基于ISSR标记的建兰种质资源遗传多样性分析
2021-11-08王宏利卜朝阳曾艳华龙蔷宇
王宏利 卜朝阳 曾艳华 龙蔷宇
摘 要:为揭示建兰种质间的亲缘关系,促进建兰种质资源的有效保护和利用。本研究利用ISSR标记分析了源于中国各地的39份建兰品种,并采用UPGMA方法进行了聚类分析。结果表明:6个ISSR引物对39份建兰品种的基因组DNA进行扩增,共扩増出64条清晰条带,其中57条为多态性条带。平均每条引物检测到的条带数为11条,多态性条带10条,多态性比率为89.88%。通过UPGMA法进行聚类分析表明供试材料的遗传相似系数在0.500~0.953之间,供试材料被分为6群集,说明ISSR标记在建兰的品种间具有丰富的多态性,能够很好地揭示建兰品种间的亲缘关系。本研究结果可为建兰种质资源品种鉴定及杂交育种等提供理论依据。
关键词:建兰;ISSR;遗传多样性分析;聚类分析
中图分类号:S682.31 文献标识码:A
Study on Genetic Diversity of Cymbidium Ensifolium Germplasm Based on ISSR Marker
WANG Hongli, BU Chaoyang*, ZENG Yanhua*, LONG Qiangyu
Flower Research Institute, Guangxi Zhuang Autonomous Region Academy of Agricultural Sciences, Nanning, Guangxi 530007, China
Abstract: To reveal the genetic relationship between C. ensifolium germplasm, promote the effective protection and utilization of C. ensifolium germplasm resources. In this study, ISSR markers were used to analyze 39 C. ensifolium varieties originating from various parts of China, and cluster analysis was performed using UPGMA method. The results showed that 6 ISSR primers amplified the genomic DNA of 39 C. ensifolium varieties, and a total of 64 clear bands were amplified, including 57 polymorphic bands. The average number of bands detected per primer was 