生境对药食两用植物接骨草的生长、叶绿素含量与叶绿素荧光特性的影响
2021-11-13陈兰英许航徐正刚肖娟
陈兰英, 许航, 徐正刚, 肖娟 , *
生境对药食两用植物接骨草的生长、叶绿素含量与叶绿素荧光特性的影响
陈兰英1, 许航2, 徐正刚2, 肖娟2, *
1. 西华师范大学, 生命科学学院, 南充 637000 2. 西华师范大学, 环境科学与工程学院, 南充 637000
探讨接骨草对不同生境适应能力和响应机制, 以川东北丘陵地区野外自然分布的药食两用植物接骨草(Lind.)为研究对象, 研究林内(生境Ⅰ)、林窗(生境Ⅱ)、林缘(生境Ⅲ)不同生境下接骨草植株高度、叶片形态、叶绿素含量和叶绿素荧光特性等的影响, 结果表明: (1) 不同生境接骨草叶片的植株高度、叶片长、叶面积具有显著的差异性, 林窗生境的植株最高, 叶片长、叶面积最大, 且与相对光强呈显著负相关, 与湿度呈显著正相关。(2) 叶绿素含量具有显著的差异性, 与植株形态的生长具有一致性, 均是林窗生境显著高于林内和林缘生境。(3) 不同生境叶绿素荧光参数差异显著, 林窗生境的接骨草实际光化学效率(ΦPSII)值较高, 而非化学猝灭系数()较低, 同时较高的表观电子传递速率(), 表明接骨草有较高光能利用能力和光呼吸能力。接骨草兼有阴生和向阳的特性, 在林窗生境下接骨草的长势、叶绿素含量和光能利用能力均优于林内和林缘生境, 接骨草的植株形态、光合生理特征是对不同生境适应能力的主导因素。在引种栽培接骨草时, 林窗的相对光照(56.05±0.78%)和相对湿度(52.78±0.63%)可作为遮光种植和田间管理的参照。
接骨草; 植株高度; 叶片性状; 叶绿素含量; 叶绿素荧光特性
0 前言
接骨草(Lind.)为忍冬科接骨木属(Lind.)的草本或亚灌木, 因茎有八棱又称八棱麻[1]。目前, 该种自然居群分布广, 几乎遍布北半球温带和亚热带地区, 在我国主要分布在秦岭以南, 包括陕西、广西、湖南、四川、贵州、浙江、江西、福建、台湾等省区[2]。接骨草作为传统中药, 具有祛风利湿, 舒筋活血的功效, 可有效治疗跌打损伤、风湿痹痛、筋骨疼痛及活血、行瘀、止痛等病症[3]。除此之外, 接骨草还含有许多人体必需的微量元素、氨基酸和其他营养成分, 具有很高的营养保健价值, 可作保健食品的开发[4]。同时, 接骨草花序顶生聚伞圆锥花序, 外形美观, 可用于观赏; 还具有较强的抗环境污染能力, 在污染较严重的工矿区周围也能良好生长[5-7]。因此, 接骨草具有极高的经济开发价值和广阔生态应用前景。目前对接骨草的研究多集中于药用化学成分、药理活性和临床应用上的研究, 光合生理方面仅见董磊磊等[5]和王琼等[8]在特定环境和短期时间下对接骨草进行光合-光响应和光合-CO2响应研究探讨接骨草的光合特性, 而对接骨草的野外生存环境, 特别是连续时间下生境异质性对接骨草的生长、叶绿素荧光特性及叶绿素含量的影响研究还未见有相关的报道。前期的野外调查发现, 接骨草主要分布林内、沟旁、山坡杂草丛中, 不同的生境因子对其生长和生理特性可能产生不同的影响。植物的叶绿素含量、叶片形态会影响叶片光合效率, 同时又受到光照强度、气温、空气相对湿度、土壤含水量等外界因素影响, 故揭示不同生境下接骨草的光合生理特性机制对深入了解接骨草的生态分布和移植栽培非常重要[9]。同时, 通过对接骨草生长特性的长期监测利用变异系数对不同生境接骨草的植株高度、叶片生长形态、叶绿素含量及叶绿素荧光特性的变异性进行评价是挖掘其有利基因并加以综合利用的有效途径, 也是种质资源研究最直接和最基本的方法[10]。因此, 本文在2017—2019年对3种不同的生境(林内、林窗、林缘)接骨草的叶片生长状态、叶片叶绿素含量和叶绿素荧光特性进行测定, 并比较不同生境接骨草的差异并分析其变异性; 探讨生境异质性和环境因子对接骨草生理生长的影响, 为今后深入研究接骨草精细化栽培及种植技术、提高接骨草的生态适应性提供理论基础。
