草菇多酚氧化酶基因的克隆与表达分析
2021-11-10刘海峰何焕清肖自添罗学梅
刘 明,刘海峰,周 慧,何焕清,肖自添,罗学梅
(1.广东省农业科学院蔬菜研究所/广东省草菇科技创新中心/广东省蔬菜新技术研究重点实验室,广东 广州 510640;2.广东省现代农业装备研究所,广东 广州 510630;3.广东省农业技术推广中心,广东 广州 510520)
【研究意义】草菇(Volvariella volvacea),别名南华菇、兰花菇、美味草菇及中国菇等,是一种生长于热带、亚热带地区的食用菌,其人工栽培技术起源于中国[1]。新鲜草菇肉质脆嫩,易受到机械损伤,因具有强烈的呼吸作用和代谢活动,采后1~2 d 子实体萎缩褐变,价值显著降低,新鲜草菇长时间贮藏和远距离运输较难实现[2]。研究发现,草菇发生褐变主要与多酚氧化酶(Polyphenol oxidase,PPO)活性、维生素C 及总酚含量有关。在PPO 作用下酚类底物氧化形成黑色素的醌类物质,促使草菇快速褐变[3]。在运输过程中受到机械损伤的影响,草菇组织破裂会导致酚类底物与PPO 接触,从而加速褐变反应[4]。克隆并分析草菇PPO 基因家族对草菇抗褐变品种选育及改良有重要参考价值。【前人研究进展】PPO 是一种广泛存在于植物、动物、细菌和真菌中的含铜酶。PPO 可根据底物特异性和结构分为酪氨酸酶(EC 1.14.18.1)、儿茶酚氧化酶(EC 1.10.3.1)和漆酶(EC 1.10.3.2)[5]。在食用菌中如双孢蘑菇(Agaricus bisporus)、香菇(Lentinula edodes)、滑子菇(Pholiota nameko)、平菇(Pleurotus ostreatus)、玉木耳(Auricularia cornea)等已鉴定多个多酚氧化酶,其中在香菇中分离到1 个多酚氧化酶基因,表达量分析发现该基因在菌丝期到转色期表达量逐渐升高,在原基形成后表达量降低,该基因可能与香菇的转色有关[6-7];在对滑子菇的研究中分离到1 个42 ku 的多酚氧化酶,该酶以活性前体的形式表达,在蛋白酶的作用下切割C 端序列而生成有活性的成熟酶[8];在平菇中分离到1 个75 ku 的酪氨酸酶,该酶具有较高比活力,可用于食品工业[9];在玉木耳中克隆到1 个多酚氧化酶,其在菌丝期表达量较低在子实体中表达量较高[10]。在双孢菇中已克隆鉴定出6个PPO 基因[11],利用CRISPR-Cas9 基因编辑系统对双孢蘑菇基因组中1 个多酚氧化酶基因进行编辑,使得PPO 酶活降低30%,有效减缓了双孢蘑菇的褐变现象[12]。
【本研究切入点】草菇基因组测序的完成对草菇基因功能的研究起到了极大的促进作用[13]。本研究通过生物信息学分析,从草菇中筛选和克隆获得PPO 基因家族成员,分析其家族序列特征,分析该家族在各个发育时期和组织部位的表达差异,为进一步进行遗传改良提供参考。【拟解决的关键问题】分析研究草菇中PPO 基因家族的结构特点和表达模式,以期为解析草菇褐变机理积累数据,为草菇抗褐变、耐存储品种选育等提供支撑。
1 材料与方法
1.1 试验材料
供试草菇菌株v26,保存于广东省农业科学院蔬菜研究所食用菌菌种保藏中心。菌丝培养采用PDA 培养基,出菇培养基为50%稻草、50%棉籽壳,含水量65%。
1.2 试验方法
出菇试验于2021 年3 月在广东省农业科学院钟落潭白云试验基地进行,于菌丝期、原基期、蛋形期、成熟期4 个不同时期取样。另外,在蛋形期对外菌膜、菌柄、菌盖3 个不同部位进行取样,迅速用液氮冷冻,然后于-80 ℃保存,备用。
1.2.1 VvPPO 基因家族序列获得 根据广东省蔬菜新技术重点实验室2015 年测得的草菇菌丝转录组结果(PRJNA408191)和公布的草菇基因组信息(NCBI 登录号:GCA_000349905),筛选得到功能注释为PPO 的4 个基因全长序列,分别命名为VvPPO1(MZ322862)、VvPPO2(MZ440853)、VvPPO3(MZ440854)、VvPPO4(MZ440855)。据此设计CDS全长引物,设计VvPPO1、VvPPO2、VvPPO3和VvPPO4基因序列引物和CDS 引物,引物信息见表1。
