葡萄糖氧化酶对产蛋后期蛋种鸡产蛋性能、孵化性能及抗氧化能力的影响
2021-11-05李嘉辉李生杰龚建刚荆佳林郝艳霜冯志华赵国先
李嘉辉 李生杰 龚建刚 荆佳林 郝艳霜 冯志华* 赵国先*
(1.河北农业大学动物科技学院,保定 071001;2.合肥正大有限公司,合肥 230601;3.河北农业大学食品科技学院,保定 071001)
蛋种鸡的饲养周期相对较长,产蛋后期种鸡生理机能逐渐退化,同时由于集约化养殖环境中各种应激因素的影响,鸡群常处于氧化应激状态,对疾病的抵抗力减弱,产蛋性能和孵化性能大幅降低,利用年限缩短,提高了养殖难度和养殖成本。因此,研究高效绿色饲料添加剂对产蛋后期蛋种鸡繁殖性能和抗氧化能力的影响对于提高蛋种鸡的利用率、延迟蛋种鸡下架时间和提高种禽饲养的经济效益有重要意义。葡萄糖氧化酶(glucose oxidase,GOD)作为一种功能性饲料添加剂,具有无残留、无抗药性、绿色安全等优点,在动物生产中的应用越来越广泛。已有研究表明,GOD可降低动物肠道pH,改善肠道健康,促进营养物质吸收利用,提高畜禽生产性能,增强机体抗氧化能力和免疫功能[1-7]。然而目前有关GOD在种禽上的研究甚少,其对产蛋后期海兰褐蛋种鸡产蛋性能、孵化性能和抗氧化能力方面的研究还未见报道。因此,本试验通过在产蛋后期蛋种鸡饲粮中添加不同水平的GOD,探究其对蛋种鸡产蛋性能、孵化性能、抗氧化能力及相关基因表达的影响,以期为GOD的抗氧化作用机制研究及其在种鸡生产中的合理应用提供科学依据。
1 材料与方法
1.1 试验材料
GOD购自宁夏某有限公司,活性为1 200 U/g。
1.2 试验设计
采用单因素随机试验设计,选用480只55周龄、体重和产蛋率基本一致的海兰褐蛋种母鸡,随机分为4组,每组6个重复,每个重复20只鸡。对照组饲喂基础饲粮,试验组分别饲喂在基础饲粮中添加200、350和500 mg/kg GOD的试验饲粮。预试期1周,正试期12周。
1.3 基础饲粮及饲养管理
