降解水稻秸秆细菌−真菌复合菌系的构建与评价
2021-11-09梅新兰郑海平李水仙杨天杰江高飞韦中徐阳春沈其荣
梅新兰,郑海平,李水仙,杨天杰,江高飞,韦中,徐阳春,沈其荣
(南京农业大学资源与环境科学学院;江苏省固体有机废弃物资源化高新技术研究重点实验室;江苏省有机固体废弃物资源化协同创新中心;资源节约型肥料教育部工程研究中心;国家有机类肥料工程技术研究中心,南京 210095)
提高秸秆生物腐解的效率是水稻等大宗农作物秸秆直接还田利用研究的热点[1]。细菌和真菌是秸秆中木质纤维素降解的主要参与者,如芽孢杆菌、纤维杆菌等细菌通过大量富集、膨胀和分泌纤维素酶来破坏和分解秸秆;木霉和青霉等真菌通过菌丝的物理作用和分泌大量纤维素酶实现秸秆降解[1−2]。近年来,微生物降解秸秆研究发展迅速,但自然条件下秸秆的生物腐解速率依然较慢,其生物降解效率仍需提高[2]。一方面,秸秆的主要成分是纤维素、半纤维素、木质素等高分子物质,它们之间通过氢键和共价键连接成的网状结构,使纤维素降解相关酶难以直接接触到木质纤维素,从而影响降解效果[3];另一方面,秸秆降解需要细菌、真菌等多种微生物及其分泌的纤维素酶相互协同发挥作用,而单一菌株通常难以完成如此复杂的生化过程,如真菌菌丝对木质纤维素结构的破坏可帮助细菌定殖,细菌对木质纤维素的改性更有利于真菌纤维素酶对木质纤维素的有效降解[4−5]。大量研究表明,不同微生物菌株组合的降解效果比单一菌株好[6]。因此,越来越多的学者开始关注不同降解微生物之间的复配,探究复合菌系在秸秆降解过程中发挥的协同作用。
近年来,国内外学者在秸秆降解复合菌系方面的研究取得了系列进展。王春芳等[7]发现从猪粪−水稻秸秆堆肥中筛选出的复合菌系中细菌菌株种类更为丰富,分解纤维素的能力更强。KAUSAR 等[8]将8 株高效降解真菌进行两两组合,发现复合菌系降解水稻秸秆的效果优于单菌,其中黑曲霉和绿色木霉的组合降解率最高。当前,复合菌系主要通过直接筛选和降解菌人工组合的方式获得。直接筛选的优势在于维持了复合菌系成员间的协同关系,降解效果稳定,菌株 组 成 也 相 对 复 杂 ,如 复 合 菌 系 GF − 系 列[9−10]、RWS1[11]和LDC[12]等。降解菌人工组合的优势在于将功能菌株进行重新组合,可针对性地提高秸秆的降解效果,如复合菌系F1[13]、T8[14]和G[9]等。尽管人工构建的复合菌系种类较为丰富,但主要是细菌或真菌的组合,仅有少量细菌−真菌的组合。如夏强[15]发现地衣芽孢杆菌、黑曲霉和绿色木霉组成的细菌−真菌复合菌系产纤维素酶的能力优于单一菌株,复合菌系显著提高了降解水稻、玉米和小麦秸秆的能力。细菌−真菌组成的复合菌系其菌株的种类、丰度较高,菌株产酶具有协同作用,因此提高了秸秆中纤维素的降解率[16]。目前,在构建细菌−真菌复合菌系降解秸秆的研究中,对于菌株组合方式和种类对复合菌系降解效果和产酶活性影响的认识还不够深入,需开展进一步研究。为此,本研究从腐解的水稻秸秆中筛选出降解能力较强的细菌和真菌,构建降解菌的全组合,探究各组合降解水稻秸秆的效果和产酶活性,最后解析产酶活性与秸秆降解效果之间的关系及其贡献程度,筛选获得高效降解水稻秸秆的降解菌组合,为水稻秸秆资源化利用中复合菌系的构建研究提供理论支持。