11, polymorphic bands were 10 and the polymorphic ratio was 89.88%. Cluster analysis by UPGMA method showed that the genetic similarity coefficient of the test materials was between 0.500–0.953, and the test materials were divided into 6 clusters, indicating that ISSR markers are rich in polymorphism in C. ensifolium varieties, which can Reveal the relationship between C. ensifolium varieties. The results of this study can provide a basis for the identification and cross breeding of C. ensifolium germplasm resources.
Keywords: Cymbidium ensifolium; ISSR; genetic diversity analysis; cluster analysis
DOI: 10.3969/j.issn.1000-2561.2021.09.012
建蘭[Cymbidium ensifolium(L.) Sw.]属于国兰的一大种群。花浅黄绿色,芳香。与其他种类国兰明显区别是,花期较长且可多次开花,故又名四季兰。常见栽培品种包括金丝马尾、凤尾素、小桃红、龙岩素、铁骨素、君荷、朝阳三星蝶等。喜温暖湿润和半阴环境,耐寒性差,越冬温度不低于3 ℃,怕强光直射,不耐水涝和干旱,宜疏松肥沃和排水良好的腐叶上。建兰不仅观赏价值高,还可以入药使用,有极高的经济价值。
由于兰属植物品类繁多,种间和品种间的杂交严重,导致遗传背景错综复杂,谱系亲缘关系模糊不清,严重影响到兰属种质鉴定、品种改良和新品种的培育与开发利用。因此,加强对建兰种质资源遗传多样性与进化的研究对建兰种质资源保护和利用以及新品种培育具有重要意义。ISSR(inter-simple sequence repeat,内部简单重复序列多态性)是分子标记的一种,不需要提前预知基因组的任何信息就可以进行标记,具有方便快捷等优点,通常具有很好的稳定性和多态性,因而是非常理想的分子标记[1]。ISSR分子标记技术已广泛运用于动植物类种质资源的研究,但在我国兰属植物的遗传学研究中还不够全面深入。传统方法注重兰花的花、叶等表型性状特点并据此来分类,但表型可能受到外部因素等影响,所得结果并不具备极强的准确性。因此,通过分子标记技术分析兰属植物各品种的亲缘关系,可以得到更为科学和稳定的分析结果。