参照NRC(1994)蛋种鸡营养标准和企业生产情况配制玉米-豆粕型基础饲粮,其组成及营养水平见表1。各组试验鸡于半机械通风舍内使用3层阶梯笼养,自由采食和饮水,每隔5 d人工授精1次,每天人工光照16 h,光照强度为20 lx。试验期间各组试验鸡均保持相同的环境条件,并按常规饲养管理规程及免疫程序进行饲养和免疫。

表1 基础饲粮组成及营养水平(风干基础)
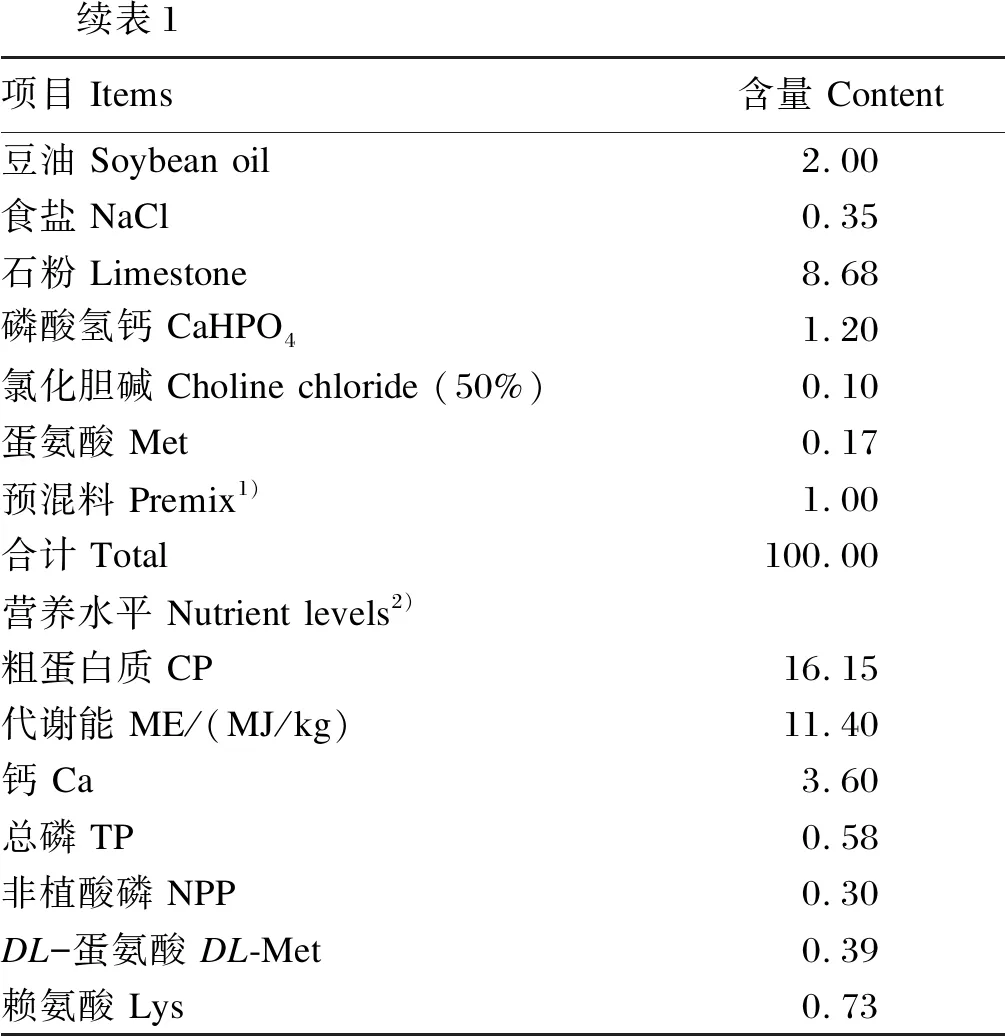
续表1项目 Items含量 Content豆油 Soybean oil2.00食盐 NaCl0.35石粉 Limestone8.68磷酸氢钙 CaHPO41.20氯化胆碱 Choline chloride (50%)0.10蛋氨酸 Met0.17预混料 Premix1)1.00合计 Total100.00营养水平 Nutrient levels2)粗蛋白质 CP16.15代谢能 ME/(MJ/kg)11.40钙 Ca3.60总磷 TP0.58非植酸磷 NPP0.30DL-蛋氨酸 DL-Met0.39赖氨酸 Lys0.73
1.4 检测指标及方法
1.4.1 产蛋性能及孵化性能指标
产蛋性能:试验期间每天15:00收集种蛋,记录各重复产蛋数、总蛋重和采食量,并记录破损蛋、软壳蛋、沙壳蛋、双黄蛋和畸形蛋等不合格蛋数。试验结束后对各组产蛋率、平均蛋重、平均日采食量、料蛋比和合格蛋率进行统计。
产蛋率(%)=100×总产蛋数/(鸡数×天数);平均蛋重(g)=总蛋重/总产蛋数;平均日采食量(g/d)=总采食量/(鸡数×天数);料蛋比=总采食量/总蛋重;合格蛋率(%)=100×合格蛋数/产蛋数。
孵化性能:试验最后1周,每个重复收集50枚合格种蛋进行孵化指标的测定,并于孵化第18天进行照蛋,识别并捡出未受精种蛋,孵化过程中记录各重复入孵蛋数、受精蛋数和出雏数,计算受精率、孵化率和出雏率。
受精率(%)=100×受精蛋数/入孵蛋数;
孵化率(%)=100×出雏数/受精蛋数;
出雏率(%)=100×出雏数/入孵蛋数。
1.4.2 蛋品质
