橡胶树魏克汉种质成龄树经预割诱导后次生乳管分化的比较
2021-11-08史敏晶张世鑫晁金泉吴绍华杨署光田维敏
史敏晶 张世鑫 晁金泉 吴绍华 杨署光 田维敏

摘 要:巴西橡膠树树干中次生乳管列的多少与胶乳产量密切正相关,而割胶伤害能诱导次生乳管分化。分析不同魏克汉种质成龄树在预割伤害后树皮次生乳管的分化,不仅为不同种质鉴定和评价提供依据,也为成龄大树割胶后次生乳管分化能力提供预测。采用碘-溴染色方法和石蜡切片技术,对不同种质材料新分化的次生乳管进行了统计分析。研究结果表明,选取的110份魏克汉种质预割后新分化的次生乳管列数不同,具有明显的种质特征。根据分化次生乳管列数的多少,将预割诱导乳管分化的级别由低到高分为1、2、3、4级。在110份种质中,对应1级32份、2级49份、3级24份和4级5份的种质,所占比例分别为29.09%、44.55%、21.82%和4.55%,可见,分化能力强的种质占比较低,大部分种质为2级乳管分化。国外直接引种和国内自主选育的种质在预割诱导分化能力上也存在差异。在110份种质中,国外引种58份种质中对应分化能力1、2、3、4级的种质占比分别为29.31%、44.83%、22.41%以及3.45%;国内选育的52份种质中对应1、2、3、4级的种质占比分别为28.85%、44.23%、21.15%和5.77%;可见,国内选育种质受预割诱导乳管分化的能力优于国外引种的种质,尤其是4级分化能力的种质比例明显高于国外引种的种质。不同系列来源的种质乳管分化能力也存在差异,其中,国外PB、RRIM和RRIC系列和国内的大岭、海垦和大丰系列表现出较强的乳管分化能力。比较分析2个杂交组合后代的乳管分化情况发现,PB86×PR107的4份子代中次生乳管有3份为3级,1份为4级,后代表现出较强的乳管分化能力;而RRIM600×PR107的6份子代中1级2份,2级2份,3级2份,可见其后代乳管分化能力略差。PB86和PR107后代的乳管分化表现出较强的双超亲现象,可作为优良的育种亲本。通过预割判断乳管分化能力不仅为魏克汉种质鉴定和评价奠定基础,也为橡胶树高产育种以及早熟、晚熟品种的早期预测提供有效指标。
关键词:橡胶树;魏克汉种质;预割;次生乳管分化;早期预测
中图分类号:S794.1 文献标识码:A
Comparision of Secondary Laticifer Differentiation of Mature Tree Induced by Pre-tapping in Wickham Germplasm of Rubber Tree
SHI Minjing, ZHANG Shixin, CHAO Jinquan, WU Shaohua, YANG Shuguang, TIAN Weimin*
Rubber Research Institute, Chinese Academy of Tropical Agricultural Sciences / Key Laboratory of Biology and Genetic Resources of Rubber Tree, Ministry of Agriculture and Rural Affairs / State Key Laboratory Incubation Base for Cultivation and Physiology of Tropical Crops, Haikou, Hainan 571101, China
