SCN2B在APP/PS1小鼠脑组织中对APP加工处理的作用
2021-11-13赵浩然严国纪董晓函习杨彦彬
赵浩然,李 珊,严国纪,董晓函,习杨彦彬
(昆明医科大学 基础医学院神经科学研究所,云南 昆明 650500)
随着我国老年人口的不断增长,人口老龄化所带来的健康问题也日益严峻。随着年龄的增加,脑老化导致包括学习记忆、感觉运动和认知能力等一系列大脑机能衰退[1]。随着认知功能的持续下降,很有可能进一步发展为以学习记忆能力严重减退为主要表现的阿尔兹海默病(Alzheimer disease,AD)。AD作为一种隐匿的进行性神经退行性疾病,是目前导致老年人最终发生失智或痴呆(Dementia)的主要原因。Aβ产生的一个关键因素是淀粉样前体蛋白(Amyloid precursor protein,APP)被蛋白酶水解成不同活性的片段。目前,AD的病理学改变主要为β淀粉样蛋白(Amyloid-β,Aβ)在功能脑区的过度生成和累积,这是神经纤维缠结产生、神经变性以及神经炎症的主要因素[2]。目前,针对AD多以对症、支持治疗为主,缺乏有效的治疗手段。而对AD发病机制,尤其是对AD病理进程中Aβ产生的机制做进一步阐明,可为疾病的精准治疗指引正确方向。
最近一系列研究表明,电压门控性钠离子通道(Sodium channel-voltage-gated,VGSC)作为中枢神经元动作电位产生和传递的基础,在心脏传导相关疾病、癫痫、神经性疼痛、神经系统退行性疾病等中扮演着重要的角色[3]。神经元动作电位的产生通常由电压门控钠离子通道去极化介导,钠离子通道由一个核心α亚基和两个辅助β亚基(β1和β2)共同组成蛋白质复合体,通过脑内的SCN1A、SCN1B和SCN2B等基因编码产生。其中辅助β亚基虽然不直接参与离子通道开放或失活等主要生物学功能组成,但在细胞信号传递、维持细胞膜稳定性、调节部分离子通道门控特异性等方面的作用同样不可忽视。现有研究表明这些钠离子通道蛋白结构与功能的变化导致的钠离子内流改变在神经退行性相关疾病中起着关键作用[4],这些研究提示钠离子通道干预药物可能在AD治疗中是一个潜在的作用靶点。
本研究通过分析电压门控性钠离子通道二型β亚基(Sodium channel-voltage-gated-beta2,SCN2B)在AD模式动物脑内参与APP介导的Aβ加工和处理中发挥的作用,以阐述AD疾病中的一种潜在的调控机制,进一步验证SCN2B与AD病理性APP代谢途径、Aβ沉积之间的可能关系。
1 材料与方法
1.1 实验动物APPswe/PS1ΔE9 (APP/PS1) 转基因小鼠由合作伙伴澳大利亚莫纳什大学肖志诚教授惠赠。SCN2B-/-、SCN2B+/- 及SCN2B+/+ 三种转基因小鼠均由协和医科大学中国医学科学院实验动物研究所张连峰教授实验室团队提供。分别选取6~8周龄的不同性别的APP/PS1分别与SCN2B-/-、SCN2B+/- 及SCN2B+/+转基因小鼠按照(2雌1雄)方式合笼杂交,获得APP+SCNB2-/-(APP/PS1SCN2B低表达纯合子)、APP+SCN2B+/-(APP/PS1SCN2B杂合子)和APP+SCN2B+/+(APP/PS1SCN2B过表达纯合子)3种转基因小鼠。小鼠均在昆明医科大学SPF级实验动物房饲养,温度控制在22°C左右,每日12 h光照、12 h黑暗,自由摄食和饮水。