1 材料与方法
1.1 试验材料
1.1.1 水稻秸秆
水稻秸秆采自南京市后村。用清水将水稻秸秆表面尘土洗净,晾干,切成2~3 cm的秸秆段,保存备用。将晾干的水稻秸秆段在1%NaOH溶液中浸泡24 h,然后80 ℃恒温水浴20 min,用清水反复冲洗至pH为中性,65 ℃烘干,粉碎过30目筛,制备成水稻秸秆粉备用。
1.1.2 培养基
使用无机盐培养基作为基础培养基,添加抗生素或水稻秸秆粉用于水稻秸秆降解菌的筛选和培养等[17]。细菌富集培养基、真菌富集培养基分别用于水稻秸秆降解细菌、真菌的富集筛选[18];LB培养基、PDA培养基分别用于秸秆降解细菌、真菌的纯化[17];产酶培养基[19]和羧甲基纤维素(CMC)−刚果红培养基[20]用于水稻秸秆降解菌产酶活性的检测。培养基制备完成后,置于115 ℃下高压蒸汽灭菌30 min备用。
1.2 试验方法
1.2.1 秸秆降解细菌和真菌的分离筛选
将1.0 g 自然腐解的水稻秸秆置于三角瓶中,加入 90 mL 无菌水,室温 170 r·min−1振荡 1 h 后静置 30 min,制备悬浮液。将悬浮液分别涂布在细菌富集培养基和真菌富集培养基中,30 ℃静置培养,细菌培养2~3 d,真菌培养3~4 d,待长出明显菌落后随机筛选菌株。细菌在LB 培养基中连续划线培养,纯化3 次,获得秸秆降解细菌分离株的培养液,加入30%甘油保存于−80 ℃超低温冰箱中备用。真菌在PDA 培养基中培养,连续挑选菌丝,分离纯化3 次,获得秸秆降解真菌孢子的悬浮液,置于30%甘油管中,于−80 ℃保存备用。
1.2.2 秸秆降解细菌和真菌的菌株鉴定
以分离得到的水稻秸秆降解细菌和真菌为对象,参考细菌、真菌基因组提取试剂盒的使用说明(OME⁃GA公司),分别提取细菌和真菌基因组DNA,置于−80 ℃超低温冰箱中保存备用。分别采用通用引物27F/1492R[21]和 ITS1/ITS4[22]扩增细菌的 16S rRNA 基因序列和真菌的基因间隔区序列(ITS),对水稻秸秆降解菌进行分子鉴定。细菌和真菌的通用引物序列分别为 27F:5′ − AGAGTTTGATCATGGCTCAG − 3′ 和1492R:5′−TACGGTTACCTTGTTACGACTT −3′;ITS1:5′ − GGAAGTAAAAGTCGTAACAAGG − 3′ 和ITS4:5′−TCCTCCGCTTATTGATATGC−3′。采用Takara PCR 扩增试剂盒 50 µL 反应体系:DNA 模板 1µL,2.5 mmol·L−1dNTP 4 µL,1 mmol·L−1引物各1 µL,10× Loading Buffer 5 µL,25 mmol·L−1MgCl23 µL,5 U·µL−1Taq 酶 0.5 µL,ddH2O 34.5 µL。