Abstract: The number of secondary laticifer rows in the trunk of Hevea brasiliensis has a significantly positive correlation with the latex yield, and the differentiation of the secondary laticifer can be induced by tapping wound in mature tree trunk. In the study, the secondary laticifer differentiation of different Wickham germplasm mature trees induced by pre-tapping were studied, which would provide a basis for the identification and evaluation of germplasm resources, and the prediction of the differentiation ability of secondary laticifer of mature trees after tapping. Fresh bark samples were dyed by iodine-bromine reagent and embedded in paraffin. The differentiation of the secondary laticifers were observed with an optical microscope and the number of secondary laticifer rows were recorded for statistical analysis. There was a significant difference of secondary laticifer differentiation among the 110 different accessions of Wickham germplasms induced by pre-tapping. According to the number of secondary laticifer rows after pre-tapping, the differentiation ability of secondary laticifer from low to high was roughly classified into 4 grades:1, 2, 3 and 4 grades. Based on these criteria, there were 32, 49, 24 and 5 accessions corresponding to grade 1, 2, 3 and 4, with the percentage of 29%, 45%, 22% and 4% among 110 accessions respectively. Therefore, the secondary laticifer differentiation ability most of the Wickham germplasms was relatively low level of grade 2. Further, there was also obvious difference in the differentiation ability of secondary laticifer between the germplasms introduced directly from abroad and those selected from domestic germplasms. Among 58 germplasms introduced from abroad, the percentage of accessions corresponding to