1 材料与方法
1.1 研究地及接骨草生境概况
实验于2017—2019年5月在南充市西山风景区进行(30°48'54'' N, 106°03'47''), 该区域属亚热带湿润型季风气候, 年平均气温17.4 带, 年平均降雨量1021 mm, 该地植物的生长期较长, 年无霜期约300 d, 土壤为红棕紫泥。在林缘、林窗和林内生境下分别选取有接骨草生长的20 m×20 m的固定样方, 3个样地的环境因子见表1, 3个样地中相对光强差异呈极显著(<0.01), 相对湿度具有显著差异(< 0.05)。其中, 相对光强为林缘>林窗>林内, 相对湿度为林内>林缘>林窗。

表1 接骨草不同生境主要环境因子概况
注: Ⅰ, 林内; Ⅱ, 林窗; Ⅲ, 林缘。同列数据凡具有不同小写字母表示在<0.05或<0.01水平上差异显著, 下同。
1.2 植物生长情况及叶片叶绿素含量测定
在2017—2019年5月1日—10日选择林缘、林窗、林内样方下2 a生的接骨草(生长良好、生长基本一致)各10株进行标记, 对接骨草进行植株的高度、植株中部叶片长、宽生长情况测量, 叶面积用LI-3000C便携式叶面积仪测定, 连续3 a测定的植株高度、叶片生长情况尽量保持同一植株、同一叶片。另在林缘、林窗、林内样方中随机标记10株接骨草植株, 每年从标记的10株接骨草植株中采摘鲜叶各300 g, 带回实验室进行叶绿素含量的测定, 叶绿素含量采用丙酮乙醇浸提法测定[11]。
1.3 叶片叶绿素荧光参数的测定
2017—2019年5月1日—10日选择晴朗的3 d, 使用LI-6400便携式光合仪测定叶绿素荧光特征参数。2017年每个生境随机选5株, 每株测4—5片叶, 在2018年和2019年尽量选择同株同片叶片进行测量, 3个生境共选择15株植株。测定前将待测叶片进行暗适应20 min, 然后用弱光(0.1 μmol·m–2·s–1)照射, 测定初始荧光(F), 再照射饱和脉冲光(3000 μmol·m–2·s–1)2s, 测定暗适应最大荧光(F)。通过计算得到: PSII最大光化学效率F/F= (F–F)/F, 光合效率潜能F/F= (F–F)/F。在叶片充分光适应30 min后测定稳态荧光(), 最后照射饱和脉冲光(3000 μmol·m–2·s–1)0.8 s, 测出光适应最大荧光(F’)和表观光合电子传递速率()。通过计算得到PSII光合量子产额PSII= (F’ –)/F’, 表观电子传递速率= 0.58×φPSⅡ××0.5, 光化学淬灭系数()= (F’ –)/(F’–F’), 非光化学淬灭系数()= (F–F’)/F’[11]。

1.4 数据分析
采用SPSS 20.0统计分析软件进行数据分析, 双因素方差分析(Two-way ANOVA)用于检验不同年份和不同生境交互作用对接骨草生长、叶绿素含量和叶绿素荧光特性的影响; 同一时间不同生境之间的指标比较采用单因素方差分析(One-way ANOVA), 并采用LSD法进行多重比较, 采用相关分析和曲线拟合方法揭示环境差异因子对接骨草植株生长的影响, 使用Excel软件作图。