在兰科植物遗传多样性研究上,梁红健等[2-5]通过RAPD(random amplified polymorphic DNA,随机扩增多态性DNA标记)标记方法研究了部分国兰品种植物。孙彩云等[6]采用RAPD分子标记分析50个不同材料的国兰之间的相关相似度(亲缘关系),表明春兰和建兰相似度低,非同组种系。张俊祥等[7]对我国云南地区兰科野生种进行AFLP(amplified fragment length pol?ymorphism,扩增片段长度多态性)分析,推测其种间相似度;严华等[8-9]则使用ISSR标记对38种国兰从遗传角度进行亲疏分类;刘翠华[10]通过ISSR标记对江西野生建兰进行遗传多样性分析研究认为建兰居群内的遗传变异大于居群间的遗传变异;杨光穗等[11]通过对海南区域进行野生兰属的调查取样及SRAP(sequence-related am?plified polymorphism,相关序列扩增多态性)分子标记分析,认为墨兰、建兰、兔耳兰以及寒兰4种种质亲缘关系较近,与传统按外观形状的分类结果一致。高岭等[12]则对兰属进行SCoT(start codon targeted polymorphism,目标起始密码子多态性)分子标记,按照其分子学水平特性将24种亚属大致区分为5类。曹雯静[13]利用SARP分子标记技术,探讨了国兰部分主栽品种的分类和亲缘关系的研究。卢家仕等[14]利用ISSR技术对20份兰科材料进行了遗传多样性研究。陈起馨[15]以玉女兰和黄叶红花墨兰为材料进行杂交,用转录组测序技术开发SSR标记,并得到了兰属植物的分子层面遗传图谱。孙彩云等[16]通过RAPD分子标记国兰28个原生种和部分种的不同品种50个材料间的亲缘关系,证明春兰和建兰是属于不同的组。叶庆生等[17]利用同工酶和SDS-PAGE技术对兰属5个种和2个变种的11个品种进行了分类研究。梁红健等[18]利用聚丙烯酰胺凝胶电泳和淀粉胶凝胶电泳,对国兰的6个种1个变种20个品种叶片的过氧化物酶、酯酶、酸性磷酸酯酶,多酚氧化酶的同工酶进行了分析,共获得98条酶带,为国兰品种鉴定和分类提供了生物化学依据。文李等[19]利用RAPD分子标记检测兰属13个品种的亲缘关系,因为每个兰花品种都有特异的扩增带,可与其他品种区分开来,这种特异的条带可作为一个品种的分子标记应用于品种鉴定和分类,可以为中国兰花品种的鉴定和分类提供新的途径。本研究运用ISSR分子标记技术,对39份建兰种质资源的遗传多样性和亲缘关系进行分析,以期为建兰种质关系鉴定、优良基因型挖掘和保护、新品种选育提供理论依据。
1 材料与方法
1.1 材料
实验所用39份建兰材料是2017年6月陆续从台湾、四川、福建、广东等地收集而来,种植在广西农业科学院花卉研究所资源圃。详见表1。
1.2 建蘭材料DNA的提取