grade 1, 2, 3, and 4 was 29.31%, 44.83%, 22.41%, and 3.45%, respectively. Among 52 germplasm selected from China, the percentage of accessions corresponding to the grade 1, 2, 3, and 4 was 28.85%, 44.23%, 21.15%, and 5.77% respectively. So, the domestic germplasms generally were superior to the abroad ones, in which especially the proportion of germplasms with 4-grade was significantly higher than that of foreign germplasms. In different breeding series, the PB, RRIM and RRIC series from abroad and the Daling, Haiken, and Dafeng series from domestic germplasms showed a better differentiation effect. To analyze the progenies of two hybrid combinations, the results showed that generations of PB86 × PR107 had strong differentiation ability, in which 3 accessions were grade 3 and 1 accession was grade 4. In comparison, the generations of RRIM600 × PR107 showed lower differentiation ability with 2 accessions from grade 1 to grade 3 respectively. PB86 and PR107 progenies showed strong double transgressive phenomenon in the aspect of laticifer differentiation and could be chose as superior breeding parents. The ability of secondary laticifer differentiation induced by pre-tapping could be used as an effective indicator for early prediction of high-yield and peak yielding period, and the results also provided the basis for identification and evaluation of the germplasm resources for rubber tree.
Keywords: Hevea brasiliensis; Wickham germplasm; pre-tapping; secondary laticifer differentiation; early prediction
DOI: 10.3969/j.issn.1000-2561.2021.09.016