扩增程序:94 ℃ 5 min,94 ℃变性 30 s,退火温度 52 ℃ 30 s,72 ℃ 1 min,共 30 个循环;72 ℃ 10 min。将细菌 16S rRNA 序列和真菌ITS 序列的PCR 扩增产物送往上海生物工程有限公司测序,将测序结果在NCBI 数据库(https://ncbi.nlm.nih.gov/)中进行BLAST 比对分析,选取相似性>99.5% 的序列在MEGA7 软件(https://www.megasoftware.net)中 构 建 Neighbor −Joining系统发育树。
1.2.3 秸秆降解率的检测
细菌细胞悬浮液或真菌孢子悬浮液的制备:将保存的细菌、真菌分离株分别在LB 培养基平板或PDA培养基中30 ℃活化复苏2~3 d。细菌挑取单菌落过夜培养,制备成1.0×107cfu·mL−1的细胞悬浮液[17];真菌挑取活化后的孢子接种到水稻秸秆粉培养基中产孢,用无菌生理盐水收集真菌孢子,制备成1.0×107cfu·mL−1的孢子悬浮液[13]。
按照1%(V∶V)比例[13,17],接种1 mL细菌细胞悬浮液或者真菌孢子悬浮液到含有100 mL 产酶培养基的500 mL三角瓶中,以秸秆段(5 g)为唯一碳源,置于恒温摇床中发酵,条件为30 ℃、170 r·min−1。连续培养4 d 后,取少量培养液用于提取粗酶液、测定酶活性;连续培养8 d 后,取发酵秸秆段残渣用无菌水反复冲洗干净,65 ℃烘干8 h称质量,计算秸秆降解率(D):
式中:W0为对照秸秆残渣干质量,g;W为处理秸秆残渣干质量,g。各处理重复3次。
1.2.4 纤维素酶活的测定
取 1.2.3 中连续培养 4 d 的发酵液 2 mL,10 000 r·min−1离心5 min 收集上清液,即得粗酶液,用于滤纸酶活性的测定。采用DNS比色法[23],分别测定发酵液中滤纸酶(FPase)、纤维素内切酶(CMCase)和木聚糖酶(Xylanase)的活性。将1 min 内水解生成1 µmol 还原糖所需的酶量定义为一个活性单位(U)。将3种酶活性Z−score标准化后的总和定义为总酶活性。
1.2.5 细菌−真菌复合菌系的构建
通过秸秆降解率和滤纸酶活性评价,分别筛选获得2 株降解秸秆能力较好的细菌和真菌。通过平板对峙和牛津杯试验发现4 株菌株之间不存在拮抗作用,具有良好的兼容性,适合进行菌株组合。利用1.2.3 中的方法,分别制备细菌细胞悬浮液和真菌孢子悬浮液,并将4 株秸秆降解菌进行全组合复配:按照各株菌等频率出现和等比例混合的原则,构建菌株丰富度(多样性水平)为1、2、3、4 的复合菌系。复合菌系丰富度由低到高的组合数分别为4、6、4、1,共得到15 种组合,每个组合重复3 次并依次编号(表1)。复合菌系对水稻秸秆的相对降解率和3 种纤维素酶活性的检测分别参考1.2.3 和1.2.4 中描述的方法。
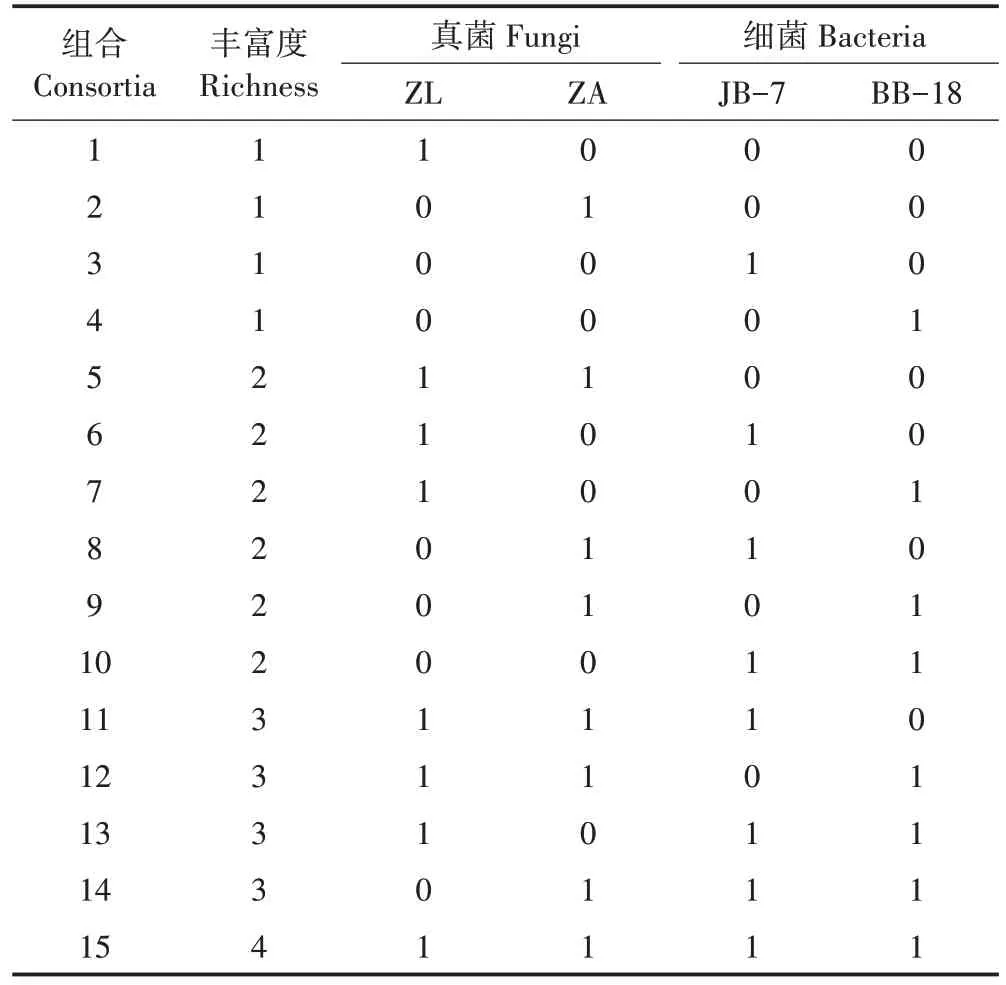
表1 秸秆降解细菌−真菌的全组合复配Table 1 Compositions of straw degradation communities using bacterial and fungal strains