利用北京全式金生物有限公司生产的新型植物基因组DNA提取试剂盒提取建兰叶片的DNA,并用1.0%的琼脂糖凝胶电泳检测。提取的样品基因组DNA用核酸微量测定仪稀释至一致浓度(50 ng/?L),置于–20 ℃下保存备用。
1.3 建兰材料ISSR-PCR扩增体系构建
参照陈云飞[20]针对金佛山方竹研究所提出的PCR体系,在此基础之上进行细微调整,利用20 ?L反应体系当中的DNA模板1.0 ?L(20 ng DNA),其中引物1.0 ?L,1xT3 Super PCR Mix 15.0 ?L,ddH2O 3 ?L。PCR反应程序为:98 ℃预变性3 min,98 ℃变性30 s,根据引物对应退火温度标准退火30 s,采用72 ℃高温持续120 s,并35次重复,随后在72 ℃温度条件下持续退火300 s。
1.4 ISSR引物筛选与ISSR-PCR扩增
使用1号‘梨山狮王和2号‘君荷2份供试材料的DNA通过对生工生物工程(上海)股份有限公司合成的36条通用引物进行筛选。遵照既定反应体系和反应程序,经过初步筛选及温度梯度复筛,筛选出6条条带多态性高、重复性好、条带清的引物,可以用于建兰的ISSR-PCR扩增,分别为P809、P815、P827、P835、P836、P842,其引物序列号、碱基数以及退火温度已经列出(表2)。基于优化的扩增体系,将筛选出的6条引物对39份建兰材料基于优化反应体系和扩增过程进行PCR扩增,采用1.0%琼脂糖凝胶电泳检测扩增产物(电泳缓冲液浓度为1xTAE,电压值为120 V),用Bio-Rad凝胶系统进行成像处理。
1.5 数据处理
在数据处理阶段,采用人工阅读的方式来分析和解读电泳扩增图谱,将电泳清晰和可重复的记录为“1”,其他情况则记录为“0”,随后依据记录结果生成原始数据矩阵。根据NeiandLi提出的方法,由分子生物学分析软件NTSYS计算供试材料的遗传相似系数,使用UPGMA法对数据进行遗传相似性聚类分析[21]。
2 结果与分析
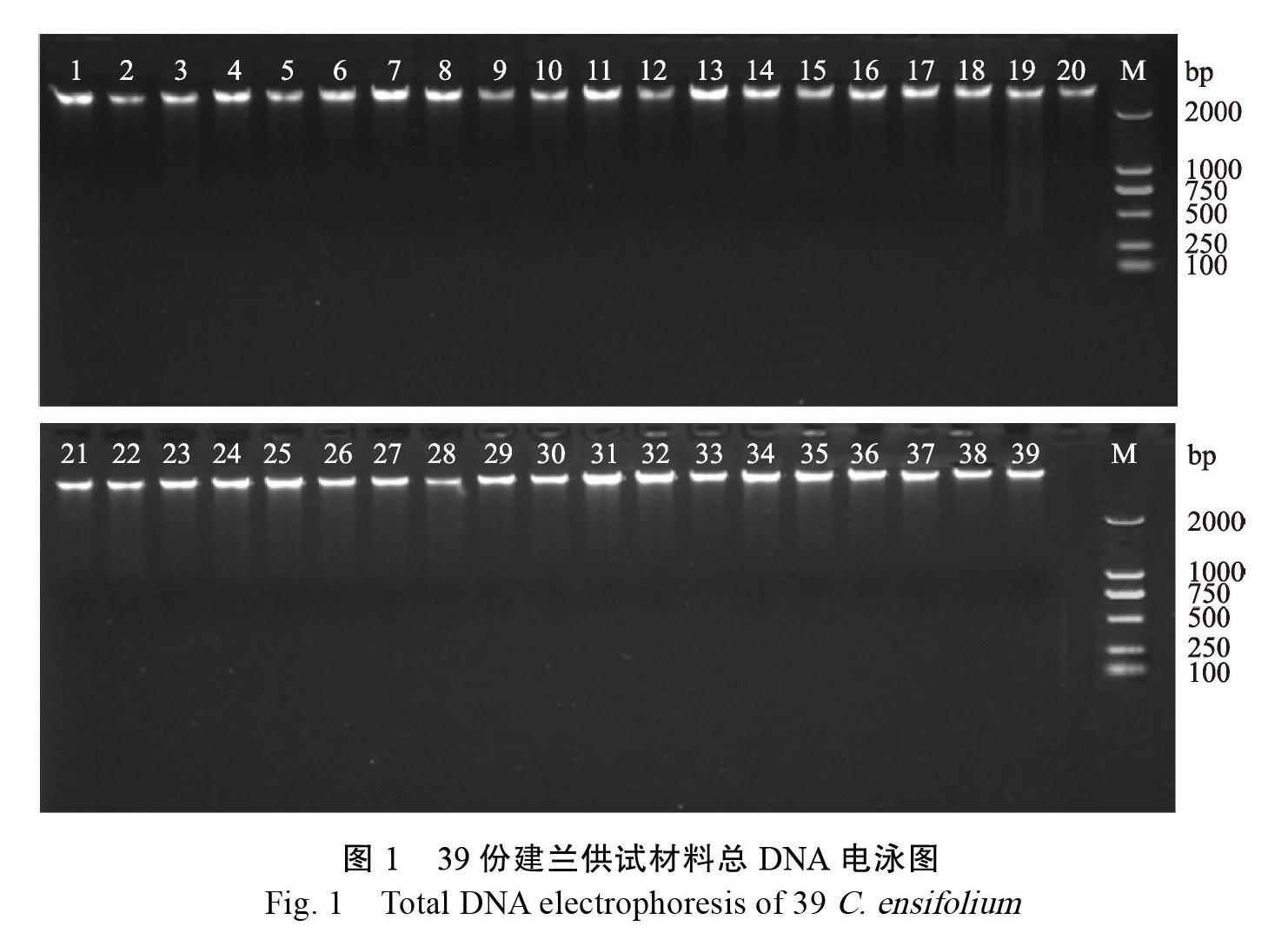
2.1 供试材料基因组总DNA提取
经紫外分光仪检测结果表明,本次实验提取的建兰供试材料的基因组总DNA OD260/OD280在1.8~2.0之间,DNA浓度均大于280 ng/?L。通过1%琼脂糖凝胶电泳测试,结果如图 所得条带清晰,无蛋白质RNA污染,无残留无拖尾现象且纯度高,能够满足ISSR标记分析的要求。
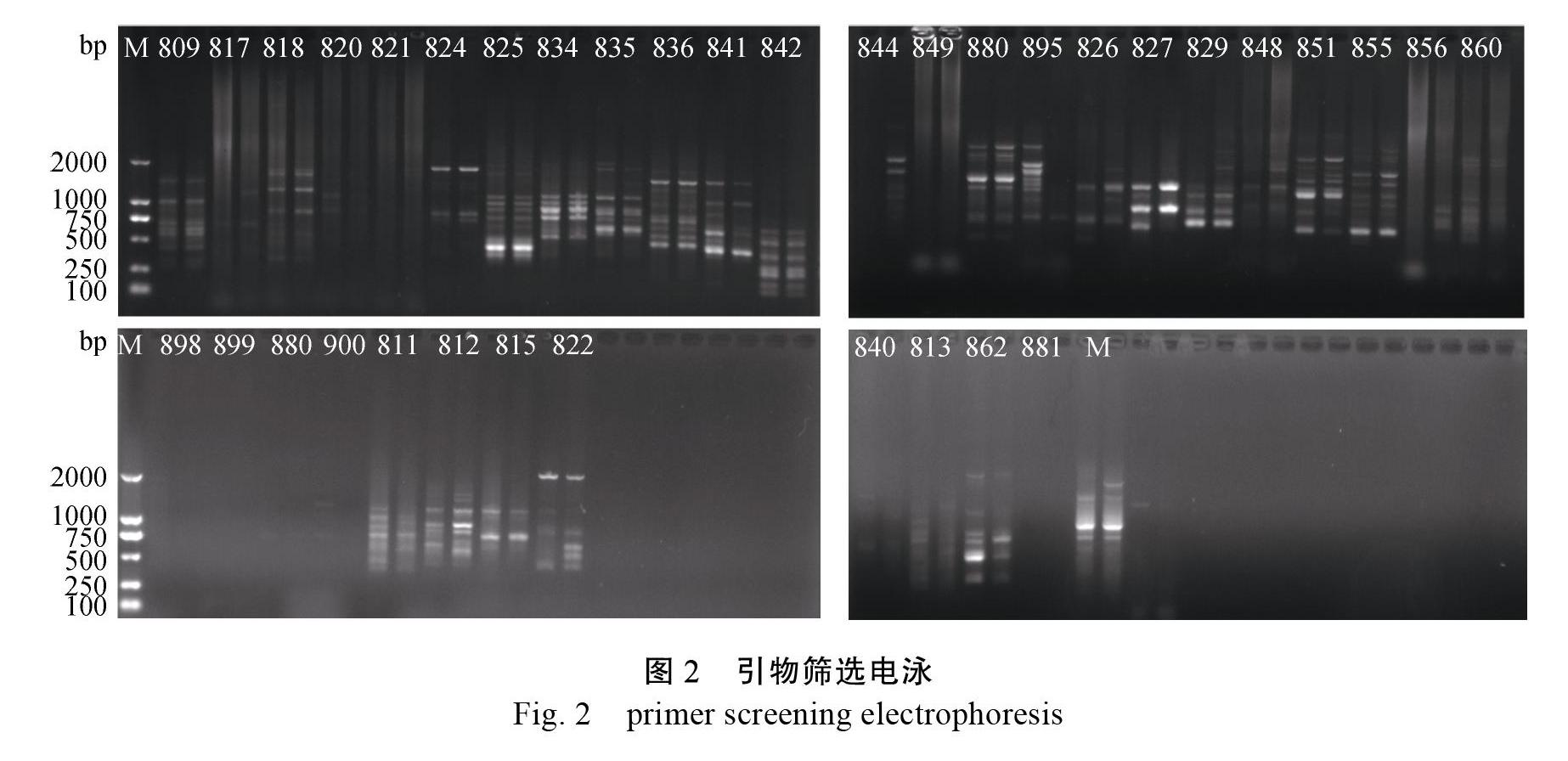
2.2 引物筛选
使用1号‘梨山狮王兰花和2号‘君荷2份供试材料的DNA对36条ISSR引物进行引物筛选,结果显示(图2),其中P809、P815、P827、P835、P836、P842引物较理想,因此选择他们进行ISSR-PCR扩增。
2.3 引物筛选后扩增
利用筛选获得的6个ISSR引物P809、P815、P827、P835、P836、P842对39个供试样品按照优化的PCR进行PCR扩增,结果如图3。