巴西橡胶树(以下简称橡胶树)是人工栽培最重要的一种产胶植物,其提供了世界所需98%以上的天然橡胶[1]。作为原产南美亚马孙河流域的热带雨林树种,橡胶树主要生长在南纬10至北纬15之间的区域,形成了典型的喜高温、多雨和静风等的生态习性[2]。与东南亚主要植胶国家相比,我国属于国际公认的非传统植胶区,适于植胶的国土面积有限[2-3],因此,如何提高单位面积产量一直是我国天然橡胶产业面临的主要问题,如何有效筛选高产、稳产的种质,一直是橡胶树育种研究的重点[4-5]。当前选育种的主要方法是试割3~4龄幼树后统计橡胶树胶乳产量,根据其高低来选择优良单株种质,但胶乳产量是一个典型的综合数量性状,受多种因素的影响,并不能完全真实的反应种质自身产胶的潜力。另外,试割法本身也存在诸多缺陷[1]。如何寻找有效指标提高育种效率,减少具有高产潜力的种质漏选几率并缩短育种年限,一直是橡胶树育种中亟待解决的问题。
橡胶树中胶乳产生和贮藏在一种特化的组织——乳管中,橡胶树树干中的次生乳管是由维管形成层中的纺锤状原始细胞切向分裂形成的,与形成层呈同心圆状排列[1],生产中割胶获取的胶乳都来自于成龄大树的次生乳管,其数量的多少与天然橡胶的产量显著正相关[6]。目前已有研究表明,乳管分化能力、2次割胶之间的胶乳再生能力以及排胶时间长短是决定橡胶树产胶能力的3个最核心的因素[7],可以作为产量的构成性状对种质的产胶潜力进行筛选。其中,橡胶树树干中的次生乳管分化能力,最终反映出乳管列数量的多少是当前最受关注的因素[8-15]。研究表明,割胶造成的机械伤害和排胶都能诱导成龄树树干次生乳管分化,并且排胶起主要作用[8]。对7~8龄橡胶树预割,既造成机械伤害也导致了排胶,经证明也能诱导乳管分化[9]。预割通常是针对即将正式开割的大树,分析不同成龄树种质在预割伤害后次生乳管的分化特征能有效预测正式割胶后橡胶树的乳管分化情况,为更早、更有效地筛选出具有高产潜能的种质提供依据。
魏克汉种质普遍产量高、综合性状好,是目前橡胶树生产栽培中的核心种质,是我国最重要的种质资源[16]。分析其成龄大树在预割伤害后次生乳管分化的特征,不仅为鉴定和评价魏克汉种质提供依据,也为成龄大树割胶后乳管分化的能力提供早期预测,这对于缩短橡胶树产量育种时间和提高产量效率具有重要的应用价值。
1 材料与方法
1.1 材料
1.1.1 试验材料 选取种植于中国热带农业科学院儋州试验场三队(海南儋州宝岛新村)8年生
魏克汉种质110份(国外直接引种58份,我国自主选育52份,见表1),当年8月于距地面高2 m处以1/4树围开割线并同时采集割线上垂线处树皮一份作为对照。按照3天1刀的割制,阴刀连续割胶10刀,然后停割1个月,在割线上垂线位置再采集树皮一份。树皮经80%酒精固定后,碘溴染色处理,制备石蜡切片。
1.1.2 主要试剂和仪器 制备石蜡切片所需的乙醇、正丁醇、冰醋酸、二甲苯为国产分析纯,石蜡为生物组织切片专用。切片机为Leica RM2235型旋转切片机,成像显微镜为德国Leica DMLB型光学显微镜。
1.2 方法
1.2.1 光学显微切片的制备 材料采集后分割,并经80%(V/V)乙醇固定24 h,乙醇梯度脱水,碘-溴冰醋酸试剂60 ℃处理48 h,乙醇脱水后经正丁醇透明,石蜡包埋,具体方法参见史自强等[17]方法并稍作修改。树皮横切,切片厚度15 m,二甲苯脱蜡后固绿染色1 h,中性树胶封片。
1.2.2 数据统计分析 光学显微镜下观察横切面切片并成像。光镜下统计每份切片材料新生的次生乳管分化列数情况,采用Excel 2003制作图表。
2 结果与分析
2.1 预割诱导橡胶树魏克汉种质次生乳管的分化
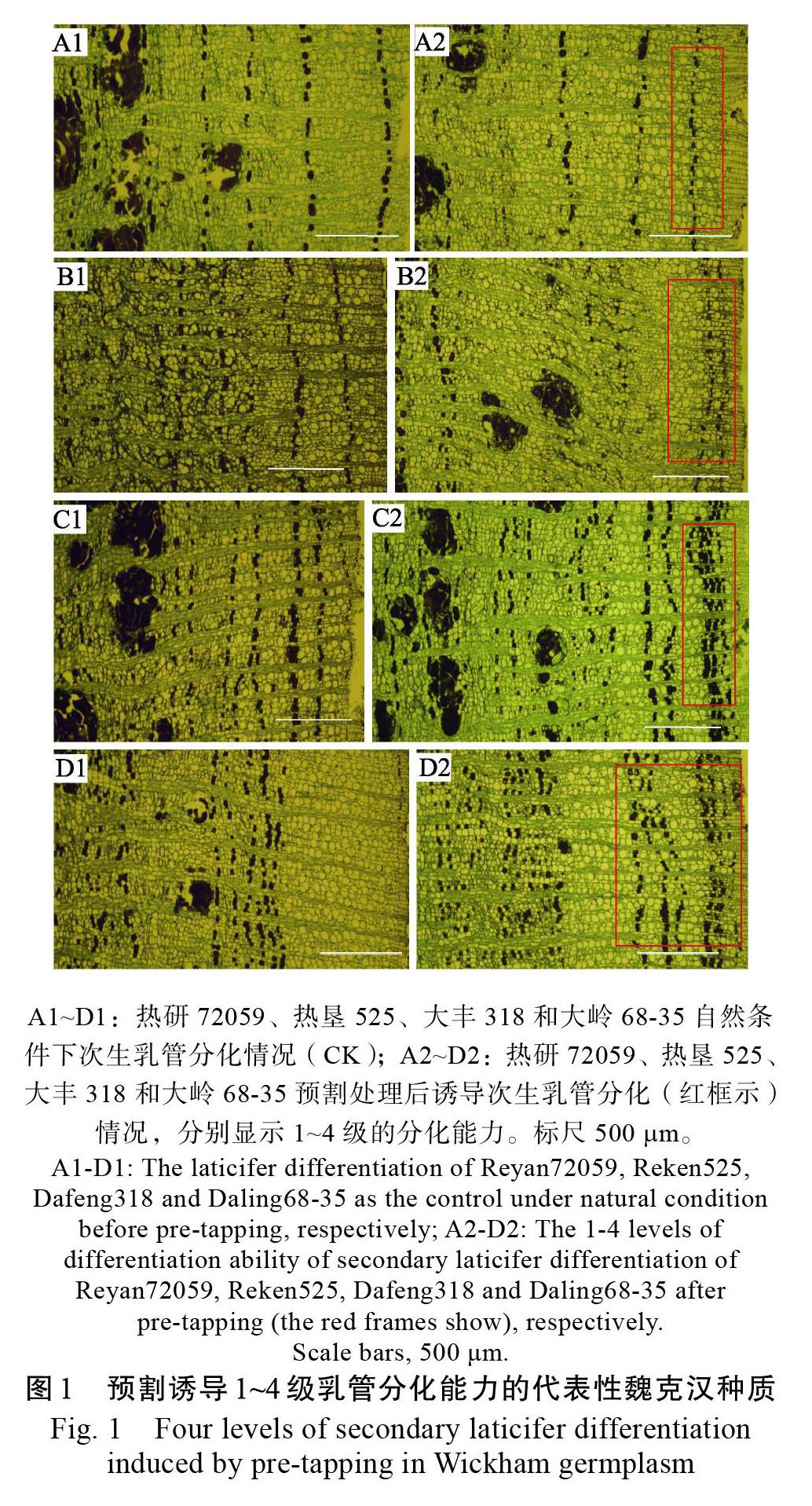
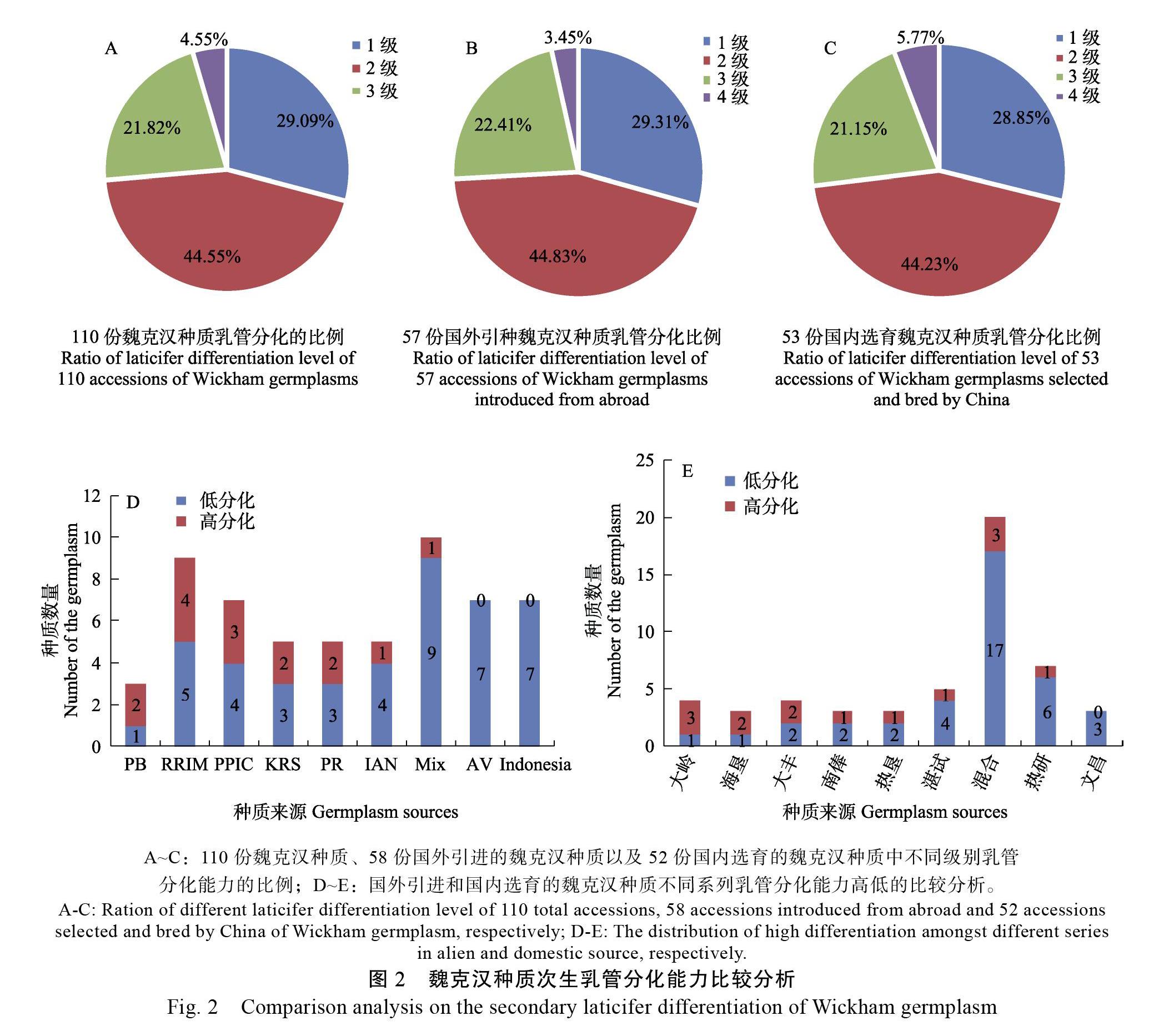