试验第12周末,每重复随机挑选5枚合格种蛋进行蛋品质测定。蛋形指数使用蛋形指数测量仪(NFN-385,以色列)进行测定,蛋壳强度、蛋白高度和哈氏单位使用多功能蛋品质测定仪(EMT-5200)进行测定,蛋壳厚度使用蛋壳厚度测量仪(ETG-1061型)测定。
1.4.3 血清及肝脏抗氧化指标
试验结束后,各组以重复为单位随机选取2只接近平均体重的健康种鸡,翅静脉采血,3 000 r/min离心15 min后将血清分装于离心管,置于-20 ℃保存。采血完成后将试验鸡屠宰并解剖,取适量相同部位的肝脏组织,生理盐水冲洗后将分装于5 mL离心管中,置于液氮中,随后转移到-80 ℃冰箱保存。使用试剂盒测定血清及肝脏丙二醛(MDA)含量、超氧化物歧化酶(SOD)、谷胱甘肽过氧化物酶(GSH-Px)活性和总抗氧化能力(T-AOC)。所用试剂盒均购于南京建成生物工程研究所,并严格按照试剂盒说明书进行操作。
1.4.4 肝脏抗氧化相关基因的mRNA表达
样品采集:在进行1.4.3步骤的同时,额外取试验鸡2块筋膜较少的相同部位的肝脏组织,分装于1.5 mL冻存管后,迅速投入液氮,最后置于-80 ℃保存。
肝脏抗氧化相关基因mRNA相对表达量的测定步骤:1)使用超纯RNA提取试剂盒(Cat#CW0581,CWbio. Co., Ltd.,)进行总RNA提取;取5 μL RNA,1%琼脂糖凝胶进行电泳,检测RNA的完整性。2)使用HiFi-MMLV cDNA第一链合成试剂盒(Cat#CW0744,CWbio. Co., Ltd.,)进行反转录。3)应用Line Gene 9 600 Plus型PCR仪进行荧光定量,以β-肌动蛋白(β-actin)作为内参基因,采用2-ΔΔCt法计算目的基因的相对表达量。引物序列(表2)由北京基谱生物科技有限公司设计并合成。

表2 实时定量PCR引物序列
1.4.5 经济效益计算
经济效益[元/(只·d)]=合格种蛋收入-
基础饲粮成本-GOD成本。
1.5 数据统计分析
采用SPSS 22.0统计软件中的单因素方差分析(one-way ANOVA)程序对数据进行统计分析,并利用Duncan氏法进行多重比较,试验结果均以“平均值±标准差”表示,P<0.05表示差异显著。