1.2 主要试剂 DEME购买于以色列BI公司(biological industries);胎牛血清及Neurobasal购买于GIBCO公司;鼠尾DNA检测试剂盒购买于北京全式金公司;Trizol试剂盒购自biosharp公司;Revert AidcDNA试剂盒购买于美国fermentas公司;SCN2B抗体购买于以色列Alomone Labs公司;抗鼠抗体和GAPDH1抗体(sc47724和sc516132)购买于Santa Cruz公司;ELISA 试剂盒购自于美国Andygene公司。
1.3 转基因小鼠基因型检测 取3~7 d新生小鼠,每只约剪0.5 cm鼠尾组织,使用鼠尾DNA检测试剂盒按步骤提取总DNA,样本置于﹣80°C保存,特异性引物序列(见表1)。分别加入PCR Mix10 μL,PCR上下游引物各0.6 μL,DNA5 μL,双蒸水3.8 μL构成PCR总反应体系,样本置于PCR仪进行扩增。反应条件为:循环35次,94℃变性1 min,59℃退火30 s,72℃延伸1 min,结束后置于4℃保存。PCR产物用1%琼脂糖凝胶电泳,用紫外凝胶成像仪(Bio-Gel,Bio-red)进行可视化分析。获得相应的目的片段即为所需该品系转基因小鼠。
1.4 RT-PCR检测转基因小鼠脑脑皮质与海马中SCN2B的表达 分别选取5种品系转基因小鼠,用戊巴比妥按照(1 mL/kg)进行腹腔注射麻醉,后取出脑组织并分离出海马组织和脑皮质部分。使用Trizol试剂盒提取总RNA。利用primer 6.0设计引物(见表1)。使用Revert AidcDNA合成试剂盒(Fermentas Company)合成cDNA。然后加入PCR Master Mix Kit置于PCR仪进行PCR 扩增,反应条件为:循环35次,94°C下变性1 min、退火1 min,72℃下延伸1 min。获得的PCR产物在1%的琼脂糖凝胶中电泳,用紫外凝胶成像仪(Bio-Gel,Bio-Rad)进行可视化分析。使用Imager J 6.0软件进行图像分析。

表1 引物序列
1.5 Western blot检测转基因小鼠脑脑皮质与海马中SCN2B的表达 戊巴比妥按照(1 mL/kg)进行腹腔注射以麻醉小鼠,取出脑组织,收集海马和脑皮质部分置于1.5 mL离心管中,分别加入10 mM Tris-HCl缓冲液(pH 7.4),10 mM EDTA,30%Triton-1000、10%SDS,蛋白酶抑制剂混合物(Roche)和NaCl,在冰盒上使用匀浆机进行匀浆。随后置于离心机中以12 000 rpm/min,4°C条件下离心30 min。配置10%SDS-PAGE凝胶进行电泳,保留目的蛋白一并蛋白质印迹分析。转移到PVDF膜上,将膜放入5%脱脂奶粉封闭1 h,随后将PVDF膜与抗SCN2B抗体(1∶800)和抗GAPDH(1∶500)一起孵育抗体在4℃过夜。TBST洗涤后,再将膜与抗鼠抗体(1∶1 000)在25°C条件下孵育2 h。Image J 6.0软件计算蛋白质条带的密度。
