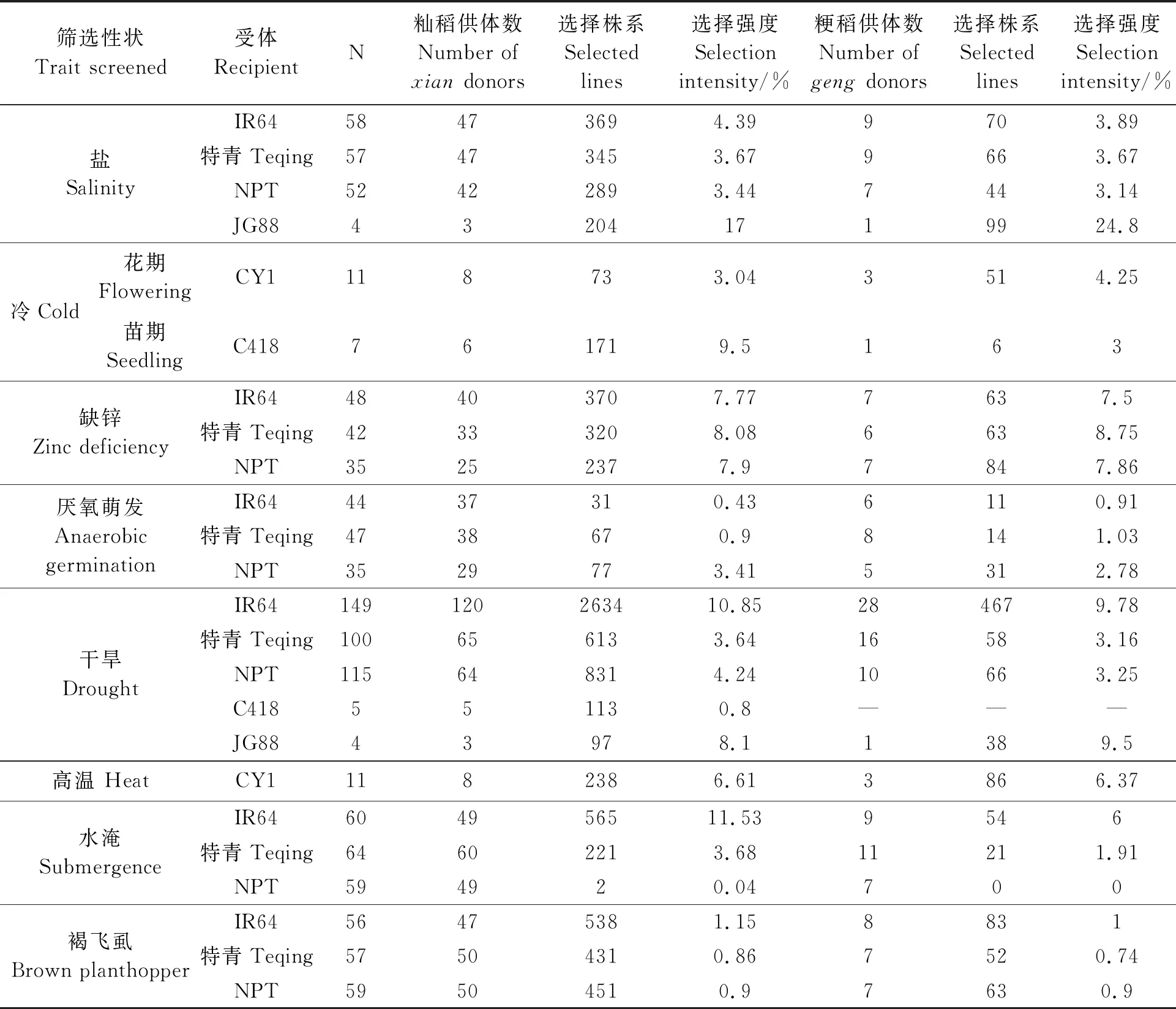
表1 利用BBSI改良8个生物及非生物胁迫性状[56-57]
BBSI育种不仅可以改善水稻单一性状,还可以通过对不同目标性状的多轮表型选择同时改善2~3个复杂性状。Ali等[58]以具有广适性的优异品种黄华占(Huanghuazhan, HHZ)为受体,8个不同来源的亲本(Phalguna、IR64、特青、OM1723、IR50、PSBRC66、CDR22、PSBRC28)为供体,构建8个BC1群体,通过3轮表型选择共获得了496个BC1F4HHZ_SILs。在1~3个非生物胁迫和非胁迫条件下,大部分SILs的产量得到显著提高。在496个BC1F5SILs中,至少有13个在短短的6年时间内被直接利用,成为东南亚、南亚等国家的绿色超级水稻(green super rice, GSR)主栽品种,推广种植超过100万hm2[9]。BBSI 步骤①~③和⑤中的BC育种部分已在中国和国际水稻研究所(IRRI)大规模实施,从200多份来自世界各地的微核心种质中,将有利的性状/基因/等位基因导入46个优良亲本(商业品种和杂交亲本)。通过BBSI的实施,这些品种在少水、少肥和少药的条件下仍表现出高产和稳产的特性[9]。上述研究都清楚地证明了BBSI作为一种改良复杂性状育种策略的强大力量和优势。这些SILs是未来BBD育种中揭示控制复杂性状背后的基因/QTL及等位基因的核心材料[12]。
2.2 BBSI对遗传学研究的理论贡献
目前,已经鉴定和克隆到多个影响农艺性状的QTL,但是大部分位点尚未克隆。影响复杂数量性状的非等位基因间互作,尤其是多个基因间普遍存在的高级互作是造成绝大多数QTL定位研究及解释困难的主要原因[59-62]。
通过整合基因信号转导的网络模型和传统数量/群体遗传的理论,Zhang等[63]提出了用于剖析复杂性状遗传网络的分子数量遗传学理论。该模型根据影响复杂性状的信号转导途径中两种功能基因之间最基本的功能依赖关系(functional dependency, FD)定义了两个概念:功能遗传单元(functional genetic units, FGUs)和层次原则(principle of hierarchy)。FGUs代表在每个信号通路水平上同一层次起作用的一组基因,它们之间的功能存在相互依赖的关系。基因间的两大类功能依赖关系(上下游功能依赖和同一个FGU内的功能依赖)决定了数量遗传和群体遗传理论模型中的QTL间互作(epistasis)和非随机关联(non-random associations)的存在(图2)。