2 结果与分析
2.1 GOD对产蛋后期蛋种鸡产蛋性能和孵化性能的影响
由表3可知,与对照组相比,350和500 mg/kg GOD组产蛋后期蛋种鸡的产蛋率显著增加(P<0.05),料蛋比显著降低(P<0.05);200、350和500 mg/kg GOD组总产蛋数分别较对照组增加1.92、3.12和3.85枚/只,总合格蛋数较对照组分别增加1.97、3.74和3.86枚/只;与对照组相比,200、350和500 mg/kg GOD组的种蛋受精率显著增加(P<0.05),但不同水平GOD添加组之间差异不显著(P>0.05);饲粮中添加不同水平GOD对平均日采食量、平均蛋重、合格蛋率、种蛋孵化率和出雏率均无显著影响(P>0.05)。
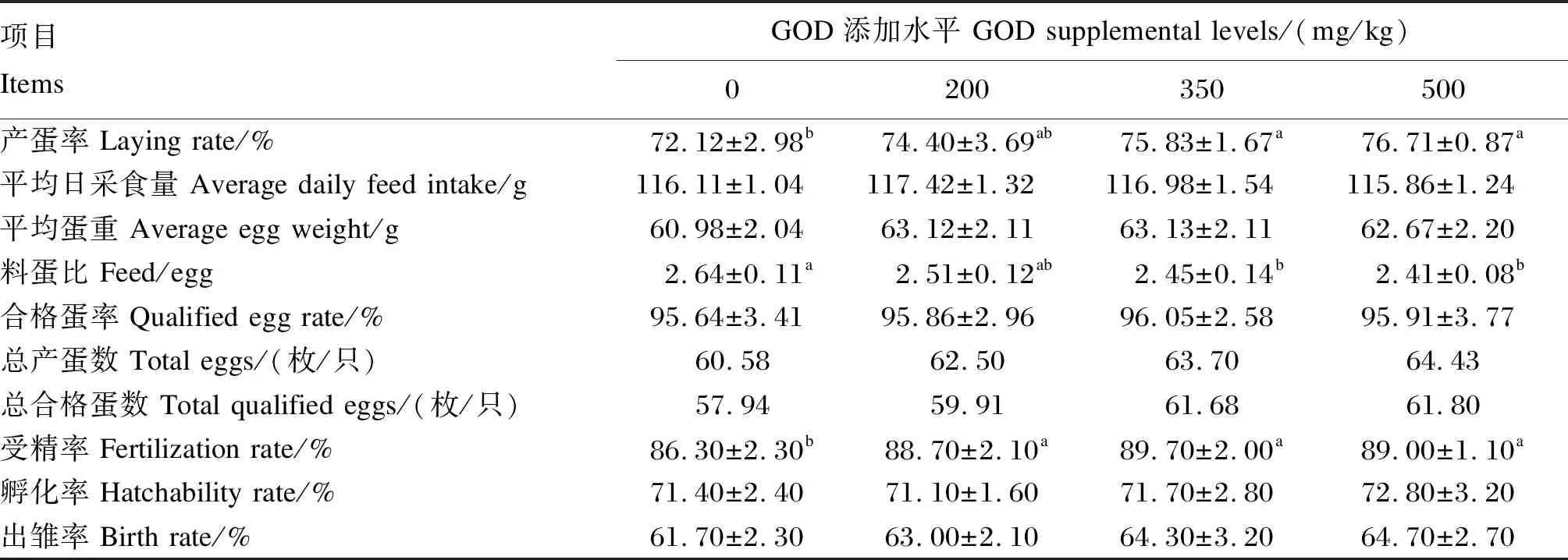
表3 GOD对产蛋后期蛋种鸡产蛋性能和孵化性能的影响
2.2 GOD对产蛋后期蛋种鸡蛋品质的影响
由表4可知,与对照组相比,350和500 mg/kg GOD组产蛋后期蛋种鸡的种蛋蛋壳厚度及哈氏单位显著增加(P<0.05);饲粮中添加不同水平GOD对种蛋的蛋壳强度、蛋白高度和蛋形指数均无显著影响(P>0.05)。
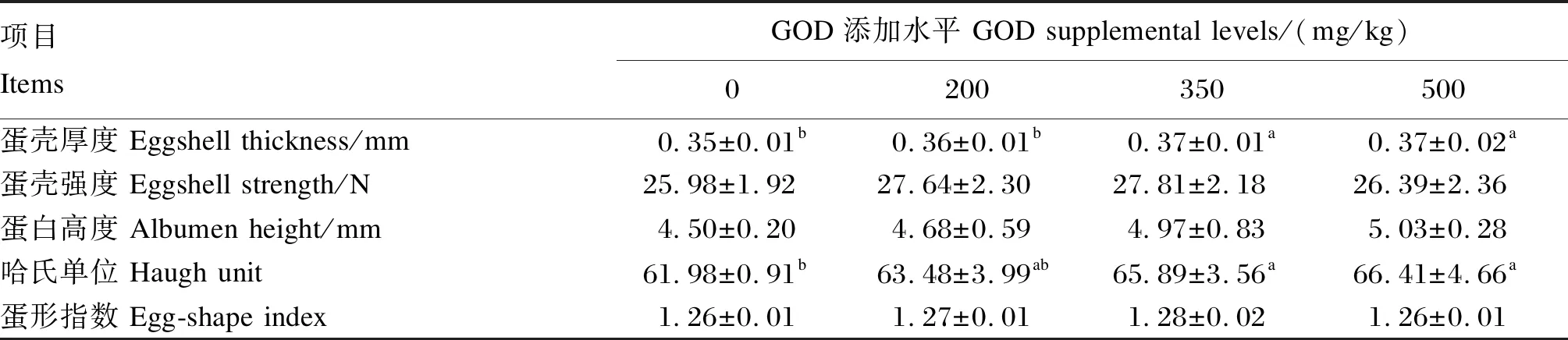
表4 GOD对产蛋后期蛋种鸡蛋品质的影响
2.3 GOD对产蛋后期蛋种鸡血清抗氧化指标的影响
由表5可知,500 mg/kg GOD组产蛋后期蛋种鸡的血清MDA含量显著低于对照组、200和350 mg/kg GOD组(P<0.05);与对照组相比,200、350和500 mg/kg GOD组的血清SOD活性和T-AOC显著增加(P<0.05),但不同水平GOD添加组之间无显著差异(P>0.05);与对照组相比,350和500 mg/kg GOD组的血清GSH-Px活性显著增加(P<0.05)。
2.4 GOD对产蛋后期蛋种鸡肝脏抗氧化指标的影响