采用生工生物工程(上海)股份有限公司总RNA 提取试剂盒提取草菇菌丝RNA,采用赛默飞有限公司生产的逆转录试剂盒合成cDNA,采用CTAB 法提取草菇DNA。以草菇菌丝cDNA 和基因组DNA 为模板进行扩增。PCR 扩增产物纯化后,与pMD18-T 载体进行连接,阳性克隆进行测序。
1.2.2 生物信息学分析 蛋白质一级结构分析采用ProtParam(https://web.expasy.org/protparam/)在线程序,三级结构分析采用SWISSMODEL(https://swissmodel.ex pasy.org/interactive/);蛋白序列相似性分析运用LALIGN(https://www.ebi.ac.uk/Tools/psa/lalign/);基因结构分析运用Simple Modular Architecture Research Tool(http://smart.embl-heidelberg.de/)。
1.2.3 进化树分析 利用 Clustal W 对草菇、双孢蘑菇、香菇、巴西蘑菇(Agaricus blazei)、烟曲霉(Aspergillus fumigatus)、粗糙脉孢菌(Neurospora crassa)的PPO 氨基酸序列进行多重序列比对,并在 MEGA7 软件中使用邻位相连法(Neighborjoining)构建进化树[14],并进行1 000 次抽样置换检测。
1.2.4 VvPPO 基因家族表达分析 以草菇不同发育时期和蛋形期不同组织部位的cDNA 为模板,以VvTUBα为内参[15],反应引物见表1。采用擎科2×T5 Fast qPCRMix (SYBRGreen)试剂盒,PCR 反应程序:95 ℃预变性30 s;95 ℃变性5 s、58 ℃退火30 s,40 个循环;每个反应重复3 次,采用2-ΔΔCt计算基因的相对表达量[16]。

表1 供试引物信息Table 1 Information of tested primer sequences
2 结果与分析
2.1 VvPPO 基因家族的获得
测序结果表明,VvPPO1CDS 全长1 596 bp(图1 A),含7 个外显子和6 个内含子;VvPPO2CDS 全长2 685 bp(图1B),含9 个外显子和8 个内含子;VvPPO3CDS 全长1 593 bp(图1C),含7 个外显子和6 个内含子;VvPPO4CDS 全长2 067 bp(图1D),含14 个外显子和13 个内含子。
2.2 VvPPO 基因家族编码蛋白质特征分析
ProParam 分析表明,草菇PPO 基因家族所编码蛋白的氨基酸数目在530~894 之间,相对分子量最小的是VvPPO1,最大的是VvPPO2;理论等电点pI 范围5.34~5.95;蛋白的不稳定系数均大于40,表明草菇PPO 蛋白为不稳定蛋白;总平均疏水指数均为负值,表明4 个草菇PPO 蛋白为亲水蛋白(表2)。
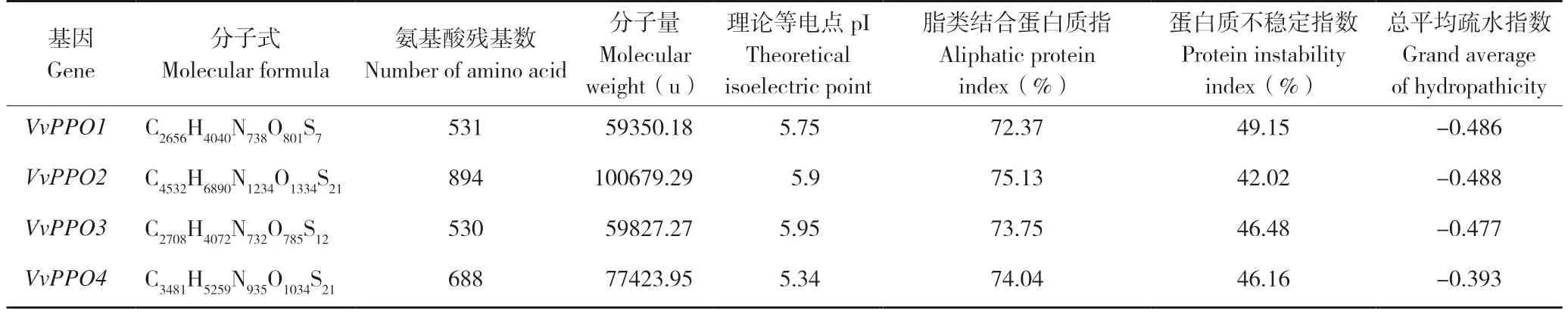
表2 VvPPO 蛋白理化性质预测Table 2 Prediction of physical and chemical properties of VvPPO gene family
SMART 分析表明,4个草菇PPO 蛋白的Tyrosinase 结构域长度为236~335 个氨基酸(图2)。双孢蘑菇 PPO 催化活性中心都包括2 个保守的含铜结构域CuA 和CuB,其中含高度保守的3 个组氨酸,蛋白序列比对显示草菇PPO 蛋白,酪氨酸酶CuA 和CuB 结合位点保守(图3)。Swiss-model 分析表明,三级结构中间有2 个铜离子结合域(图4)。
2.3 VvPPO 蛋白的同源比对与进化树分析