对110份8年生的魏克汉种质大树的树干进行预割处理,处理采样与第一刀预割割胶伤害的时间间隔约为2个月,根据次生乳管形成到脱离有输导功能韧皮部至少1 a的时间,可知新形成的乳管列距形成层较近,而预割前自然形成的乳管列相对远离形成层,在新乳管列的外侧。新的乳管列之间间隔薄壁细胞的层数少,有密集趋势,而自然形成的乳管列之间间隔的细胞层数较多,间距较远。研究表明,不同的魏克汉种质在预割期间新形成的次生乳管列数不同,具有明显的种质特征。根据新形成次生乳管列数的多少,将成龄树被诱导的乳管分化能力分为1、2、3和4级,分别对应于新分化乳管列数少于2列、2~3列之间(含2列)、3~4列之间(含3列)和4列及以上(图1)。根据这一分级标准,在110份魏克汉种质中,1级分化的种质有32份,占比为29.09%;2级分化的种质有49份,占比为44.55%,二者合计为73.64%,可见大部分种质的乳管分化能力处在较弱的1级和2级。分化出3列及以上次生乳管的被认为具有较好的乳管分化能力,分别对应于3级和4级分化,其中3级分化的种质有24份,占比为21.82%;4级分化的种质仅有5份,占比为4.55%,可见分化能力强的种质总体占比较低(图2A)。
根据魏克汉种质的来源,可将之分为国外直接引进和国内自主选育两部分。国外引进种质有58份,对应分化能力为1级的种质为17份,占比为29.31%,代表种质有PR107、RRIM600、IAN2887等;分化能力2级的种质数量明显增加至26份,占比高达44.83%,代表种质有GT1、PB86、RRIC110等;分化能力3级的种质数量减少至13份,占比為22.41%,代表种质有IAN2904、PB260、RRIM725和泰1等;具有4级分化能力的种质数量极少,仅2份,分别为KRS33和RRIC130,占比低至3.45%(图2B)。国外引种种质根据其来源系列的不同,乳管分化能力高低也有区别。将分化能力1、2级的合并记为低分化类型,3、4级的合并记为高分化类型,则这些系列中具有高分化能力的比例由高到低分别为PB系列、RRIM系列、RRIC系列、KRS系列、PR系列、IAN系列、其他合并系列(少于3份种质的系列合并统计)、AV系列以及Indonesia系列(图2D),结果与生产中PB系列、RRIM系列种植比较广泛、有良好的产量表现相对应。
国内选育的种质共52份,对应次生乳管分化能力为1级的种质有15份,占比为28.85%,代表种质有热研72059、红星1、大丰117等;分化能力2级的种质数为23份,占比为44.23%,代表种质有保亭155、桂研74-1、热垦525等;分化能力3级的种质有11份,占比为21.15% ,代表种质有大丰318、热研7-33-97、热垦165、海垦2、海垦6等;具有4级分化能力的种质数量也是最少的,仅3份,分别为大丰95、大岭68-35、南俸37,但占比为5.77%,明显高于国外引进的种质(图2C)。可见,国内选育的种质整体分化能力优于国外引种的种质。我国自主选育的种质根据系列不同预割诱导分化次生乳管的能力也有差异,由高到低分别为大岭系列、海垦系列、大丰系列、南俸系列、热垦系列、湛试系列、其他、热研系列以及文昌系列(图2E)。
比较分析预割前后树皮中的次生乳管列,包括有输导功能韧皮部、黄皮层和砂皮内层中的成列的有效乳管,发现自然条件下有效乳管列少、乳管列间距大的种质在预割诱导后新形成的乳管列普遍数量少,表明分化能力较弱;而自然条件下有效次生乳管列数量多、乳管列之间间距小的种质在预割诱导后通常新形成的乳管列数量也较多,表现出较强的分化能力。
2.2 预割诱导不同杂交组合后代的次生乳管分化情况
针对2组来自不同橡胶树亲本和杂交后代预割后次生乳管的分化能力进行了分析。来自PB86×PR107的4份子代分别是大丰95、海垦2、海垦6和大豐318。PB86和PR107作为亲本均是从国外引进的初生代无性系,定植8年的植株尚处于成龄幼树阶段,无论是自然条件下还是预割伤害诱导,次生乳管的分化能力都较差,分别评为2级和1级(图3A1~A2、B1~B2);但在其杂交子代中出现了3份分化能力为3级的种质,分别为海垦2、海垦6和大丰318,1份分化能力为4级的种质,即大丰95,可见,该组合的后代表现出较强的次生乳管诱导分化能力。