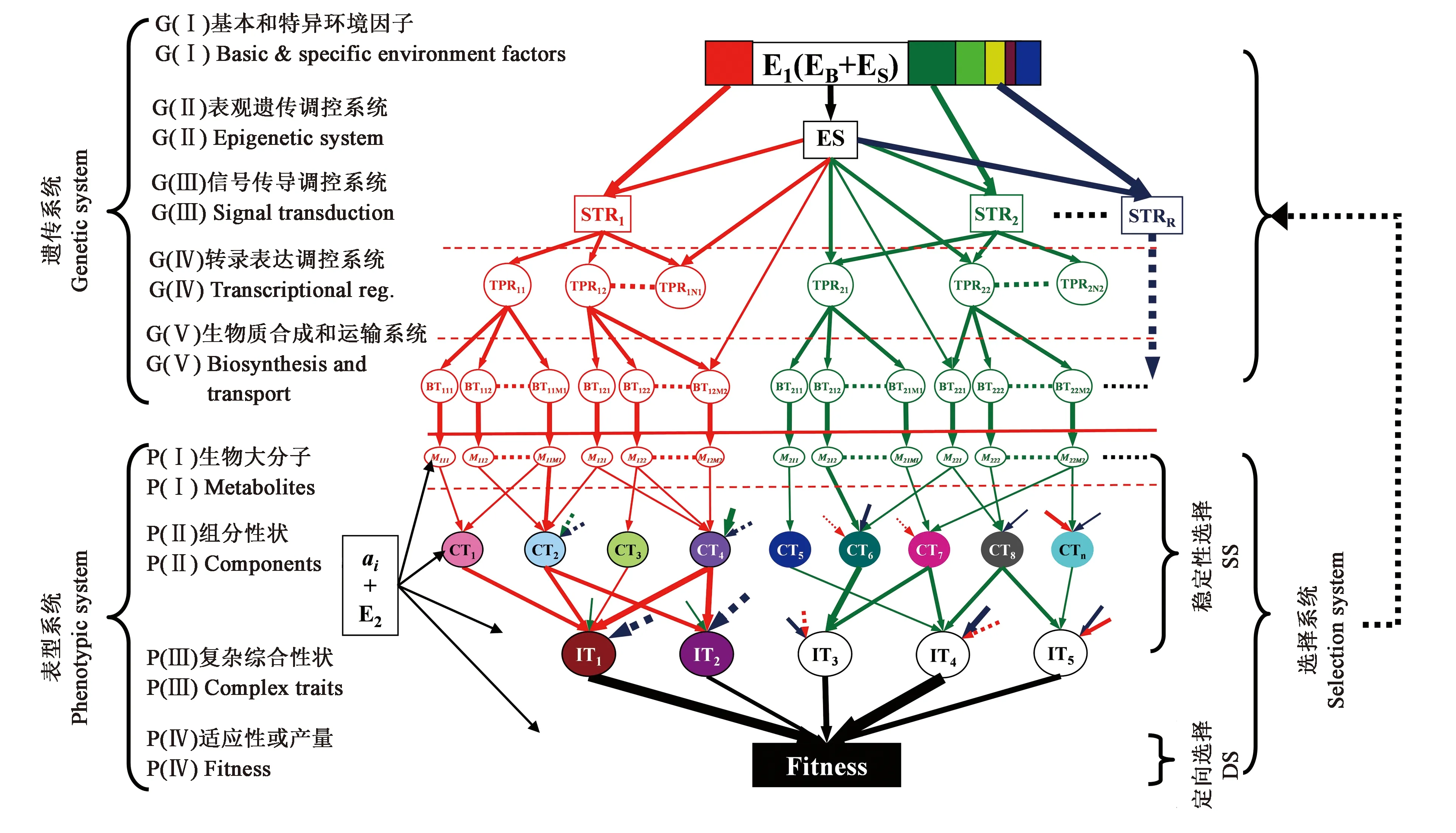
图2 复杂性状的基本遗传模型
基于选择导入群体的BBSI过程可以初步验证这一理论模型,步骤⑤是利用选定的SILs和DNA标记,揭示目标性状和非目标性状的基因/QTL和遗传网络,具体如下:从BC群体中选择出一套SILs来改良目标性状,这些性状必需与影响这些性状的供体等位基因相关联。因此,对于入选的SILs,其目标性状相关基因位点的供体等位基因出现的频率将高于未被选择的BC群体中理论上预期的频率。SILs中特定位点(QTL)上供体等位基因的频率越高,说明供体等位基因对目标性状的影响越大。对于来自一个特定BC群体的SILs,使用DNA标记检测控制目标性状的主效基因/QTL需要进行全基因组标记检测以确定供体等位基因频率超出其在未选择群体中的理论预期的位点。从统计学上说,控制目标性状的非连锁主效位点(存在上位性互作)选择可能导致供体等位基因之间的一种很强的非随机关联[64]。因此,通过对BC群体中的目标性状进行强胁迫选择选育出的SILs是检测控制目标复杂性状的上位QTL的有用材料[63]。
BBSI成功地利用SILs和上述方法在水稻中分析了多种非生物胁迫(干旱、水淹、低温)耐受性的遗传网络。例如,利用优质粳稻品种吉粳88(Jigeng 88, JG88)为RP,分别与四个供体杂交,从产生的BC2F2分离群体中选择了72个SILs用于干旱胁迫下的产量性状改良与遗传网络构建[65]。利用上述理论模型鉴定到一个由29个FGUs(包含30个QTLs)组成的假定遗传网络;利用来源于三个轮回亲本IR64、特青、NPT和三个供体TKM9、FR13A 和 Khazar 之间的杂交,从产生的9个BC群体中选择出162个耐淹SILs用于水淹胁迫下的遗传网络构建[55],共鉴定到12个遗传网络,每个遗传网络都包含多个控制耐淹性状的FGUs,共挖掘出由68个bin区间组成的167个FGUs,包含295个QTLs;利用相同策略,对于优良粳稻C418(RP)与4个冷敏感籼稻(供体)杂交形成的BC2F2群体,经两轮选择和后代测试,筛选出30个耐冷的SILs[53]。在水稻营养阶段,分析了强烈冷胁迫选择的全基因组响应,最终构建了一个由28个FGUs组成的遗传网络,包含56个QTLs,其中有4个显著超导入供体等位基因位点。又利用超优一号(Chaoyou 1, CY1)(RP)和5个遗传背景多样但冷敏感的供体(生殖阶段进行两轮CT强选择)的回交BC2群体,选择出48个CT_SILs,构建了一个由46个FGUs(包含50个QTLs)组成的遗传网络[66]。