2 结果与分析
2.1 不同年份和不同生境下接骨草植株高度、叶片生长的动态变化差异及变异系数
3种生境下接骨草的植株高度、叶片生长均存在明显的年份动态变化(图1, 表2)。随着年份的增长, 接骨草的植株高度、叶片长、宽和叶面积均增大。从重复测量方差分析结果来看, 不同生境对植株高度、叶片长、叶面积具有显著的影响(<0.05), 而对叶片宽无显著的影响(0.05), 不同年份对接骨草植株高度、叶片生长均有显著影响(<0.05), 而生境和年份的交互作用对接骨草的植株高度、叶片生长无显著影响(0.05)。
3种生境接骨草的植株高度随着年份的增长均呈线性增长, 在叶片生长动态中, 2017—2019年植株高度、叶片长及叶面积随着时间的增长不断增加(图1, 表2)。在2018—2019年, 林窗生境Ⅱ植株高度增长显著大于林内生境Ⅰ和林缘生境Ⅲ(< 0.01), 均是林窗>林内>林缘。在叶片长中, 生境Ⅱ显著大于林内生境Ⅰ和林缘生境Ⅲ(<0.05), 生境Ⅰ和生境Ⅲ无显著差异(>0.05); 在3个生境下, 叶片宽度生长情况无明显区别; 叶面积在不同生境和不同年份均具有不同程度的差异(<0.05), 生境Ⅱ显著大于其他两个生境, 叶面积的增长基本上与叶片长呈一致性的规律。整体上, 3个生境的接骨草植株高度, 叶片长、宽和叶面积的变异系数均较低, 植株高度的变异系数变化幅度为1.376%—5.375%, 叶片长的变化幅度为1.005%—6.598%, 叶片宽的变化幅度为3.009%—12.872%, 叶面积的变化幅度为1.500%—7.966%, 这说明3个生境的接骨草的植株高度、叶片长和叶面积的表型性状遗传稳定性较高。
植物地上部分的生长会影响光能的获取和水分利用, 同时植物通过对形态特征的适应调整去适应环境的变化, 从而获得更多有效的资源[12-14]。3个样地中相对光强和相对湿度具有显著差异(表1), 为进一步表述不同生境中环境差异因子对接骨草表观形态的影响, 对接骨草的植株高度, 叶片生长性状和相对光强和相对湿度进行线性拟合。如图2可见, 株高、叶长和叶面积与相对光强呈显著负相关(p< 0.05), 叶宽与相对光强无显著相关性(p>0.05); 株高、叶长、叶宽和叶面积与相对湿度均呈显著正相关(<0.05), 这说明3个生境中具有显著差异的环境因子湿度、光照条件影响接骨草的生长, 与相对光强的负相关和相对湿度正相关也进一步论证了接骨草属喜阴性植物, 在其他自然条件无显著差异的条件下, 光照强度越高越不利于接骨草的生长, 而湿度越大越利于接骨草的生长。

图1 不同生境接骨草植株高度、叶片生长动态差异及变异系数
Figure 1 Comparison of plant height, leaf growth dynamics and variation coefficient ofin different habitats

表2 植株高度、叶片生长对不同生境和年份的双因素方差分析
注: *表示差异显著(<0.05), **表示差异极显著(<0.01); 下同。
2.2 接骨草叶绿素含量的动态差异及变异性
3种生境下接骨草的叶绿素含量均具有显著差异(表3)。不同年份、生境和年份的交互作用对接骨草叶绿素含量无显著影响(0.05)。