蛋白序列相似性分析显示(表3),VvPPO蛋白序列相似度介于48.1%~73.5%,其中VvPPO1与VvPPO3 最高相似度为63.1%,VvPPO2 与VvPPO4 最高相似度为73.5%。将草菇PPO 蛋白序列与双孢蘑菇、香菇、巴西蘑菇、烟曲霉、粗糙脉孢菌PPO 蛋白序列进行进化树分析,VvPPO1、VvPPO3 与双孢蘑菇、香菇、巴西蘑菇PPO 聚为一类,VvPPO4 与烟曲霉聚为一类,VvPPO2 单独聚为一类(图5)。

表3 VvPPO 蛋白序列相似性Table 3 Sequence similarity of VvPPO Proteins
2.4 VvPPO 基因家族表达分析
VvPPO基因在不同发育时期的表达量分析表明,4 个多酚氧化酶基因在草菇各个发育时期中都有表达。VvPPO1在原基期表达量最高,随着发育的成熟表达量逐渐降低;VvPPO2在菌丝期表达量最高;VvPPO3在原基期表达量最高,其在各个时期的表达模式和VvPPO1较为相似;VvPPO4在菌丝期表达量最低,蛋形期表达量最高(图6 A)。VvPPO基因在蛋形期不同组织部位的表达量分析表明,VvPPO1、VvPPO4在外菌膜中表达量较高,在菌盖和菌柄中表达量较低,其中VvPPO1在外菌膜的表达量要比在菌盖和菌柄中的表达量高10 倍以上;VvPPO2、VvPPO3在菌柄中的表达量较高,在外菌膜和菌盖中表达量较低(图6 B)。
3 讨论
研究发现PPO 与果蔬发生褐变有关[17],在大花金鸡菊(Coreopsis grandif lora)的研究中发现多酚氧化酶AUS1 与aurone 的合成有关[18],说明PPO 功能的多样性。草菇PPO 家族蛋白序列相似性分析显示,草菇VvPPO1 与VvPPO3 相似度较高,在不同发育时期的表达模式也较相似;三级结构预测显示VvPPO2 与另外3 个草菇PPO蛋白结构差异明显;草菇PPO 蛋白序列与双孢蘑菇、香菇、巴西蘑菇、烟曲霉、粗糙脉孢菌PPO序列进化分析结果显示,VvPPO2 单独聚为一类,说明VvPPO2 与另外3 个PPO 生物功能不同。
不同的PPO基因表达模式不同,在植物不同发育时期表达量差异明显,且PPO的表达还受到生物和非生物胁迫的影响[19]。双孢蘑菇的发育过程中,子实体时期PPO 的活性增强,菌盖表面褐变程度与PPO 活性呈正相关关系[6]。对VvPPO基因的表达分析结果显示,除VvPPO2外,其他草菇PPO基因在菌丝期表达量较低,原基形成后表达量迅速升高,到成熟期时表达量渐渐降低,在蛋形期的不同组织部位表达量检测结果显示,在蛋形期PPO 在各个组织部位均有表达。VvPPO1和VvPPO4在外菌膜中的表达量最高,可能是蛋形期引起草菇褐变的关键基因。VvPPO2在外菌膜中表达量最低,在菌柄中表达量最高,VvPPO2在各个组织部位的表达量差异不明显,这意味着草菇PPO 功能的差异。
在果蔬抗褐变的研究中以PPO基因为靶标进行基因编辑已经获得了抗褐变的苹果(Malus sieversii)、茄子(Solanum melongena)和马铃薯(Solanum tubersoum)[20-22]等,如在马铃薯中通过编辑多酚氧化酶基因StPPO2获得了多酚氧化酶活性降低63%、酶促褐变降低73%的植株。在双孢蘑菇中通过编辑PPO基因获得了抗褐变双孢蘑菇[12],本研究初步分析了草菇PPO基因家族的特征和表达模式,为进一步揭示草菇褐变的发生机理、培育抗褐变草菇提供理论依据。
4 结论
本研究基于草菇基因组数据,挖掘获得4个多酚氧化酶基因,蛋白序列分析显示4 个草菇PPO 蛋白中菌含有保守Tyrosinase 结构域,具有PPO 蛋白典型特征,含有铜离子A(CuA)和 B(CuB) 结合区。同源比对与进化分析显示VvPPO1与VvPPO3 相似性较高,VvPPO4 与烟曲霉同源性较高,VvPPO2 单独聚为一类,表明它们可能具有不同功能。草菇PPO基因表达模式分析显示VvPPO1、VvPPO3与VvPPO4子实体时期表达量比菌丝期表达量高,VvPPO2在各个发育时期表达量变化不明显。在蛋形期外菌膜中VvPPO1和VvPPO4表达量较高,VvPPO1和VvPPO4可能与蛋形期草菇褐变有关。