与传统利用大量随机分离群体的QTL定位方法相比,BBSI策略在非生物胁迫耐受性的遗传分析方面具有四大优势:首先,BBSI在位点检测数量、等位基因挖掘和高阶上位性检测能力方面更为高效,利用BBSI总共鉴定出110多个控制非生物胁迫耐受性的QTL(在一个平均群体大小为14的SILs/群体中,平均可以检测到17.7个QTLs)[57]。其次,通过BBSI可以进一步揭示控制DT、SUT和CT性状的QTL遗传网络,以及对应DT、SUT 和 CT的SILs控制目标性状 QTL位点的图形基因型(图3)。这与对基因位点之间的遗传关系的认知是一致的,即植物非生物胁迫耐受性涉及许多信号通路和复杂的基因网络调控。第三,大量遗传网络的位点研究表明,亲本功能等位基因的遗传互补性和排斥性解释了水稻非生物胁迫耐受性的隐蔽多样性和超亲分离(表1)。最重要的是,所鉴定的影响目标性状的位点的SILs图形基因型为BBD提供了直接的信息,揭示了每个SILs的目标性状如何通过检测到的QTL组合实现目标性状改良。
3 后基因组时代BBSI的机遇和挑战
3.1 水稻基因组遗传多样性对设计育种的巨大潜在价值
近年来,以“3 000份水稻基因组研究”(3K-RG)为代表的水稻群体基因组研究取得了令人瞩目的进展,揭示了水稻种群中具有的丰富遗传多样性[11]。这些研究成果在未来育种上的应用潜力反映在两大方面:其一,水稻种内存在极为丰富的遗传变异。主要体现在:①3 010份水稻基因组中发现了超过29 M单核苷酸多态性(single nucleotide polymorphisms,SNPs)和超过25万个小插入缺失标记(small insertion and deletion poly-morphisms,Indels)[11];②在>35 000个水稻基因位点上存在超过255 000个功能等位基因[67];③>30 000个水稻基因的存在-缺失性变异(presence and absence variation, PAV)[11];④水稻品种基因组间存在大量微结构变异(缺失、易位、重复和倒位)。其二,水稻种内极大的遗传变异主要表现在不同地理来源种群间的遗传差异,如何充分利用种质资源中的有利遗传变异是未来育种取得突破进展的关键[67]。上述变异数据库的构建最终目的是能够充分利用这些信息实现设计育种,即利用基因功能及其多样性的精准信息按照人类的意愿设计、并高效地培育出符合人们各种需求并适应不同生态区域的优质、高产、多抗水稻新品种。毫无疑问,随着全球水稻功能基因组研究的飞速发展和相关大数据的快速累积,我国水稻育种正在进入基于基因组大数据的分子设计育种时代。