来自RRIM600×PR107的6份子代中次生乳管分化能力1级的有2份,分别为热研72059和热研7-18-55;分化能力2级的有2份,为文昌11和保亭155;分化能力3级也有2份,分别为热研7-33-97和大岭21-65。作为亲本,RRIM600和PR107的乳管分化能力均为较差的1级(图3C1~C2、B1~B2),而其后代种质的次生乳管诱导分化差异比较明显,从1级到3级都具有。
由此可见,即使橡胶树育种亲本的乳管分化能力不强,但后代仍有可能出现较强的乳管分化能力这一性状。培育优良品种的主要关键之处是选择优良的亲本,从优化乳管分化能力方面,PB86×PR107组合可能优于RRIM600×PR107组合。
3 讨论
橡胶树育种周期长达30多年,需耗费大量的人力、物力,占地面积极大,并且一直以来都是以胶乳产量这一数量性状来筛选种质的优劣,而不是按照产胶量的相关构成性状选配杂交组合,导致真正互补的性状极少,杂交后代很难大幅度超亲[4,18]。随着天然橡胶产业研究的推进,与胶乳产量密切相关的因子主要被分解为3个,其中,次生乳管作为天然橡胶产生和贮存的主要组织,其数量的多少是决定天然橡胶产量的最关键因子之一[7]。因此,选育具有更多的有效次生乳管列的品种是当前橡胶树育种工作中的重点,而如何在早期预判一个种质的次生乳管分化能力强弱,进而筛选出具有高产潜能的种质对于缩短橡胶树选育种年限、提高育种效率具有重要的意义。
为建立起一套有效、准确的次生乳管分化能力早期鉴定方法,田维敏团队对处在不同年龄段的橡胶树种质进行了次生乳管分化能力的分析,其中包括机械伤害诱导幼嫩萌条分化次生乳管[19-21],模拟割胶2年生幼树[13]、试割诱导3~4龄的幼龄树分化次生乳管[22],结果都表明,伤害诱导次生乳管分化的能力在不同种质中存在明显差异,是橡胶树种质本身一种稳定的遗传特性。由于橡胶树产胶存在早熟、晚熟等特性,为更好的了解不同种质在不同年龄段割胶伤害后乳管分化的特征,对与常规割胶树状态最为接近的8龄树进行预割,分析预割诱导次生乳管分化的能力,可对橡胶树天然橡胶生产中在不断的割胶伤害下次生乳管的分化能力进行科学的预判,同时进一步完善乳管分化能力的早期鉴定方法。
本研究对110份魏克汉种质8龄大树进行了预割,发现不同的种质对预割伤害的反应不同,具有种质特征。从伤害开始到取材的2个月时间内,大部分种质只能分化出1~2列的次生乳管,占所有种质的70%以上;能分化出3列及以上乳管的种质占比不足30%。根据分化能力高低,我们将之划分为4个等级,其中分化能力最强的4级占比最低,不足5%;分化能力2级的占比最高,接近50%,可见在乳管分化这一性状上表现很好的种质只有少数。比较国外直接引进和国内自主选育以及不同系列来源的种质,可知国内选育的种质分化能力普遍强于国外引进的,而其中又以海南选育的大岭系列、海垦系列、大丰系列表现最好,这可能与国内尤其是海南选育种主要以高产为定向目标有关,长期的高产选育对于高乳管分化能力有一定的选择作用。
大丰95、大岭68-35以及南俸37在预割时诱导乳管分化的能力强,为4级,在生产中,尤其在海南省是分别作为大规模和中规模推广级种植的品种[4,23];但PR107、RRIM600以及热研72059等品种预割时表现差,乳管分化能力仅1级,但在海南省同样为大规模以及中规模推广等级。这可能与被广泛推广的品种往往是有较好的综合产量表现这一要求有关,在生产中,除考虑乳管数量多少外,也会考虑抗风、抗寒以及耐死皮等相关性状[24-25]。PR107和热研72059等品种属于晚熟品种[26-27],即在最初几年割胶时产量不高,但随着割龄增加,到中晚期产量大幅提升,分析认为这种晚熟特性可能与这些品系的大树在割胶初期次生乳管受伤害诱导增加并不明显有关,即如本研究中预割的效果,但割胶中、后期有效乳管数量增多,且具有抗风及其他良好的性状表现[24,28],因此这些品种也能成为高产品种而被推广种植。