1.2.6 数据分析
所有结果采用R 4.0(https://www.r−project.org)分别进行数据处理和统计分析,通过tidyverse、reshape2和tidyr 包进行数据转化处理,采用stats 和ggpubr 包进行Z−score 标准化和统计分析,利用ggplot2 进行数据可视化。统计分析采用单因素方差分析(ANOVA)和最小显著性差异法配对比较(LSD 检验),显著性水平为0.05。相关性分析采用线性回归(y=ax+b)评价检测指标随菌株多样性或秸秆相对降解率改变的变化趋势(a为斜率,b为截距),采用多元回归(y=ax1+bx2+cx3+d)评价不同酶活性对秸秆降解率的影响,进而明确发挥关键作用的酶。
2 结果与分析
2.1 水稻秸秆降解细菌和真菌的筛选与鉴定
以水稻秸秆粉为唯一碳源,通过室内富集、分离、纯化,从腐解的水稻秸秆中筛选获得8 株水稻秸秆降解菌(图1),包括4 株细菌(BB4、BB18、JB7 和JF7)和4 株真菌(JH、ZA、ZB 和ZL)。16S rRNA 基因与ITS 序列测序和系统发育分析结果(图1A)表明,细菌分离株 BB4、BB18、JB7 和 JF7 分别与类香味菌属(Myroi⁃des)、普罗威登斯菌属(Providencia)、短小芽孢杆菌(Lysinibacillus)和丛毛单胞菌属(Comamonas)基因的相似度超过99.5%,命名为玄武类香味菌(Myroides odoratimimusBB4)、雷氏普罗威登斯菌(Providencia rettgeriBB18)、球形赖氨酸芽孢杆菌(Lysinibacillus sphaericusJB7)和睾丸酮丛毛单胞菌(Comamonas tes⁃tosteroniJF7)。真菌分离株JH 和ZL 与曲霉属真菌ITS 序列高度同源,相似度超过99.6%,分别命名为黑曲霉(Aspergillus nigerJH)和烟曲霉(Aspergillus fumig⁃atusZL);真菌分离株ZA 和ZB 与青霉属(Penicillium)和漆斑霉属(Striaticonidium)真菌ITS 序列相似度超过99.8%,分别命名为草酸青霉(Penicillium oxalicumZA)和包围漆斑菌(Striaticonidium cinctumZB)。
2.2 细菌和真菌分离菌株的秸秆降解效果和滤纸酶活性评价
水稻秸秆降解试验表明,细菌、真菌不同分离株之间的降解能力存在显著差异,且真菌降解水稻秸秆的能力远高于细菌(图1B)。不同真菌分离株对水稻秸秆的降解率之间的差异显著(F3,8=177.0,P<0.000 1),ZA 最高(24.18%±1.71%),ZL 较高(17.39%±1.00%),JH次之(9.51%±1.01%),ZB最低(4.60%±0.32%)。细菌分离株的水稻秸秆降解率之间同样存在显著差异(F3,8=4.3,P<0.044 2),以 JB7(1.33%±0.40%)和BB18(1.28%±0.33%)最高,且二者之间差异不显著(P>0.05),BB4 略低(0.96%±0.09%),且与BB18 差异不显著(P>0.05),JF7最低(0.57%±0.26%)。