2.4 引物扩增条带多态性统计分析
如表2所示,6条引物对39个建兰种质材料共扩增出64条清晰带纹,其中57条多态性条带。平均每条引物检测到的条带数为11条,多态性条带10条,多态性比率为89.88%。说明ISSR标记在建兰的品种中具有丰富的多态性,能够很好地揭示建兰品种间的亲缘关系。
2.5 建兰实验材料间的遗传相似性分析
39份供试材料的遗传相似性系数范围为0.500~0.95 说明供试的39份建兰材料的遗传变异较为丰富。其中1号台湾花莲的‘梨山狮王与17号广东揭阳的‘秋雪,二者之间的遗传相似系数均达到0.95 表明它们之间的遗传物质存在很大程度上的相似性;其次是25号广东佛山的‘水晶皇梅与32号广东潮汕的‘铁骨素梅,二者的遗传相似性系数为0.875,表明这2份材料间的亲缘关系较近;遗传相似性系数最小的是3号四川自贡的‘黄光登和39号台湾台中的‘锦旗,8号福建漳州的‘红月、18号四川峨眉山的‘中华彩霞和38号台湾基隆的‘市长红,其值均为0.500,表明这二者间的遗传差距最大,亲缘关系最远。
2.6 39个建兰材料的聚类分析
如图4所示,在遗传相似系数约为0.71时可将39份建兰材料分成6个群集:
类群I:‘梨山狮王‘秋雪‘君荷蝶‘朝阳三星蝶‘黄一品‘富山奇蝶‘小桃红红月‘峨眉牡丹‘一品梅‘红君荷遗传相似系数约在0.77时,类群I可以划分为7个亚类,I-i亚类包括‘梨山狮王‘秋雪等18份材料,这18份材料依照叶长和萼片长等性状分别聚在2个大类中。‘梨山狮王与‘秋雪种内材料之间聚在同一个类群中,表明‘梨山狮王与‘秋雪种内材料的遗传背景相似,亲缘关系比较接近,基于表型性状的聚类中‘梨山狮王与‘秋雪种内材料聚为一类,表明这2种聚类结果较一致;I-ii亚类包括‘龙岩素、‘水晶皇梅、‘铁骨素梅和‘青山玉泉4份材料,因为生长周期或者环境影响,‘龙岩素没有采集到花葶着花数、萼片长等表型性状,但是ISSR分子标记说明的是这些材料间的遗传背景相似。类群Ⅱ、Ⅲ、Ⅴ、Ⅵ有且仅有一个建兰品种被单独聚在一起,说明其跟其他群集的建兰在遗传水平上有较大差异,与其他类群遗传背景较远。类群Ⅳ包括‘绿梅、‘荷三星和‘峨眉荷仙3份材料,虽然没有采集到‘绿梅和‘峨眉荷仙的表型参数,但它们基于ISSR标记聚类被聚到一起,表明这3种建兰材料虽然来源地不同,但亲缘关系较近。
3 讨论
与其他分子标记相比,ISSR标记为研究者提供了一种能够有效跟踪性状相关目的基因的新型分子标记[22]。本研究对ISSR-PCR反应体系和扩增程序进行优化,并从36条ISSR引物中最终筛选出6条扩增条带清晰、重复性高、多态性好的引物,利用这6个引物对39份建兰材料进行PCR扩增,共扩増出64条带,其中多态性条带57条。平均每条引物检测到的条带数为11条,多态性条带为10条,多态性比率为89.88%。ISSR标记的遗传相似性系数范围为0.500~0.95 表明39份实验材料具有较丰富的遗传多样性。对39个供试材料的ISSR分子标记使用层次聚类法进行树状图绘制,在遗传相似系数约为0.71时39个建兰供试品种分成6个群集。