1.6 细胞培养及分组 选用新生的5种品系小鼠,包括野生型(WT)、APP/PS1+、APP/PS1+SCNB2-/-、APP/PS1+SCN2B+/- 、APP/PS1+SCN2B+/+。每个品系各5只,断颈处死后,手术器械剥离颅骨与硬脑膜后迅速分离脑皮质与海马组织放入完全培养基,经木瓜酶消化20 min,过滤后收集细胞悬浮液进行神经原代培养;第1天12 h全量更换神经元培养基,随后每24小时半量更换神经元培养基。
1.7 ELISA检测Aβ40和Aβ42的水平 神经元培养5d后取培养基与细胞溶解产物。按照Aβ40ELISA 试剂盒(AD2170MO)、Aβ42ELISA试剂盒(AD2169MO)的说明书指示将标准品梯度稀释,绘制标准曲线。1 500 rmp/min离心10 min后收集细胞上清液样品按照每孔加入50 μL含酶试剂样品至酶标板,封板膜封板37°C恒温箱孵育30 min,静置1 min后弃液,重复5次,酶标仪设置为450 nm后依次测量各孔OD值。比较各品系转基因小鼠神经元细胞中的Aβ40和Aβ42的表达水平。
1.8 Western blot检测蛋白含量 收集细胞悬浮液,除去残留的培养基并离心,后加入裂解液提取总蛋白。配置10%SDS-PAGE凝胶进行电泳,保留目的蛋白部分转移到PVDF膜上,将膜放入5%脱脂奶粉封闭1 h。在4°C下分别与sAPPα(1∶100)、sAPPβ(1∶500)、APP C-Terminal(APPβ-CTF,1∶500)、AICD(1∶1 000)抗体孵育过夜,随后用Image J 6.0软件测量条带密度。
事实上,当{xn}关于度量d收敛到0时,对任意的ε>0,存在N,使得d(xn,0)=xn<ε。此时ρ0(xn,0)=1-(xn→0)∧(0→xn)=1-(1-xn)=xn>ε。此时ρ0(xn,0)=1-(xn→0)∧(0→xn)=1-(1-xn)=xn

2 结果
2.1 转基因小鼠基因型鉴定 分别选取SCN2B-/-、SCN2B+/-及SCN2B+/+转基因鼠与APP/PS1小鼠交配,运用特异性引物对各品系转基因小鼠后代鼠尾组织提取的DNA进行PCR检测,获得相应目的片段的样品对应为该品系转基因小鼠。由此筛选出APP/PS1遗传背景的APP/PS1+SCN2B-/-、APP/PS1+SCN2B+/-、APP/PS1+SCN2B+/+转基因鼠小鼠,后将F1代同胞兄妹交配持续扩大繁殖数量,PCR检测结果如图1所示。结果显示SCN2B-/-转基因组新生小鼠均表达SCN2B-/-敲除插入的300bp片段(A);SCN2B+/-转基因组新生小鼠均表达SCN2B+/-部分敲除插入的456bp片段(B);SCN2B+/+转基因组新生小鼠均表达SCN2B+/-部分敲除插入的250bp片段(C);说明本实验按照预期获得了相应的转基因小鼠品系(见图1)。