滤纸酶活性检测结果表明,不同细菌和真菌分离株滤纸酶活性差异显著,其中真菌分离株的滤纸酶活性较高(图1C)。真菌分离株之间的滤纸酶活性差异较大(F3,8=189.0,P<0.000 1),而细菌分类株之间的滤纸酶活性差异相对较小(F3,8=15.0,P<0.001)。其中,真菌分离株ZA 的滤纸酶活性最高,为(12.73±1.26)U·mL−1,ZL 次之,为(9.99±0.42)U·mL−1,二者间差异显著(P<0.05);ZB 和 JH 最低,且二者间差异不显著(P>0.05),分别为(1.66±0.42)U·mL−1和(1.44±0.41)U·mL−1。细菌分离株 JB7 和 BB18 的滤纸酶活性最高,分别为(1.63±0.02)U·mL−1和(1.53±0.01)U·mL−1,二者间差异不显著(P>0.05);BB4 略低,为(1.15±0.31)U·mL−1,JF7最差,为(0.85±0.05)U·mL−1。综上,细菌分离株雷氏普罗威登斯菌BB18、球形赖氨酸芽孢杆菌JB7 和真菌分离株草酸青霉ZA、烟曲霉ZL 具有较强的水稻秸秆降解能力(图1B),以及较高的滤纸酶活性(图1C),因此选择上述4株秸秆降解菌进行细菌−真菌复合菌系的构建。
2.3 复合菌系的菌株多样性与秸秆降解效果和产酶活性之间的关系
4 株降解菌全组合复配的秸秆降解试验结果(图2)表明,复合菌系对水稻秸秆的降解率与降解菌多样性成正相关关系(y=0.35x−0.75,R2=0.562 4,P<0.000 1)。随降解菌种类的增加,复合菌系的相对降解率逐渐增大(F3,41=8.2,P=0.000 2)。复合菌系降解水稻秸秆的能力均优于单菌(12.06%±10.29%,P<0.05),其中以4株菌的组合对水稻秸秆的降解率最高(31.40%±1.01%,P<0.01),较单菌的平均降解率提高了1.6 倍(图2A)。2 株与3 株菌的组合降解水稻秸秆的能力相当(20.20%~21.03%,P>0.05),较单菌的降解率提高了67.50%~74.38%(图2A)。因此,复合菌系的水稻秸秆降解能力可能存在菌株多样性效应。
为探究复合菌系秸秆降解能力多样性效应形成的原因,考察了与3 种秸秆中木质纤维素降解过程相关的酶的活性。结果表明,复合菌系产滤纸酶、纤维素内切酶和木聚糖酶的活性也具有多样性效应,即3种酶的活性随复合菌系菌株多样性的增加而增加,但变化程度有所差异(图2)。随菌株多样性的增加,复合菌系木聚糖酶的活性增幅最大(y=8.99x−2.36,R2=0.504 6,P=0.000 4),滤纸酶的增幅次之(y=0.35x−0.76,R2=0.316 7,P=0.034 0),纤维素内切酶增幅最小(y=0.04x+0.03,R2=0.554 1,P<0.000 1)。