本研究中的39份建兰实验材料具有较丰富的遗传多样性,其ISSR标记的遗传相似性系数范围为0.500~0.953。唐源江等[23]曾利用SRAP分子标记技术对国兰的遗传相似性进行了研究,实验材料是139份的国兰种质材料,实验结果得出来的相似系数在0.51~0.91之间变化;袁媛等[24]也利用SRAP分子标记技术进行了国兰的研究,其实验材料是154份国兰材料,所得出来的相似系数在0.772~1.000之间。本次研究与唐源江等[23]的研究结果相似,与袁媛等[24]的研究结果存在一定的差异。导致这种情况的原因应该是研究材料的不同,或者使用的分子标记技术不同所导致的。39份建兰种质在遗传相似系为0.71左右处被聚为6个类群,白坚等[25]对47个建兰品种进行SRAP聚类,在相似系数为0.68时47份建兰品种被聚为四大类,结果表明相同地区的建兰品种亲缘关系较近,这与本实验结论大体一致。胡薇等[26]对建兰的38个品种使用RAPD标记聚类,在遗传距离为0.307 2处,38个建兰品种被聚为9组,结果表明彩心品种较素心品种遗传分化高,本实验39份建兰材料中唇瓣为素心的分别为‘素君荷‘秋雪‘铁骨素梅,唇瓣为彩心仅有一份材料‘青山玉泉,它们之间的遗传相似系数依次为0.766、0.734、0.766,二者之间相似性较高。关于建兰花色变化的研究是否有多基因參与调控,有哪些基因参与调控,调控机制如何等方面的研究有待进一步深入。
本研究明确了广西农科院花卉研究所收集保存的39份建兰种质资源的遗传变异相近程度,可为今后建兰杂交选育及杂交种间遗传多样性研究提供基础数据。同一物种国兰间的性状差异较大,这可能是由于遗传变异,种群动态以及环境等因素的影响,更为后续开展优良品种提供有效依据,对兰属种质资源的开发与利用有重要意义。
参考文献
[1] 孙 玥, 苏京平, 王胜军, 等. ISSR分子标记技术在作物遗传育种中的应用[J]. 种子科技, 2019, 37(12): 26-27, 30.
[2] 梁红建, 刘 敏, 钟志宇, 等. 中国部分兰花品种RAPD分析[J]. 园艺学报, 1996(4): 365-370.
[3] 王慧中, 王玉东, 周晓云, 等. 兰属14种植物遗传多样性RAPD及AFLP分析[J]. 实验生物学报, 2004(6): 482-486.
[4] Choi S H, Kim M J, Lee J S, et al. Genetic diversity and phylogenetic relationships among and within species of oriental cymbidiums based on RAPD analysis[J]. Scientia Horticulturae, 2006, 108(1): 79-85.