叶绿素是维持植物光合作用的主要因子, 其含量和比例的变化是植物适应和利用环境因子的重要指标, 也是植物对环境胁迫的适应反应。由表3可知, 2017—2019年同一生境的叶绿素a()、叶绿素b()、总叶绿素(总)含量和叶绿素()无显著差异, 但不同生境差异显著(<0.05), 这表明不同的生境中的接骨草在吸收和利用光能方面差异较大(表4)。由表4可知, 在中, 生境Ⅱ显著大于生境Ⅰ和生境Ⅲ, 生境Ⅰ和生境Ⅲ间无显著差异;和总中, 均是生境Ⅱ>生境Ⅰ>生境Ⅲ(0.01); 在中, 生境Ⅱ>生境Ⅰ和生境Ⅲ(<0.05), 生境Ⅰ和生境Ⅲ无显著差异。变异系数中, 生境Ⅱ的变异系数均低于9%, 说明该生境植株叶片的叶绿素含量的变异性较低。

图2 接骨草植株高度、叶片生长特征与环境差异因子相关性分析
Figure 2 Correlation betweenplant height, leaf growth characteristics and environmental difference factors

表3 叶绿素含量对不同生境和年份的双因素方差分析

表4 接骨草不同生境叶绿素含量的差异及其变异性
2.3 不同生境接骨草叶绿素荧光参数的差异及变异性
通过对叶绿素荧光参数的测定, 可获得植物光能利用途径的信息[15-16]。重复测量方差分析的结果表明(表5), 3个生境的叶绿素荧光参数均达到了显著的水平(<0.05或<0.01), 这表明接骨草在不同生境下对光能的利用有着显著的差异。不同年份、生境和年份的交互作用对接骨草叶绿素荧光参数无显著影响(0.05)。

表5 叶绿素荧光特性对不同生境和年份的双因素方差分析
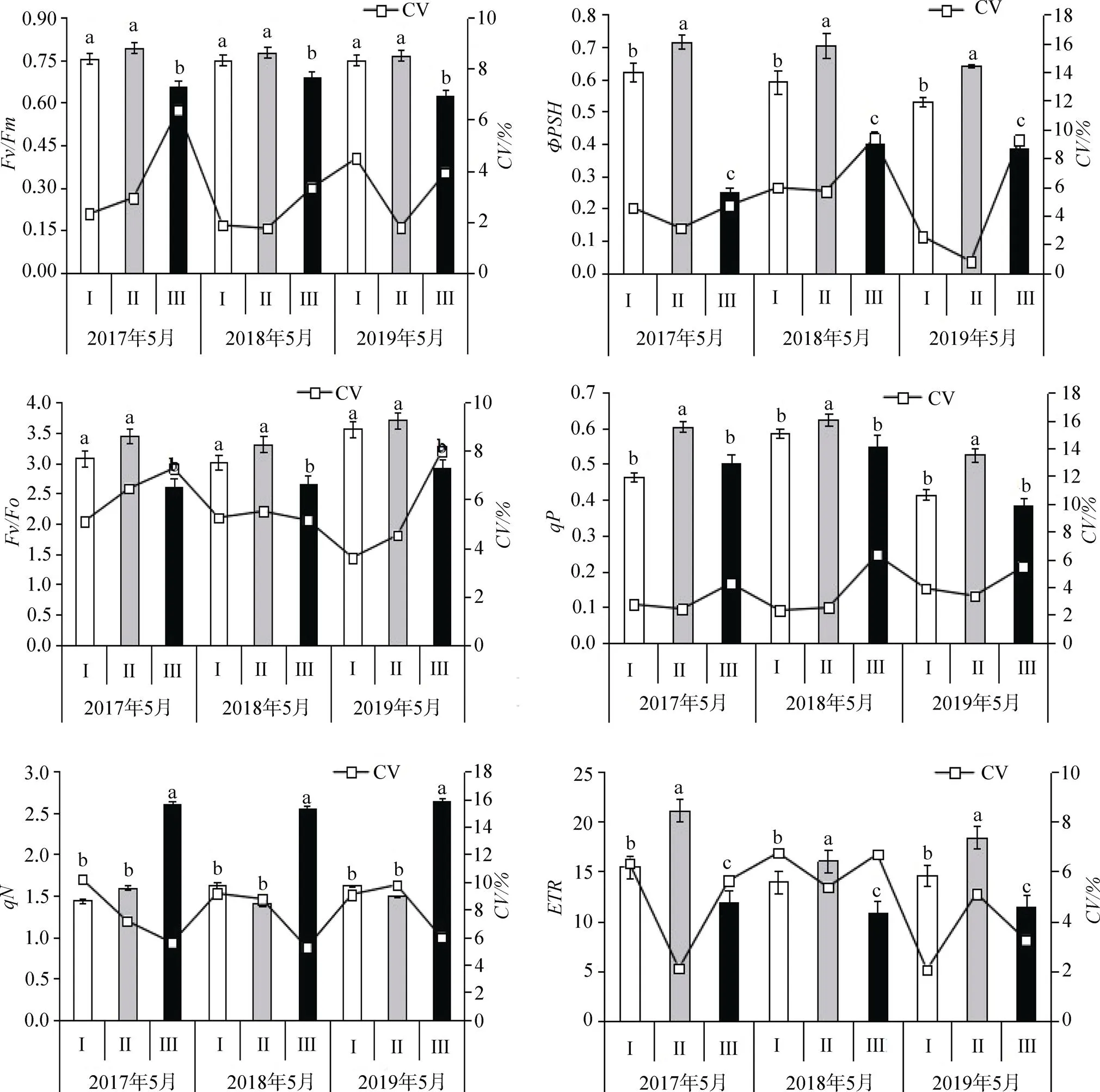
注: Fv/Fm: PSⅡ最大光化学效率, ΦPSa: 实际光化学效率, Fv/Fo: PSⅡ的潜在活性, qP: 光化学淬灭系数, qN: 非光化学淬灭系数, ETR: 表观电子传递速率。
Figure 3 Differences in chlorophyll fluorescence parameters ofin different habitats
/反映是PSII反应中心最大光能转换效率的反映指标, 反映PS中反应中心内禀光能转换效率, 也是表征植物是否受到长期光胁迫的一个有效指标[17]。在不受光抑制的情况下,v/m一般介于0.75—0.85之间, 当值低于0.75, 越低则表明植物受光抑制的程度越高, 不利于植株的生长。由图1可知, 接骨草林缘生境Ⅲ的v/m小于0.75, 在0.627—0.693范围内, 说明该生境下的接骨草植株的生长受到光抑制, 生境Ⅰ和生境Ⅲ的v/m值无显著差异, 且值均在0.75—0.85之间, 这说明在这两个生境下生长的接骨草植株PSII 系统光能转换效率正常。v/m在非胁迫条件下数值变化极小, 不受物种和生长条件的影响, 而在胁迫条件下明显下降[18]。3个生境的在2017—2019年测定值中, CV的变化范围为1.736%—6.373%, 这也进一步说明不同生境中接骨草v /m遗传变异性较小, 接骨草长期生长在强光条件中会导致PSII 反应中心失活或受到破坏, 进而导致接骨草的光合作用受到光抑制。
反映植物实际的光合效率, 从分析结果可知, 不同生境接骨草的值具有极显著差异(<0.01), 其中, 生境Ⅱ其生境Ⅰ境生境Ⅲ。在生境Ⅱ中值为0.640—0.715, 与v/m值相差约0.07—0.1; 在生境Ⅰ值为0.532—0.621, 与v/m值相差约0.135—0.219; 生境Ⅲ值为0.255—0.401, 与v/m值相差约0.292—0.404; 说明接骨草生境Ⅱ叶片所捕获的光能转化能植株所需的化学能的转化效率高于生境Ⅰ和生境Ⅲ, 生境Ⅲ的转化效率最低。