不同多样性复合菌系之间的纤维素内切酶(F3,41=6.2,P=0.001 3)和木聚糖酶(F3,41=5.8,P=0.002 1)的活性存在显著差异,而滤纸酶活性的差异不显著(F3,41=1.8,P=0.166 0)。LSD 检验发现,3、4 种菌株组合的纤维素内切酶(164.59~189.03 U·mL−1)和木聚糖酶(2.55×103~2.90×103U·mL−1)的活性均较高,且均不存在显著差异(P>0.05),较单菌的平均酶活性分别提高了1.15~1.47 倍和2.67~3.17 倍;两两组合与单菌产纤维素内切酶(110.13 U·mL−1)和木聚糖酶(1.38×103U·mL−1)的活性均较低,且均不存在显著差异(P>0.05)。因此,秸秆降解能力多样性效应,可能是由复合菌系产酶活性多样性效应引起的。综上,4 株细菌−真菌组成的复合菌系对水稻秸秆的降解能力和总酶活力较强,命名为BF1。
2.4 复合菌系产纤维素酶活性与秸秆降解效果之间的关系
相关分析结果(图3)表明,复合菌系对水稻秸秆的降解能力与其产3 种纤维素酶的活性呈正相关关系,即酶活性越强,复合菌系对水稻秸秆的降解效果越好。其中,复合菌系的秸秆降解率随纤维素内切酶活性的增加,增长幅度最大(y=0.87x+8.54,R2=0.708 2,P<0.000 1);随滤纸酶活性的增加,增长幅度次之(y=0.53x+13.29,R2=0.536 6,P=0.000 2);随木聚糖酶活性的增加,增长幅度最小(y=0.22x+15.28,R2=0.504 6,P=0.000 4)。通过Z−score 标准化计算秸秆降解率与总酶活性(标准化后3 种酶活的总和)并进行相关分析发现,复合菌系的降解能力与总酶活性也呈正相关关系,即复合菌系秸秆降解效果随总酶活性的增加而显著增强(R2=0.587 3,P<0.000 1)。通过进一步多元回归分析发现,复合菌系的秸秆降解能力与3 种酶的活性密切相关(F3,41=18.1,R2=0.569 6,P<0.000 1),其中纤维素内切酶(F3,41=48.4,P<0.000 1)和木聚糖酶(F3,41=6.1,P<0.017 4)的活性对复合菌系降解水稻秸秆能力的贡献较大,而滤纸酶活性的贡献较小(F3,41=0.7,P=0.515 9)。上述结果说明,纤维素内切酶和木聚糖酶是复合菌系降解水稻秸秆的重要驱动力。
3 讨论
通过4 株秸秆降解菌的全组合复配,构建菌株多样性由低到高的15种细菌−真菌复合菌系,评价了菌株多样性对复合菌系降解水稻秸秆的效果,分析了3种与秸秆木质纤维降解相关的酶活性受到的影响,探究了复合菌系秸秆降解效果与产酶活性之间的关系。研究发现,细菌−真菌复合菌系对水稻秸秆的降解效果优于单一降解菌株,且复合菌系的秸秆降解能力随菌株多样性的增加而显著增强。这与其他复合菌系降解秸秆效果的研究结果相似[24-26],例如,江高飞等[13]通过对5 株真菌复配研究发现,复合菌系对玉米秸秆的降解效果优于单一菌株,且降解效果随降解真菌多样性的增加而提高,真菌复合菌系对秸秆的降解率也具有多样性效应。