2.2 转基因小鼠脑皮质海马中SCN2B的表达检测 为验证APP/PS1+SCN2B-/-、APP/PS1+SCN2B+/-、APP/PS1+SCN2B+/+各品系转基因小鼠SCN2B的转基因干预和过表达效率,分别取各品系小鼠脑皮质及海马进行检测,每组5例(n=5),检测方法为实时荧光定量PCR及Western blot。结果显示,APP/PS1+SCN2B-/-转基因小鼠组海马及脑皮质组织中SCN2B的表达量几乎检测不到,而APP/PS1+SCN2B+/+小鼠与野生型相比,SCN2B的表达显著增加,几乎增加了2.2倍;APP/PS1+SCN2B+/-小鼠中表达则为APP/PS1+SCN2B+/+转基因小鼠的60%左右。检测结果证实,成功构建APP/PS1+SCN2B-/-,APP/PS1+SCN2B+/-以及APP/PS1+SCN2B+/+转基因小鼠(见图2)。

A:与WT型比,P<0.05,n=5;SCN2B在上述各品系转基因小鼠脑皮质中的表达蛋白质印记条带;B:与WT型比,P<0.05,n=5;SCN2B在上述各品系转基因小鼠海马中的表达蛋白印迹条带。 图2 SCN2B在APP/PS1遗传背景的SCN2B-/-,SCN2B+/-以及SCN2B+/+转基因小鼠脑皮质和海马中表达的蛋白印迹结果
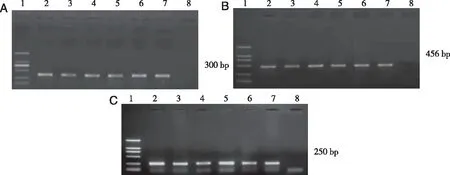
A:APP/PS1+SCN2B-/-转基因小鼠基因型检测;Lane 1:DNA MarkerDL 2,000 (从上到下分别为:2 000 bp,1 000 bp,750 bp,500 bp,250 bp,100 bp)。Lane 2-7:转基因鼠的鼠尾DNA PCR扩增产物,Lane 8:WT;B:APP/PS1+SCN2B+/-转基因小鼠基因型检测;Lane 2-7:转基因鼠的鼠尾DNA PCR扩增产物,Lane 8:WT;C:APP/PS1+SCN2B+/+转基因小鼠基因型检测:Lane 2-7:转基因鼠的鼠尾DNA PCR扩增产物,Lane 8:WT。 图1 APP/PS1+SCN2B-/-,APP/PS1+SCN2B+/-以及APP/PS1+SCN2B+/+转基因小鼠基因型检测
2.3 Aβ40和 Aβ42在不同原代培养神经元中的表达水平APP/PS1+组小鼠脑皮质海马神经元中的Aβ40和Aβ42含量与APP/PS1+SCN2B-/-、APP/PS1+SCN2B+/-、APP/PS1+SCN2B+/+3种品系小鼠比较明显降低(P<0.05)。APP/PS1+SCN2B+/+小鼠与APP/PS1+SCN2B-/-、APP/PS1+SCN2B+/- 组相比,Aβ40和 Aβ42含量相比明显升高(P<0.05)。提示SCN2B基因过表达具有促进Aβ生成的作用(见图3)。
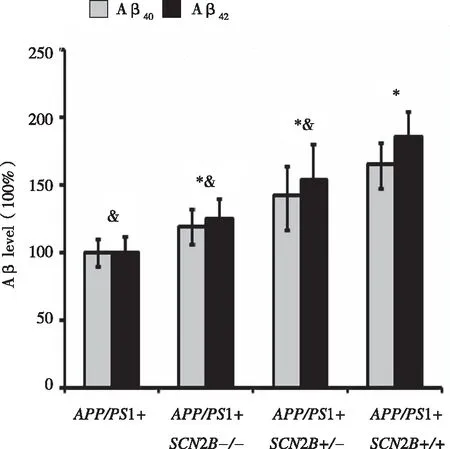
*:与APP/PS1+组比,P<0.05,n=4;&:与APP/PS1+ SCN2B+/+组比较,P<0.05,n=4。 图3 β40 和 Aβ42在各组细胞培养溶解产物中的表达水平