由表6可知,与对照组相比,200、350和500 mg/kg GOD组产蛋后期蛋种鸡的肝脏MDA含量显著降低(P<0.05),肝脏SOD活性显著增加(P<0.05),但不同水平GOD添加组之间肝脏MDA含量和SOD活性无显著差异(P>0.05);350和500 mg/kg GOD组的肝脏GSH-Px活性显著高于对照组和200 mg/kg GOD组(P<0.05);饲粮中添加GOD对肝脏T-AOC无显著影响(P>0.05)。
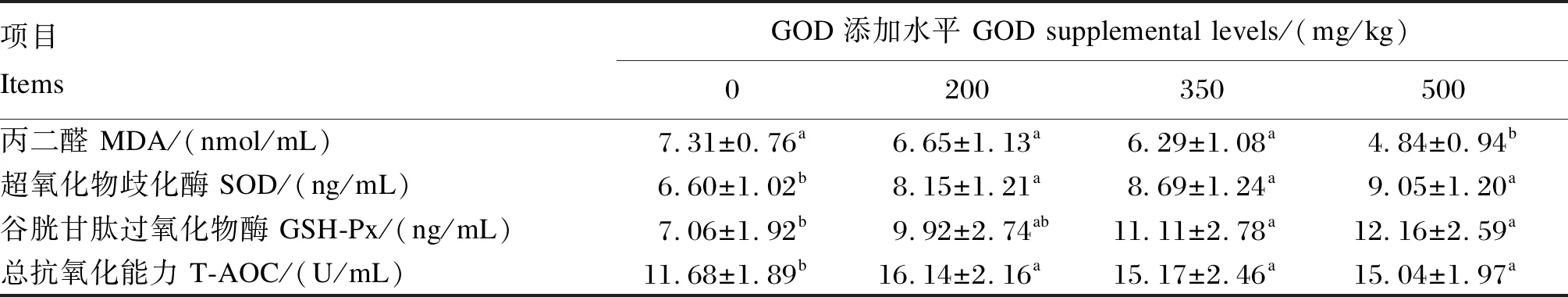
表5 GOD对产蛋后期蛋种鸡血清抗氧化指标的影响
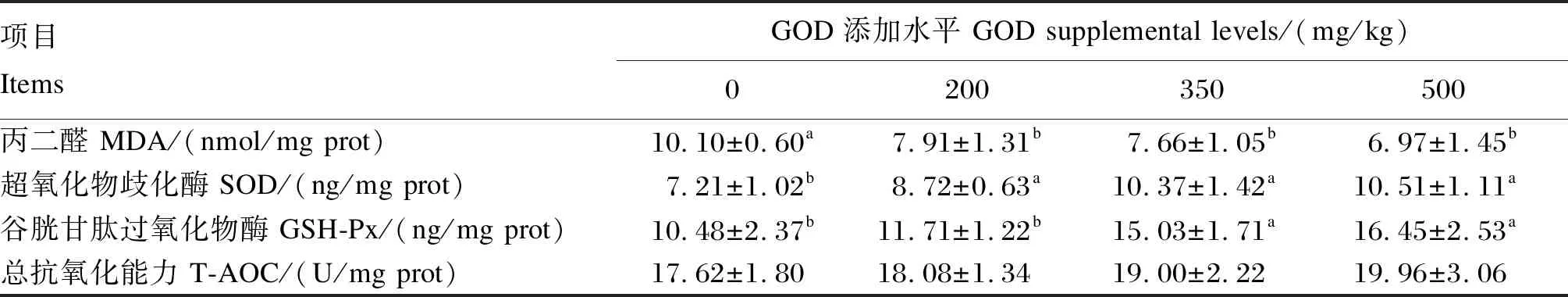
表6 GOD对产蛋后期蛋种鸡肝脏抗氧化指标的影响
2.5 GOD对产蛋后期蛋种鸡肝脏抗氧化相关基因mRNA相对表达量的影响
由表7可知,饲粮中添加不同水平的GOD显著上调了产蛋后期蛋种鸡肝脏核因子E2相关因子(Nrf2)、铜锌超氧化物歧化酶(Cu/Zn-SOD)和锰超氧化物歧化酶(Mn-SOD)的mRNA相对表达量(P<0.05);GOD添加组的肝脏Nrf2的mRNA相对表达量均显著高于对照组(P<0.05),但不同水平GOD添加组之间无显著差异(P>0.05);500 mg/kg GOD组的肝脏Cu/Zn-SOD的mRNA相对表达量显著高于对照组和200 mg/kg GOD组(P<0.05),但500与350 mg/kg GOD组之间无显著差异(P>0.05);GOD添加组的肝脏的Mn-SOD的mRNA相对表达量显著高于对照组(P<0.05),350与500 mg/kg GOD组之间无显著差异(P>0.05)。
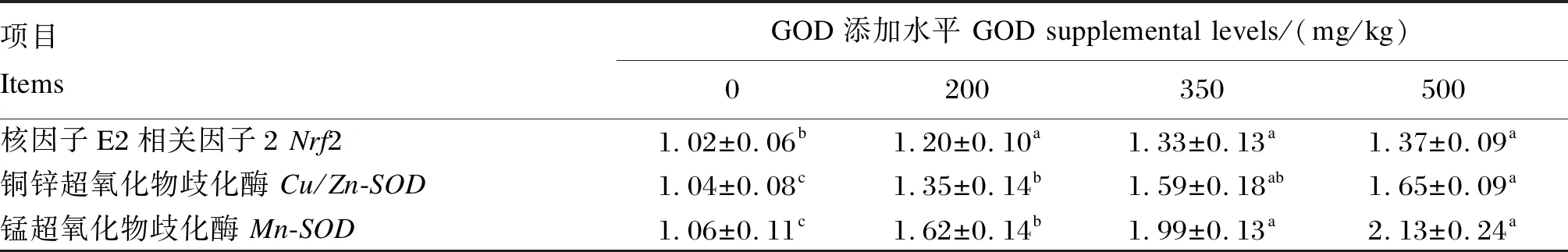
表7 GOD对产蛋后期蛋种鸡肝脏抗氧化相关基因mRNA相对表达量的影响
2.6 GOD对产蛋后期蛋种鸡经济效益的影响
由表8可知,饲粮中添加不同水平的GOD均能不同程度提高产蛋后期蛋种鸡的经济效益。在仅考虑饲粮成本的前提下,当GOD的添加水平为500 mg/kg时利润最高,每只鸡每日盈利1.030元,与对照组相比增加8.42%。
3 讨 论