比较史敏晶等[21]利用机械伤害诱导幼嫩萌条的次生乳管分化系统和本研究中8龄树预割伤害诱导次生乳管分化的系统发现,大丰95、大岭68-35、热研7-33-97、海垦6和泰1等很多品种在2种系统中都有优良的表现;PR107、RRIM600以及PB86在萌条诱导中表现出良好的次生乳管分化能力,但在大树预割中却分化能力差,二者呈现出相反的趋势。可见,2个不同的实验系统在预测不同树龄种质的次生乳管分化能力方面既有一致性,也有差异。在2个系统中都表现出高乳管分化能力的种质,基本都对应于生产中的早熟、高產品种;在苗期具有高乳管分化能力、在预割分化能力一般的如PR107等种质,认为这种类型的种质具备较强的乳管分化能力,在幼苗时期这种潜力比较敏感容易体现出来,但大树在割胶初期对诱导的反应不强,所以表现出高产、晚熟的特性。综合分析幼苗期和成龄期橡胶树对伤害的反应,能够更全面地反映不同的种质次生乳管分化潜力以及树龄对次生乳管分化能力的影响,为生产中判断高产以及早熟、晚熟特性提供参考依据。
对2个杂家系后代的乳管分化这一性状的分析可知,在预割时乳管分化能力差的亲本同样可以获得乳管分化能力强的子代,关键是看亲本之间是否具有较好的互补性。所以,单从乳管分化能力这一性状来看,培育具有高乳管分化能力的后代其亲本的选择是比较广泛的,可以是本身分化能力并不很强的种质。PB86×PR107组合中,后代表现出较明显的超亲趋势,且PB86具有多花多果的特质,而PR107雌花发育不良,所以二者可以作为优良的育种亲本,筛选出乳管分化能力强的后代。利用已经具有很强的乳管分化能力的种质作为亲本是否可以获得更强分化能力的后代在育种工作中也值得进一步尝试研究。
4 结论
不同魏克汉种质成龄树对预割诱导分化次生乳管的反应不同,具有明显的种质特征。国内自主选育的种质被诱导分化次生乳管的能力整体高于从国外直接引进的种质,尤其是次生乳管分化能力4级的种质国内占比明显高于国外引种的。在不同系列中,国外的PB系列、RRIM系列、RRIC系列和国内的大岭系列、海垦系列、大丰系列表现出较强的乳管分化能力,通常对应于生产中的高产品种。PB86和PR107作为亲本,其后代在乳管分化这一性状上表现出较强的双超亲现象,可以作为优良的育种亲本。本研究不仅为魏克汉种质鉴定和评价奠定基础,也为橡胶树高产育种以及早、晚熟品种的早期选择提供一个有效指标。
参考文献
[1] 田维敏, 史敏晶, 谭海燕, 等. 橡胶树树皮结构与发育[M]. 北京: 科学出版社, 2015.
[2] 何 康, 黄宗道. 热带北缘橡胶树栽培[M]. 广东: 广东科技出版社, 1987.
[3] 李 达, 张绍文. 天然橡胶产业发展政策分析: 红线与红利[J]. 林业经济问题, 2020, 40(2): 208-215.
[4] 黄华孙, 吴云通. 中国橡胶树育种五十年[M]. 北京: 中国农业出版社, 2005.
[5] 吴春太, 李维国, 黄华孙. 近年来国内外橡胶树种质资源与育种方法研究新进展[J]. 西北林学院学报, 2013, 28(2): 118-124, 146.
[6] Gomez J B. Anatomy of Hevea and its influence on latex production[J]. Kula Lumper: Malaysia Rubber Research and Development Board, 1982.
[7] 郝秉中, 吴继林. 巴西橡胶树乳管生物学与胶乳生产[J]. 热带作物学报, 2004, 25(4): 1-7.