2.4SCN2B对不同品系转基因小鼠细胞中APP酶解产物sAPPα,sAPPβ,APP-APPβ-CTF及AICD的表达水平 与APP/PS1+SCN2B+/+组小鼠相比较,WT组小鼠sAPPα,sAPPβ含量均升高,但APPβ-CTF与AICD片段含量处于相反的趋势(P<0.05);在APP/PS1+SCN2B+/-组小鼠中,sAPPα、sAPPβ含量呈现减少的趋势,APP-APPβ-CTF与AICD表达显著增加(P<0.05);而与APP/PS1+组小鼠对比,sAPPα、sAPPβ含量表现出随SCN2B的表达增加而减少,而APPβ-CTF与AICD片段含量则随SCN2B的表达增加而增加(P<0.05)。此结果提示SCN2B基因对sAPPα、sAPPβ、APPβ-CTF和AICD的合成具有一定的调控作用(见图4)。
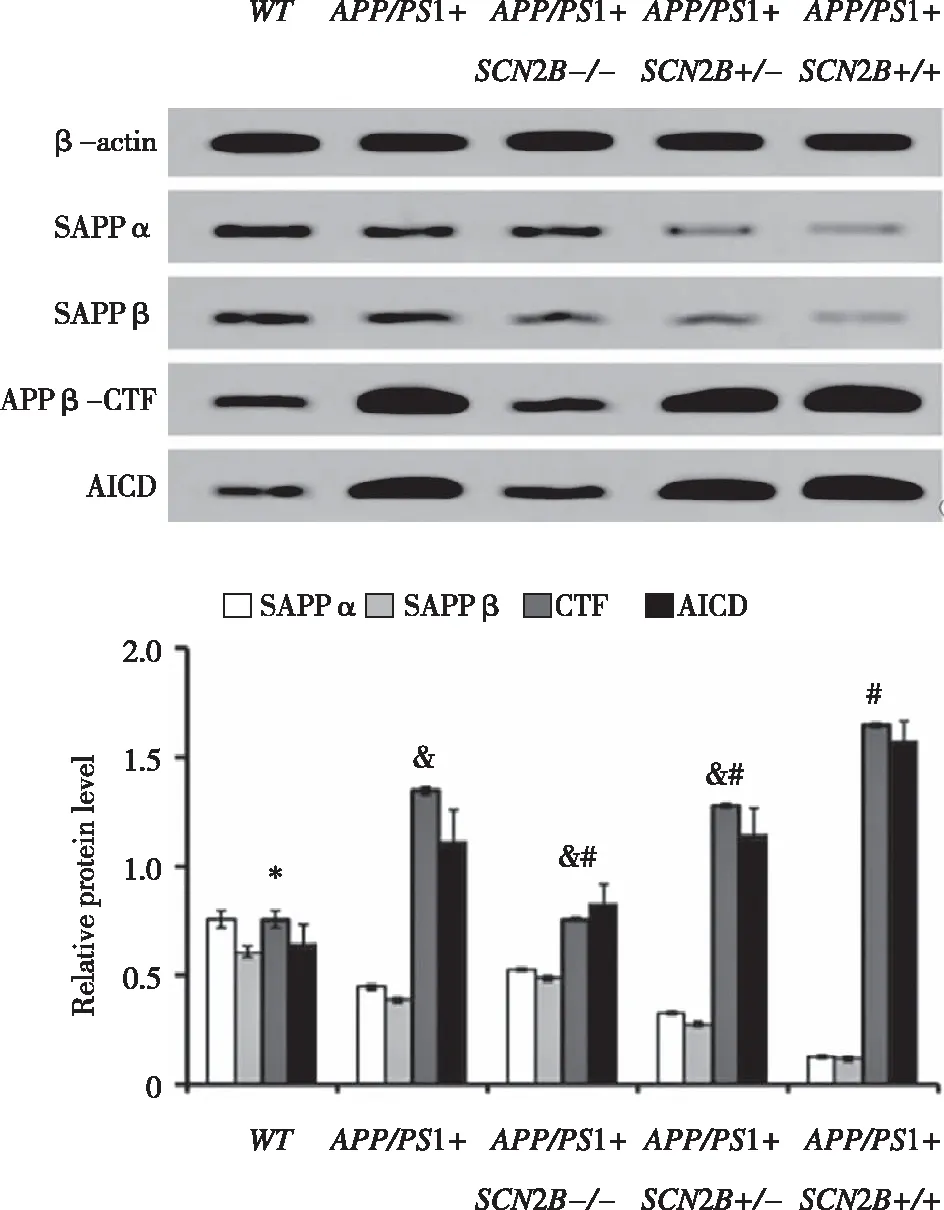
*:与APP/PS1+ SCN2B+/+组比较,P<0.05,n=4;&:与WT组比较,P<0.05;#:与APP/PS1+组比较,P<0.05,n=4。 图4 各品系细胞培养基中sAPPα,sAPPβ,APP-APPβ-CTF及AICD表达的蛋白印迹结果
3 讨论
AD的主要病理改变以淀粉样蛋白(Aβ)在正常脑区异常聚集形成的老年斑块为特征,此过程与APP蛋白的酶解过程密切相关[6]。APP的代谢途径主要有两条:淀粉样代谢途径及非淀粉样代谢途径。APP经由淀粉样代谢途径降解的过程如下:APP被β-分泌酶水解切割成可溶性APPβ片段(sAPPβ),C-末端片段(APPβ-CTF),随后被位于细胞膜上的γ-分泌酶分解释放出Aβ[7]。Aβ主要由40个和42个淀粉样肽残基(Aβ40和Aβ42)组成,Aβ寡聚体形成是导致AD患者的记忆能力衰退和神经系统毒性的关键因素,不同长度Aβ的相互聚集形成Aβ寡聚体进而表现出对神经元的毒性损伤作用[8]。而在非淀粉样蛋白途径中,APP可被α-分泌酶切割,释放可溶性片段(sAPPα),经微循环代谢排出,加速Aβ溶解并减轻Aβ对神经元的持续性损伤[9-10]。由此,对APP代谢过程及其代谢产物的监测有助于观察疾病进展及药物疗效评估。
已有研究发现SCN2B作为参与AD及脑老化的相关基因,虽不直接参与离子通道的组成,但在调节功能性亚基功能蛋白在细胞膜的表达和稳定性、信号传导、通道的激活与失活中具有重要作用[11]。SCN2B可通过调控钠离子通道结构,从而影响神经元的钠离子内流及电生理特性改变。近年来,随着研究的不断深入,还发现SCN2B可间接调控质膜中的钠通道的数量,增加钠离子峰值电流的密度提高去极化速度,从而提高钠通道从激活向失活状态的恢复效率[12]。越来越多的证据表明,SCN2B编码的钠离子通道亚基蛋白上的Ig结构域可以与其他细胞黏附分子相互作用,发挥类似细胞粘附功能[13]。但是,关于它们在老化相关的分子机制与病理性改变,尤其是与学习记忆相关功能衰退过程中的作用及机制却鲜有报道。笔者在前期研究中准备了SCN2B基因敲低小鼠模型,表型检测结果显示SCN2B基因下调60.68%可以显著改善转基因小鼠空间认知记忆能力[14],提示SCN2B可能与小鼠认知功能相关。APP/PS1小鼠在6月龄左右可表现出空间记忆能力丧失及AD相关症状,是目前国内外研究AD的使用疾病模型[15]。