[5] 蹇 黎, 武 海, 張以忠, 等. 21个兰花品种的延长RAPD(ERAPD)和PCR-RFLP分析[J]. 中国农学通报, 2010, 26(14): 232-237.
[6] 孙彩云, 张明永, 叶秀粦, 等. 中国兰属植物种间及品种间亲缘关系的RAPD分析[J]. 园艺学报, 2005(6): 1121-1124.
[7] 张俊祥, 李枝林, 范成明, 等. 云南野生兰属主要种间亲缘关系的AFLP分析[J]. 园艺学报, 2006(5): 1141-1144.
[8] 严 华, 张冬梅, 罗玉兰, 等. 38种国兰亲缘关系的ISSR分析[J]. 分子植物育种, 2010, 8(4): 736-741.
[9] 吴振兴, 王慧中, 施农农, 等. 兰属Cymbidium植物ISSR遗传多样性分析[J]. 遗传, 2008(5): 627-632.
[10] 刘翠华. 江西省野生建兰的ISSR遗传多样性研究[D]. 南昌: 南昌大学, 2012.
[11] 杨光穗, 任 羽, 王荣香. 利用SRAP技术分析海南野生兰属植物的亲缘关系[J]. 热带农业科学, 2011, 31(9): 1-3.
[12] 高 岭, 冯尚国, 何仁锋, 等. 兰属植物目标起始密码子(SCoT)遗传多样性分析[J]. 园艺学报, 2013(10): 2026- 2032.
[13] 曹雯静. 国兰部分主栽品种遗传多样性及分子鉴定研究[D]. 厦门: 华侨大学, 2014.
[14] 卢家仕, 卜朝阳, 吕维莉, 等. 20份兰科植物的ISSR遗传多样性分析[J]. 西南农业学报, 2012(6): 2252-2257.
[15] 陈起馨. 兰花分子图谱的构建[D]. 广州: 华南农业大学, 2016.
[16] 孙彩云, 张明永, 梁承邺, 等. 墨兰、春兰变种和品种间同工酶分析[J]. 园艺学报 , 2002(1): 75-77.
[17] 叶庆生, 文 李, 潘瑞炽. 利用同工酶和SDS-PAGE技术对一些兰属(Cymbidium)品种的分析(简报)[J]. 热带亚热带植物学报, 1999(4): 337-341.
[18] 梁红健, 刘 敏, 张纯花, 等. 中国兰花(Chinese Cymbidium)部分品种的叶片同工酶分析[J]. 实验生物学报, 1997(3): 343-348.
[19] 文 李, 叶庆生, 王小菁, 等. 利用RAPD技术分析兰属(Cymbidium)品种间的亲缘关系[J]. 应用与环境生物学报, 2001(1): 29-32.
[20] 陈云飞. 基于遗传多样性的金佛山方竹采样策略初探[D]. 贵阳: 贵州大学, 2018.
[21] Hua G J, Hung C L, Lin C Y, et al. MGUPGMA: A fast UPGMA algorithm with multiple graphics processing units using NCCL[J]. Evolutionary Bioinformatics, 2017, 13(1): 11-27.
[22] 邱国俊, 程 敏, 郭计华. ISSR分子标记技术在植物中的应用及其研究进展[J]. 兴义民族师范学院学报, 2020(1): 117-120.
[23] 唐源江, 曹雯静, 吴坤林. 基于SRAP标记的国兰种质资源遗传多样性分析及分子身份证构建[J]. 中国农业科学, 2015(9): 1795-1806.
[24] 袁 媛, 曹 彬, 张咏琪, 等. 基于SRAP标记的国兰种质资源遗传多样性分析[J]. 热带作物学报, 2019(4): 1-17.
[25] 白 坚, 胡 旭, 周淑婷, 等. 47个建兰品种的SRAP遗传多样性分析[J]. 植物遗传资源学报, 2012(3): 376-380.
[26] 胡 薇, 黄儒珠, 潘晓华, 等. 建兰38个品种的RAPD分析[J]. 园艺学报, 2008(2): 289-294.
责任编辑:白 净