v/o表示PSII植物叶片潜在的光化学活性,v/m和v/o被绝大多数研究者认为是反映叶片光合效率的重要依据, 两者的值越高说明植物叶片所捕获的光能可以更加有效地转化为植物所需要的能量[19]。不同生境接骨草的v/o值具有极显著差异(<0.01), 与v/m的变化趋势一致, 生境Ⅱ和生境Ⅰ无显著差异, 生境Ⅲ最低。
反映天线色素吸收的光能用于光化学电子传递比例, 反映PSII开放程度及原初电子受体(Q)的还原情况。研究结果表明, 生境Ⅱ的值显著大于生境Ⅰ和生境Ⅲ(<0.05), 生境Ⅰ和生境Ⅲ间无显著差异, 这说明生境Ⅱ利用光能效果较生境Ⅰ和生境Ⅲ高。
反映了植物热耗散的能力, 植物叶片出现非光化学淬灭是对光合机构保护的机制, 不同生境的接骨草的热耗散能力差异较显著(<0.05), 其中生境Ⅲ的值较高, 表明具有较强的热耗散能力, 说明生境Ⅲ的接骨草在光环境下PSII天线色素吸收的光能用于热耗散的比例显著高于生境Ⅰ和生境Ⅱ光环境下的对应部分, 而用于光合的比例对应的会低于生境Ⅰ和生境Ⅱ光环境下的部分, 这可能是由于生境Ⅲ的接骨草生长状态不如生境Ⅱ和生境Ⅰ的重要原因之一。
表观光合电子传递效率值反映了实际光强下的表观电子传递速率,值越高说明植株能将有效量子传递到光反应的能力越强。不同生境的接骨草值具有显著的差异(<0.05), 生境Ⅱ>生境Ⅰ>生境Ⅲ。
3 讨论与结论
植物在不同的生境条件下的形态特征的变化情况是反映植物对环境适应能力的重要指标, 叶片作为营养器官同时也是植物光合作用的主要器官, 为适应异质性生境常表现出较大的可塑性, 其形态及光合生理特征被认为能很好地体现对异质生境的适应[20]。通过对不同生境的接骨草的株高增量, 叶片长、宽和叶面积增量的比较后发现, 随着年份的增加接骨草的株高、叶片性状随之增长。不对同生境同一测定年份进行单因素方差分析表明, 生境Ⅱ更利于接骨草的生长, 株高、叶片长、叶面积均显著高于生境Ⅰ和生境Ⅲ。相关性分析表明, 植株高度、叶片长和叶面积均与相对光照呈显著负相关, 株高、叶片长和宽、叶面积均与相对湿度呈显著正相关(图2); 这说明光强越大则不利接骨草的生长, 湿度越大越利于接骨草的生长。前人对接骨草的光合-光响应曲线及光合日变化进行研究发现, 接骨草是一种喜阴性的草本植物, 对弱光利用能力较强[5,8]。这与本文的研究结果一致, 接骨草在相对光强较高的林缘生境, 其株高和叶片生长特性均最小。在本文的研究中接骨草在林窗生境中株高、叶片长和叶面积最大, 这也进一步说明接骨草耐阴, 同时也具有向阳生长的特征, 对光的适应能力比较强, 分布广泛。黄雅等人的研究结果表明不同产地的接骨草主要药用成分总多酚依次为叶>全草>茎>根[21], 在本研究中林窗生境中植株较高和叶片较大说明接骨草在高度和广度两方面都增加了对资源的获取和利用, 植物在环境压力或资源有限条件下将分配更多的生物量到叶, 这样能更有效地捕捉光能, 有利于接骨草次生代谢产物的合成及积累。同时, 3个生境的接骨草的株高、叶片生长性状的变异系数较低, 表明其有较稳定的遗传性, 生境Ⅱ的接骨草生长更好, 可从生境Ⅱ中进行引种栽培。