3.2 如何实现精准遗传变异信息下的复杂性状高效改良
为了将水稻功能基因组学中所获得的基础理论与实际应用紧密结合,我国科学家在实践BBD理念方面进行了积极地尝试。李家洋院士团队在解析理想株型基因IPA1介导的株型发育遗传调控网络基础上,创造性地提出了理想株型塑造与杂种优势利用相结合的BBD新思路,利用分子标记辅助选择等方法培育出适宜长江中下游稻区种植的“嘉优中科”系列水稻新品种,在籼-粳亚种间杂种优势利用与理想株型相结合方面获得了重要进展[67];此外,钱前院士团队通过优良目标基因辅助选择,以日本晴和93-11为供体,经过8年的杂交、回交和结合分子标记定向选择,对特青(超高产品质相对较差)进行了改良,获得了若干份优异后代材料[68]。最近,黄学辉团队构建了水稻数量性状基因关键变异(causative variation)图谱,有望为水稻新品种的快速培育提供技术支持[69]。上述研究是我国在水稻分子设计育种方面的重要突破,证实了水稻BBD的概念和前景,但如何克服基因间互作及其与环境互作对复杂农艺性状的不利影响,将水稻功能基因组和群体基因组的研究成果转化为对复杂农艺性状的精准和高效改良,仍是实现以改良复杂农艺性状为主要目标的设计育种面临的重大挑战和科学难题。
BBSI 的最后一步(图1)是利用从步骤③~⑤中获得的性状特异性SILs 及其目标性状和非目标性状的表型和遗传信息,通过DQP选育优良品种,解决基因间的互作问题,在揭示控制复杂性状的遗传信息的同时,实现多个复杂性状的改良。尽管离精准设计的目标还有一定距离,但DQP已逐渐成为未来实现 BBD 的重要策略,被广泛应用于提高水稻的高产和多种非生物胁迫耐受性[65, 70-71]。根据SILs的表型变异,结合检测到的控制目标/非目标性状的QTL/有利等位基因,通过MAS或表型选择,很容易实现“多供体来源+同一优良遗传背景”的多目标性状有利等位基因的聚合。
3.3 后基因组时代BBSI的最佳策略设想
目前,植物育种家面临的巨大挑战是育种目标的重大转变,从强调提高产量到充分利用种质资源以实现可持续的高产稳产。在全球气候变化导致的日益频繁和极端的环境威胁下,以及对粮食质量的多样化要求都增加了满足未来粮食需求的难度[12]。为了应对这一挑战,未来的BBD育种技术必需建立在作物功能基因组学和群体基因组学进展所获得的大量信息基础上,要达到BBSI的最佳效果,需要注意以下四点:首先,BBSI步骤①~③的主要目的是在优良遗传背景下选育成百上千的SILs,作为未来BBD的材料平台。