但也有研究与本研究结果不同,如李静等[25]利用15 株秸秆降解细菌进行复配研究,发现复合菌系降解玉米秸秆的能力并非随细菌种类的增加而增加,其中多样性为3 和5 的3 个组合的秸秆降解效果甚至低于链霉菌、不动杆菌、类芽孢杆菌和芽孢杆菌等降解菌单独降解玉米秸秆的能力;魏蔚等[26]将6 株降解真菌与蜂房芽孢杆菌复配成多样性为4 和7 的两种复合菌系,比较发现4 种降解菌组合降解小麦秸秆的能力显著高于7 种降解菌组合,但这两种组合产酶的能力尚不清楚。
细菌和真菌等微生物在秸秆降解过程的作用机理不同,因此能够通过协同作用完成秸秆的降解[6]。本研究发现细菌−真菌复合菌系的产酶活性具有多样性效应,增加菌株的种类可显著增强复合菌纤维素降解酶的活性,协同提高降解水稻秸秆的效果。同样,LU 等[27]发现15 株降解菌单独培养时的滤纸酶活性较低,通过不同组合方式复配后提高了降解滤纸的能力,其中6 株菌的组合降解率达26.3%。江高飞等[13]同样发现真菌复合菌系的酶活性也具有多样性效应,且滤纸酶和纤维素内切酶是复合菌株降解玉米秸秆的关键驱动因素。然而,本研究发现纤维素内切酶和木聚糖酶对复合菌系秸秆降解效果的贡献显著,这可能是玉米秸秆与水稻秸秆中纤维素种类和丰度的差异所致。本研究构建的复合菌系BF1对水稻秸秆的平均降解率为31.40%,较单菌的平均降解率提高了1.6 倍,纤维素酶活力提高了1~3 倍。张必周等[9]从GF 自然复合菌系(降解率为20.51%~34.05%)中筛选得到11 株降解菌进行组合,发现复合菌系的降解能力优于单菌,其中复合菌系G 的秸秆降解率为48.56%,较不加菌的对照提高了21.56%,酶活性提高了36.35%。诸葛荣夏[28]将6 株降解菌进行有效组合,发现复合菌系W−2 的降解率达32.51%,较单菌提高了2 倍,除木素过氧化物酶无明显差异外,其他纤维素酶均提高了2 倍以上。此外,优化复合菌系中菌株的比例能够提高复合菌系的降解效果,如枯草芽孢杆菌、产朊假丝酵母、植物乳杆菌、地衣芽孢杆菌的比例为 1∶1∶2∶1 时,细菌−真菌复合菌系产酶活性最大,降解效果最好[29]。因此,可通过调整菌株的比例和培养条件等,探索提高细菌−真菌复合菌系产酶活性和降解效果的最佳方法,为服务生产应用提供理论基础。此外,环境微生物种类复杂,建议菌株筛选鉴定、保藏管理和生产应用上应当兼顾菌种生物安全的问题。
4 结论
(1)本研究将筛选获得的秸秆降解能力较好的两株细菌和两株真菌,通过全组合的方式构建出不同组合的复合菌系,发现复合菌系的降解率和3 种纤维素酶活性均随菌株种类的增加而提高。
(2)明确了复合菌系产纤维素酶活性与秸秆降解效果之间的关系,发现纤维素内切酶和木聚糖酶是复合菌系降解水稻秸秆的主要驱动因素。
(3)筛选获得了一组具有潜在应用前景的细菌−真菌复合菌系BF1(由雷氏普罗威登斯菌BB18、球形赖氨酸芽孢杆菌JB7、草酸青霉ZA和烟曲霉ZL组成),其秸秆降解率较单菌提高了1.6倍,酶活性增加了1~3倍。在复合菌系BF1的实际应用中,还应考虑优化菌株的配比和剂量,以提高复合菌系的秸秆降解率。