3.1 GOD对产蛋后期蛋种鸡产蛋性能和孵化性能的影响
处于产蛋后期的蛋鸡,其本身生殖机能和抗氧化能力均有不同程度地减弱,而现代种鸡养殖多采用高密度笼养,鸡群常处于应激状态,进而导致产蛋性能和种蛋质量下降,孵化性能也显著降低[8-9]。研究表明,活性氧引起的氧化应激是机体衰老和卵巢功能衰退的主要机制之一[10-11]。因此,在饲粮中添加功能性添加剂可以缓解产蛋后期蛋种鸡的氧化应激状态,从而改善其生产性能。赵必迁等[12]在产蛋后期罗曼粉壳蛋鸡饲粮中添加300 mg/kg的GOD,发现显著提高了产蛋率和饲料转化率。曲浩杰等[13]在产蛋后期海兰褐蛋鸡饲粮中添加博落回提取物和GOD,发现产蛋率和料蛋比均有显著改善。本研究结果与上述报道一致,饲粮中添加350和500 mg/kg的GOD使产蛋后期海兰褐蛋种鸡的产蛋率和料蛋比得到显著改善。目前有关GOD对种用畜禽繁殖性能影响的报道有限。在猪上的研究表明,妊娠期和哺乳期母猪饲粮中添加60 U/kg的GOD能够改善母猪和仔猪的氧化还原状态,提高母猪的繁殖性能[7]。另据报道,在34周龄蛋种鸡饲粮中添加100 mg/kg的GOD对产蛋率、种蛋孵化率、受精率和健雏率等指标均无显著影响[14]。本研究中,饲粮中添加不同水平(200、350和500 mg/kg)GOD均显著改善了种蛋受精率。试验结果的差异可能归因于试验动物的种类、周龄、GOD的活性或添加水平不同。
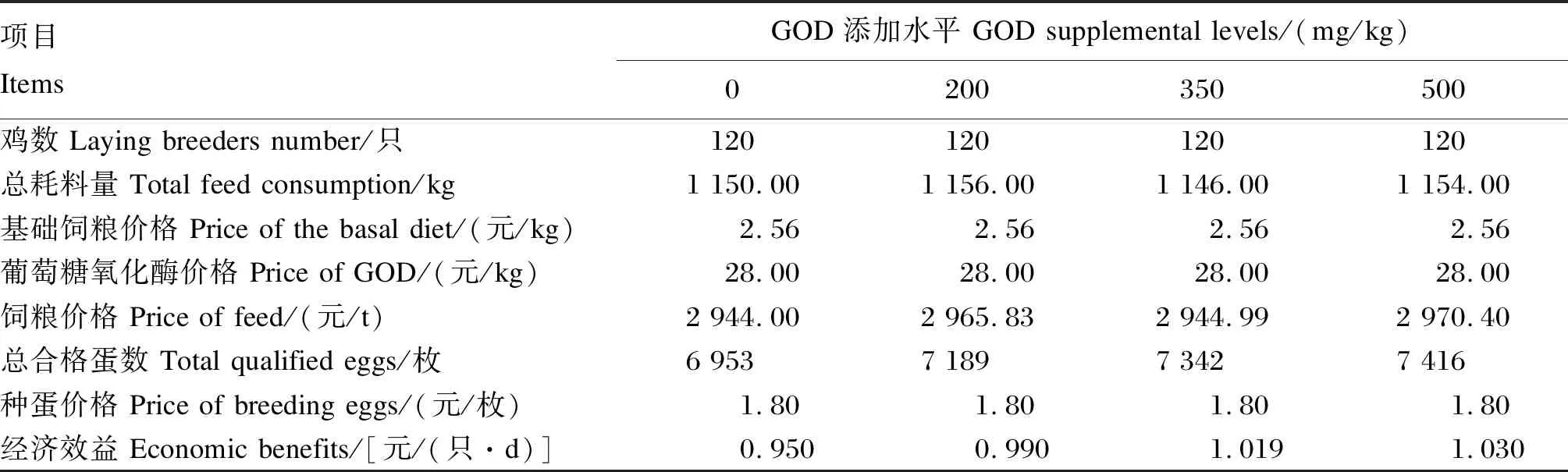
表8 GOD对产蛋后期蛋种鸡经济效益的影响
3.2 GOD对产蛋后期蛋种鸡蛋品质的影响
蛋壳厚度、蛋壳强度、蛋白高度、哈氏单位和蛋形指数是衡量蛋品质的重要指标。赵国先等[15]研究表明,在26周龄海兰褐蛋鸡饲粮中添加0.3%的GOD可显著提高鸡蛋的蛋壳厚度和哈氏单位。曲浩杰等[13]将GOD和博落回提取物联合使用,发现对55周龄产蛋后期海兰褐蛋鸡的蛋壳厚度无显著影响,但显著提高了蛋白高度。本试验中,饲粮中添加350和500 mg/kg的GOD显著提高了种蛋的蛋壳厚度和哈氏单位,蛋壳强度也有所增加,但未达到显著水平。