因此,除了选择对目标环境具有广泛适应性的“最佳”品种作为RP外,使用200~300个来自世界种质资源中的微核心种质作为供体材料是确保每套优良背景的SILs都包含大量基因位点上的“最佳”等位基因的关键,这些基因位点是在目标环境下鉴定的,控制着来自供体基因库导入的任何有益目标性状。在进一步BBD过程中,该策略将大大提高供体等位基因的功能(表型)可预测性。其次,需要针对目标性状进行有效的表型选择。在第一轮单株选择中,BC的初始分离群体的大小应在400~600之间,以便在关键发育阶段施加严重(对RP几乎致命的)胁迫来鉴定优良BC后代(直接与RP比较)。这将提高选择的准确性,在早期的BC世代,选择株系的数量减少到一个可管理的大小。然而,作为加速育种过程的一种常见做法,冬季环境是否能够有效地选择目标性状的表型以及如何选择表型目前尚不清楚。正季和冬季环境的差异如何影响不同目标性状的基因或QTL表达和选择效率是未来需要研究的一个问题。第三,除了具有较高的改良性状能力和效率之外,BBSI与传统的基因/QTL定位方法相比最大的优势是目标和非目标性状的所有遗传信息(数量、基因组位置以及基因和等位基因的遗传关系)可以直接适用于实际育种群体,因为所有这些都是在目标遗传背景和环境中获得的。与在中国和IRRI成功实施 BBSI 的BC育种部分相比[47-53],在中国实施大规模的水稻BC育种工作背景下,全面实施BBSI第⑤步进行大规模基因或QTL挖掘以及目标和非目标性状的等位基因挖掘一直是一项重大挑战。幸运的是,随着基因分型的成本快速降低以及水稻群体中各种类型基因组多样性的公开可用性[11],预计BBSI策略的这一部分将得到极大的促进和加速[67, 72]。
4 展望
综前所述,未来两项重大进展标志着中国水稻育种已准备好进入后基因组时代的分子设计育种。第一,中国多个机构已经选育了30多个优良遗传背景的数千个性状特异性SILs,这些SILs包含来自一个微核心种质的各种目标性状的大量有益遗传多样性(未发表数据);第二,全球主要作物功能和群体基因组研究的进展最终将揭示水稻基因库中的所有功能变异,这些变异已经或正在被整合到各种数据库中。此外,利用选择的SILs和全基因组选择响应的标记辅助可以很容易地扩展到高效的基因和QTL挖掘,以及利用传统系谱育种方法选育的育种群体的复杂性状等位基因挖掘中。后基因组时代下的BBSI育种策略将更加完善和精准,为未来农作物育种创新发展提供重要理论和实践参考。