[8] 郝秉中, 吴继林. 创伤(割胶)对乳管分化的影响[J]. Journal of Integrative Plant Biology, 1982(4): 96-99, 116.
[9] 于俊红, 杨署光, 黄绵佳, 等. 季节、采胶和外源茉莉酸对成龄树橡胶树乳管分化的影响[J]. 热带作物学报, 2007, 28(4): 1-5.
[10] 张世鑫, 刘世彪, 田维敏. 形成层活动对机械伤害诱导巴西橡胶树次生乳管分化的影响[J]. 热带作物学报, 2011, 32(6): 1037-1041.
[11] 曾 霞, 李维国, 高新生, 等. 巴西橡胶树选育种研究现状、趋势及我国的研究策略[J]. 热带农业科学, 2013, 33(6): 30-36.
[12] Tian W M, Yang S G, Shi M J, et al. Mechanical wounding-induced laticifer differentiation in rubber tree: an indicative role of dehydration, hydrogen peroxide, and jasmonates[J]. Journal of Plant Physiology, 2015, 182: 95-103.
[13] 张世鑫, 刘宁涛, 杨署光, 等. 巴西橡胶树次生乳管分化能力的早期评价方法研究[J]. 热带作物学报, 2018, 39(7): 1266-1275.
[14] Wu S, Zhang S, Chao J, et al. Transcriptome analysis of the signaling networks in coronatine-induced secondary laticifer differentiation from vascular cambia in rubber trees[J]. Scientific reports, 2016(6): 36384.
[15] Chao J Q, Zhao Y, Jin J, et al. Genome-wide identification and characterization of the JAZ gene family in rubber tree (Hevea brasiliensis)[J]. Frontiers in Genetics, 2019,10: 372. doi: 10.3389/fgene.2019.00372.
[16] Clement-Demange A, Priyadarshan P M, Thuy Hoa T T, et al. Hevea rubber breeding and genetics[J]. Plant Breeding Reviews, 2007, 29(6): 177-279.
[17] 史自强, 胡正海.植物含橡胶组织的制片方法[J]. Journal of Integrative Plant Biology, 1965, (2): 179-183.
[18] 林位夫. 作物次生产量特性及其利用对策探讨[J]. 热带作物学报, 2012, 33(4): 602-604.
[19] Tian W M, Shi M J, Yu F Y, et al. Localized effects of mechanical wounding and exogenous jasmonic acid on the induction of secondary laticifer differentiation in relation to the distribution of jasmonic acid in Hevea brasiliensis[J]. Acta Botanica Sinica, 2003, 45(11): 1366-1372.
[20] Yu J H, Zeng X, Yang S G, et al. Relationship between rate of laticifer differentiation, number of laticifer rows and rubber yield among 1981 IRRDB germplasm collections of Hevea brasiliensis[J]. Journal of Rubber Research, 2008, 11(1): 43-45.
[21] 史敏晶, 楊署光, 张世鑫, 等. 橡胶树魏克汉种质萌条机械伤害诱导的次生乳管分化特征[J]. 林业科学, 2020, 56(3): 28-37.
[22] Chen Y, Gao X, Zhang X, et al. Relationship between the number of tapping-induced secondary laticifer lines and rubber yield among hevea germplasm[J]. Frontiers of Agricultural Science and Engineering, 2016, 3(4): 363-367.?
[23] 刘作基, 马 伟, 谭振强, 等. 橡胶树大规模推广级无性系大丰95的研究[J]. 热带作物学报, 1996, 17(2): 19-24.
[24] 王祥军, 张源源, 张华林, 等. 巴西橡胶树抗风研究进展[J]. 热带农业科学, 2015, 35(3): 88-93.
[25] 李小琴, 赵 祺, 者国雄, 等. 橡胶树魏克汉种质资源抗寒性研究[J]. 热带农业科技, 2019, 42(1): 11-14, 29.
[26] 黄华孙, 方家林, 卓书蝉, 等. 橡胶树优良品种热研7-20-59的选育[J]. 热带作物学报, 2000, 21(2): 1-6.
[27] 吴 明, 刘实忠, 杨文凤, 等. 橡胶树热研7-20-59、PR107、RRIM600品种生理特性比较[J]. 安徽农业科学, 2013, 41(8): 3465-3467.
[28] 张晓飞, 胡彦师, 黄华孙, 等. 橡胶树优良亲本PR107的研究与利用[J]. 热带农业科学, 2015, 35(4): 39-44.
责任编辑:白 净