本研究通过制作APP/PS1+SCN2B-/-、APP/PS1+SCN2B+/-、APP/PS1+SCN2B+/+ 3种转基因小鼠疾病模型,发现在5月龄会表现出与APP/PS1+转基因小鼠相似的认知功能障碍症状,通过实时荧光定量PCR及免疫印迹检测3种转基因动物SCN2BmRNA和蛋白表达量,结果显示APP/PS1+SCN2B-/-组海马及脑皮质组织中SCN2B表达量几乎检测不到,但与野生型相比,SCN2B+/+组小鼠的SCN2BmRNA和蛋白表达量均显著增加,几乎增加了2.2倍,而SCN2B+/-组SCN2B的表达则为野生型的60%左右。结果表明,笔者基于APP/PS1小鼠的遗传背景成功构建了SCN2B-/-(敲除)、SCN2B+/-(敲低)以及SCN2B+/+(过表达)转基因小鼠。接下来,本研究用ELISA法检测了Aβ在各组转基因小鼠脑皮质与海马神经元细胞中的表达水平,结果发现APP/PS1+SCN2B+/+组脑内Aβ含量较APP/PS1+SCN2B-/-组显著增多,且当动物体内SCN2B表达增加时,Aβ含量呈现随SCN2B增加而增加的趋势;还发现过表达SCN2B可以促进APP向致病性片段Aβ40和Aβ42的分解过程,而将SCN2B敲除可以显著抑制这一过程。随后,通过验证SCN2B在APP不同酶解途径中的调控作用,证实了过表达SCN2B可以促进APP更多的通过β-分泌酶途径分解为致病性的APPβ-APPβ-CTF、AICD以及sAPPβ片段,减少经α-分泌酶途径酶解为水溶性可代谢的sAPPα片段。结果提示敲低SCN2B可以影响APP酶解过程,有利于APP向生成可溶性Aβ的方向发展,减少不溶性神经毒性的Aβ在正常脑区的过度聚集。
笔者前期在对APP/PS1小鼠进行SCN2B基因敲低处理后,发现其可诱导脑啡肽酶(Neprilysin,NEP)的活性增高和表达水平增加,增强NEP与APP致病性代谢产物AICD的结合,进而促进APP/PS1小鼠脑内淀粉样蛋白的清除[16]。还有研究表明SCN2B基因敲低可上调Nav1.2、Nav1.6在轴突中的表达,对神经电位传导恢复具有一定促进作用,Nav1.1虽然也检测到表达变化,但在神经元中的具体作用还不清楚[17]。尽管β2亚基不直接影响Na电流水平,但它的存在对维持神经元细胞结构和通道蛋白的稳定性是必需的。同时,β2亚基与α亚基共价结合形成复合物也是发挥限制功能性Na+通道电流速率的关键因素,SCN2B在神经元兴奋性调节,Na通道密度和细胞膜定位等方面起着十分重要的作用[18]。接下来对β4亚基的氨基酸序列研究发现它与β2亚基的氨基酸序列同源性>35%,提示两种亚基在参与Na通道蛋白形成过程中存在相似作用,所以其在神经元稳定性方面的作用不可忽视。这一系列研究表明,SCN2B可能是基于调控神经元轴突Na通道的密度与结构表达来发挥相应生物学功能。
结合文献报道以及本研究的发现,敲低SCN2B有利于推动APP向生成可溶性Aβ的方向酶解,减少不溶性神经毒性的Aβ在正常脑区的过度聚集,从而发挥神经保护作用。本研究部分揭示了SCN2B在对APP 加工处理及Aβ产生中的作用,为神经保护特别是AD药物的研发提供了潜在作用靶点。但SCN2B调控APP的酶解进程是否有助于认知功能改善还有待进行更深入的研究证实。