光合作用是植物生物产量的基础, 叶绿素是植物光合作用必不可少的光催化剂, 其含量和比值常作为植物适应环境的重要评价因子, 它可以将捕获的光能转化为化学能, 在植物的光合作用过程中起着重要的作用, 叶绿素含量的变化与植物的生长发育有着密切的关系[22]。本研究中, 林窗生境下的接骨草生长较好, 其叶绿素含量也相对较高, 随着光照强度的减弱, 接骨草的叶绿素a, 叶绿素b, 总叶绿素含量增加, 表明光强的降低可促使植株更好的吸收光能, 适度的林窗光照可以降低强光对叶绿素的破坏, 从而使接骨草叶片更加有效的进行光合作用, 这与蒋运生的研究规律一致[23]。高温、强光和干旱胁迫会影响叶绿素的合成, 促进叶绿素的分解, 使其含量下降[24]。本研究中接骨草不同生境叶绿素相对含量具有显著的差异(表3), 林窗生境Ⅱ的变异系数的范围低于9%, 叶绿素含量较高的植株变异系数较低。光照强度在一定范围内, 接骨草的叶绿素含量能够达到最大值, 光照过弱或过强均会使叶绿素含量呈下降的趋势, 林窗生境Ⅱ的光照范围能够使其达到最大值。
叶绿素荧光与光合作用反应过程紧密相关, 叶绿素荧光技术是一种快速、灵敏、无损伤的技术, 是光合作用的探针, 任何环境因素对光合作用的影响都可以通过叶绿素荧光反映出来[25-26]。反映PSII反映中心利用所捕获激发能的情况, 被认为是植物发生光抑制最明显的特征[27]。在3个生境中, 林缘生境的值低于0.75, 说明其生境的接骨草在生长过程中受到了光抑制。光合量子产量是用于衡量植物光合电子传递的量子产量, 可作为植物叶片光合电子传递速率快慢的相对指标, 在本研究中不同生境的实际的光合效率、潜在的光化学活性、光化学淬灭系数均表现出显著的差异性, 表明不同的生境的植株对环境的适应能力不同, 这与张其德和任士福等人对相同植物不同生境的叶绿素荧光特性的研究结果一致[28-29]。在众多荧光参数中, 高的v/m、v/o和PSII值已基本被认为是叶片高光合效率的重要依据, 且不少研究指出v/m、v/o和PSII有很好的一致性[30], 这与本研究的结果一致。大多数研究者认为, 非化学猝灭系数低的植物光合性能较好[31]。本研究中, 林窗生境和林内生境的接骨草PSII值较高, 非化学猝灭系数较低, 林缘生境的非化学猝灭系数较高, 这说明林缘生境的接骨草吸收的光能主要用于热耗散; 因此, 其实际的光化学效率相对较低。本研究中, 生境Ⅱ林窗生境的v/m、v/o和PSII值较高,值较低, 其生境更利于接骨草的生长。
林窗、林内及林缘的水、热、光等环境因子存在着显著差异, 致使生长在不同光环境中的植物光合生理特征各不相同, 从而影响接骨草的生长。本研究中不同生境的接骨草植株生长高度、叶片生长特性、叶绿素含量和叶绿素荧光参数具有显著差异。在林窗生境中, 接骨草的生长情况较好, 叶片的光合作用能力强, 对光适应生态幅较宽既耐阴也有向阳生长的特点, 接骨草在林窗的环境中竞争能力较强, 具有较高的潜在光能利用能力, 具有较好的生物学特性和生态适应能力。在林内的条件下表现也较好, 能正常生长, 但光合能力不如林窗生境。结合3种环境条件下接骨草的叶片叶绿素含量及光合生理特征, 可知在林窗条件下生长的接骨草光合作用能力更强、长势更好。在引种栽培接骨草时, 应模拟野外生境Ⅱ的生长条件, 提供适宜的光照条件和充足的水分。在实际的栽培种林隙光照条件(56.05±0.78%)和相对湿度条件(52.78±0.63%)可作为遮光种植和田间管理的参照。
[1] 李胜华, 李爱民, 伍贤进. 接骨草化学成分研究[J]. 中草药, 2011, 42(8): 55–57.