研究结果的不同,可能与试验动物的种类、产蛋阶段、饲粮组成及养殖环境等因素有关。蛋壳质量不仅影响种蛋的运输安全,还会影响种蛋的孵化率[16]。哈氏单位是表示鸡蛋新鲜程度和蛋白质量的重要指标,其与种蛋孵化率也密切相关[17]。研究表明,氧化应激可抑制钙离子在肠道内的吸收,也会阻碍其在输卵管壳腺部的转运,影响蛋壳品质[18-19]。本试验结果显示,饲粮中添加350和500 mg/kg的GOD显著改善了产蛋后期蛋种鸡的氧化应激状态,这与种蛋的蛋壳质量和哈氏单位得到显著改善的结果相一致。此外,本课题组前期研究结果表明,饲粮中添加300和400 mg/kg的GOD可显著提高蛋鸡对钙、总磷和粗蛋白质等的消化率[5]。因此推测,GOD可能通过减轻产蛋后期蛋种鸡的氧化应激状态,改善机体钙、磷和蛋白质代谢来提高种蛋的蛋壳厚度和哈氏单位,其具体机制还有待进一步研究。
3.3 GOD对产蛋后期蛋种鸡抗氧化能力的影响
血清及肝脏中GSH-Px、SOD活性和T-AOC、MDA含量都是反映机体抗氧化能力的重要指标。GSH-Px和SOD作为机体酶促抗氧化系统的第1道防线,可消除自由基,维持细胞内氧化还原平衡[20]。MDA为细胞膜脂质过氧化的终产物,可参照其含量来间接估计机体所受氧化损伤的程度。T-AOC是体现机体抗氧化能力的综合指标[21]。宋海彬[22]研究发现,饲粮中添加GOD可使肉鸡血清SOD活性不同程度增加,MDA含量下降。Wang等[2]研究证实,GOD可显著提高岭南黄羽肉鸡肝脏GSH-Px活性和T-AOC,显著降低MDA含量,增强机体抗氧化能力。另有研究表明,饲粮中添加30 mg/kg的GOD(活性为10 000 U/g)显著降低了临武鸭血清MDA含量,添加水平为40 mg/kg时血清GSH-Px活性显著增加,证实了GOD具有增强机体抗氧化能力的作用[23]。本研究中,饲粮中添加350和500 mg/kg GOD均可使蛋种鸡血清和肝脏GSH-Px、SOD活性显著增加,MDA含量显著降低,血清T-AOC显著升高,与上述结果基本一致。由此可见,饲粮中添加350和500 mg/kg GOD能够提高产蛋后期蛋种鸡机体的抗氧化酶活性,减轻脂质过氧化水平,缓解其氧化应激状态,从而改善种鸡的产蛋性能、孵化性能和蛋品质。