[2] 黄双全, 王孝民. 食果鸟白头鹎与接骨草的分布[J]. 植物学报, 2000, 42(10): 1096–1100.
[3] 姚元枝, 伍贤进, 黎晓英, 等. 接骨草的化学成分与药理活性研究进展[J]. 中成药, 2015, 37(12): 160–166.
[4] 赵湘婷. 中药接骨草化学成分研究[D]. 兰州: 兰州理工大学, 2014.
[5] 董磊磊, 黎云祥, 权秋梅, 等. 中药接骨草光合生理特性研究[J]. 草地学报, 2013, 21(4): 816–820.
[6] 陈志澄, 毋福海, 黄丽玫, 等. 砷矿尾砂污染及其治理研究[J]. 环境污染与防治, 2006, 28(3): 180–182.
[7] 班芷桦, 王琼. 喜旱莲子草和接骨草竞争对模拟增温的响应[J]. 植物生态学报, 2015, 39(1): 43–51.
[8] 王琼. 不同生境中入侵植物喜旱莲子草与本地种接骨草的光合特性[J]. 西华师范大学学报: 自然科学版, 2015, 36(3): 253.
[9] 李月灵, 金则新, 王强, 等. 不同生境华东野核桃光合生理特性及叶绿素荧光参数比较[J]. 浙江大学学报(理学版), 2013, 40(2): 221–229.
[10] 张秦徽, 王洪武, 姜国云, 等. 红松半同胞家系变异分析及选择研究[J]. 植物研究, 2019, 39(4): 557–567.
[11] 王琼. 喜旱莲子草和接骨草光合色素含量测定方法研究[J]. 西华师范大学学报(自然科学版), 2009, 30(4): 18–22.
[12] HUGHES A P. Plant growth and the aerial environment : vi. the apparent efficiency of conversion of light energy of different spectral compositions by impatiens parvielora[J].Phytologist, 2006, 64(1), 48–54.
[13] ORLANDO G, EVA R, YAJAIRA G, et al. Influence of low light intensity on growth and biomass allocation, leaf photosynthesis and canopy radiation interception and use in two forage species of(DC. ) Benth[J]. Grass and Forage Science, 2018, 73(4): 967–978.
[14] BAGOES M I, OKA P, RAHMA H, et al. Factors affecting biomass growth and production of essential oil from leaf and flower of[J]. Journal of Essential Oil Bearing Plants, 2018, 21(4): 1021–1029.
[15] 李鹏民, 高辉远, RETO J S. 快速叶绿素荧光诱导动力学分析在光合作用研究中的应用[J]. 植物生理与分子生物学学报, 2005, 31(6), 559–566.
[16] 李佳, 江洪, 余树全, 等. 模拟酸雨胁迫对青冈幼苗光合特性和叶绿素荧光参数的影响[J]. 应用生态学报, 2009, 20(9): 2092–2096.
[17] 李孟洋, 巢建国, 谷巍, 等. 高温胁迫对不同产地茅苍术开花前叶片叶绿素荧光特征的影响[J]. 植物生理学报, 2015, 51(11): 1861–1866.
[18] 徐德聪, 吕芳德, 栗彬, 等. 不同品种美国山核桃叶绿素荧光特性的比较[J]. 果树学报, 2008, 25(5), 671–676.
[19] 张守仁. 叶绿素荧光动力学参数的意义及讨论[J]. 植物学报, 1999, 16(4): 444–448.
[20] 吴涛, 耿云芬, 柴勇, 等. 三叶爬山虎叶片解剖结构和光合生理特性对3种生境的响应[J]. 生态环境学报. 2014, 23(10): 1586–1592.
[21] 黄雅, 陈国华, 周欣, 等. 黔产接骨草中总多酚的含量测定及抗氧化活性研究[J]. 天然产物研究与开发, 2017, 29(2): 255–263.
[22] AND G H K, WEIS E. Chlorophyll fluorescence and photosynthesis: the basics[J]. Annual Review of Plant Physiology, 2003, 42(42): 313–349.
[23] 蒋运生, 柴胜丰, 唐辉, 等. 光照强度对广西地不容光合特性和生长的影响[J]. 广西植物, 2009, 29(6): 792–796.
[24] 梁永富, 王康才, 薛启, 等. 高温强光胁迫下水杨酸对多花黄精生理及光合特性的影响[J]. 南京农业大学学报, 2018, 41(5): 839–847.
[25] MAXWEL K., JOHNSON G N. Chlorophyll fluorescence —a practical guide[J]. Journal of Experimental Botany, 2000, 51(345): 659–668.
[26] 段龙飞, 尚爱芹, 杨敏生, 等. 中华金叶榆不同无性系光合特性和叶绿素荧光参数日变化研究[J]. 中国农业科技导报, 2014, 16(6): 21–27.
[27] GENTY B, BRIANYAIS J M, BAKER N R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence[J]. Biochimica et Biophysica Acta (BBA) - General Subjects, 1989, 990(1): 87–92.
[28] 王建华, 任士福, 史宝胜, 等. 遮荫对连翘光合特性和叶绿素荧光参数的影响[J]. 生态学报2011, 31(7): 1811–1817.
[29] 张其德. 外加因子对生长在不同光强下的小麦叶绿体荧光特性的影响[J]. 生物物理学报, 1988, 4(3): 192–196.
[30] WANG Y, NII N. Changes in chlorophyll, ribulose bisphosphate carboxylase-oxygenase, glycine betaine content, photosynthesis and transpiration in leaves during salt stress[J]. Journal of Horticultural Science & Biotechnology, 2015, 75(6): 623–627.
[31] 徐晓昀, 郁继华, 颉建明, 等. 2, 4-表油菜素内酯对亚适温弱光下黄瓜幼苗光合特性和抗氧化系统的影响[J]. 核农学报, 2017, 31(5): 979–986.
Effects of different habitats on the growth, chlorophyll content and chlorophyll fluorescence characteristics of medicinal and edible plantsLind.
CHEN Lanying1, XU Hang2, XU Zhenggang2, XIAO Juan2, *
1. College of life science, China West Normal University, Nanchong 637000, China 2. College of Environmental Science and Engineering, China West Normal University, Nanchong 637000, China
Lind. is a traditional medicinal and edible plant; its health effects and ecological benefit values have been paid more and more attention. In order to explore the adaptive response mechanism ofin different habitats and make use of the wild resources, the plant height, leaf morphology, chlorophyll content and chlorophyll content and chlorophyll fluorescence in canopy gap, forest edge and understory were measured and compared in the hill region of northeast of Sichuan. The results were as follows: (1) The plant height, leaf length and area ofin different habitats were significantly different. In the canopy gap habitat was the highest and had significant negative correlations with relative light intensity, positively significantly correlated with humidity. (2) The chlorophyll content had significant differences, which was consistent with the growth of plant morphology. The canopy gap habitat was significantly higher than forest edge and understory. (3) The difference of chlorophyll fluorescence parameters between different habitats were significant different. The actual photochemical efficiency (Φ) in canopy gap habitat was higher, while the chemical quenching coefficient () was lower, and a higher apparent electron transport rate () indicating thathad high photorespiration.had the characteristics of combining shade plant and heliophyte, the canopy gap habitat was better than forest edge and understory, and the plant morphology and photosynthetic characteristics ofwere considered to greatly reflect the adaptation of plants to different environmental regimes. When introducing and cultivating, the relative light (56.05%±0.78%) and relative humidity (52.78%±0.63%) can be used as reference for shading cultivation and field management.
; plant height; leaf traits; chlorophyll content; chlorophyll fluorescence
10.14108/j.cnki.1008-8873.2021.05.009
Q948
A
1008-8873(2021)05-069-09
2020-04-03;
2020-05-15基金项目:国家青年科学基金(31700387); 西华师范大学青年资助专项(18D049)
陈兰英(1987—), 女, 四川南充人, 硕士, 助教, 主要从事植物生态学研究, E-mail: 284626264@qq.com
通信作者:肖娟, 女, 博士, 副教授, 主要从事植物学、环境生态学研究, E-mail: xiaojuanhj@163.com
陈兰英, 许航, 徐正刚, 等. 生境对药食两用植物接骨草的生长、叶绿素含量与叶绿素荧光特性的影响[J]. 生态科学, 2021, 40(5): 69–77.
CHEN Lanying, XU Hang, XU Zhenggang, et al. Effects of different habitats on the growth, chlorophyll content and chlorophyll fluorescence characteristics of medicinal and edible plantsLind.[J]. Ecological Science, 2021, 40(5): 69–77.