3.4 GOD对产蛋后期蛋种鸡抗氧化相关基因表达的影响
Kelch样环氧氯丙烷相关蛋白1(Keap1)/Nrf2/抗氧化反应元件(ARE)信号通路是机体抵御氧化应激损伤的重要信号通路[24],该通路中起关键作用的是Nrf2,可调控下游多种抗氧化酶的基因表达[25]。Keap1是Nrf2的抑制因子,当细胞受到应激源刺激时,处于细胞质中的Nrf2与Keap1发生解偶联,并转移到细胞核与ARE结合,激活ARE相关二相酶的基因表达,降低细胞氧化损伤[26-27]。
Zhang等[28]研究表明,饲粮中添加含有GOD的益生菌后,仔猪肝脏Nrf2的mRNA和蛋白相对表达量显著增加。本研究中,饲粮中添加GOD显著提高了产蛋后期蛋种鸡肝脏Nrf2的mRNA相对表达量,同时Mn-SOD和Cu/Zn-SOD的mRNA相对表达量也显著增加,且血清和肝脏SOD活性表现出与肝脏抗氧化相关基因mRNA相对表达量一致的规律性。这说明GOD可诱导肝脏细胞中Nrf2/ARE信号通路关键基因Nrf2的表达,以调节下游SOD等相关抗氧化基因的转录和高效表达,使得机体抗氧化酶活性增加,最终提高机体抗氧化能力。然而,GOD是否可通过激活Keap1感应元件,调控Nrf2与Keap1解偶联来调控下游抗氧化基因表达以及GOD抗氧化作用的具体分子机制还有待进一步研究。
4 结 论
① 饲粮中添加350和500 mg/kg GOD可显著改善产蛋后期蛋种鸡的产蛋率和料蛋比,添加200、350和500 mg/kg GOD可显著改善种蛋受精率。
② 饲粮中添加350和500 mg/kg GOD可显著提高产蛋后期蛋种鸡种蛋的蛋壳厚度和哈氏单位。
③ 饲粮中添加GOD可提高产蛋后期蛋种鸡的血清和肝脏的抗氧化能力,激活肝脏细胞Nrf2/ARE信号通路相关基因的表达。
④ 综合经济效益分析,本试验条件下,产蛋后期蛋种鸡饲粮中GOD的适宜添加水平为500 mg/kg。
