系统性红斑狼疮中干扰素的表观遗传学研究进展
2021-11-04沈梦佳李志军吴华勋
沈梦佳,王 涛,李志军,吴华勋
(1. 蚌埠医学院第一附属医院风湿免疫科,安徽 蚌埠 233004;2.慢性疾病免疫学基础与临床安徽省重点实验室,安徽 蚌埠 233030;3.安徽医科大学临床药理研究所,安徽 合肥 230032)
系统性红斑狼疮(systemic lupus erythematosus,SLE)是一种多器官受累的自身免疫性疾病,以自身免疫耐受破坏、大量自身抗体产生为主要特点,临床表现多样,且主要影响育龄期女性[1]。SLE发病机制非常复杂,目前尚未完全研究清楚,但一般认为其与免疫学、遗传学、表观遗传学、环境因素和性别等因素有关。其中,多种环境因素共同作用可引起SLE的表观遗传学改变,异常的表观遗传修饰会造成免疫反应失调、自身免疫耐受性丧失,进而导致SLE的发生发展[1]。另外,干扰素(interferon,IFN)系统激活,IFN刺激基因(interferon-stimulated gene,ISG)在血液和组织中表达增加,即所谓的“IFN表达特征(IFN signature)”是SLE的特征之一[2]。近年来,研究人员们对SLE中的表观遗传学和IFN系统的关系展现出了极大的兴趣。因此,探索SLE中的IFN信号通路和异常表观遗传修饰之间的相互作用或许有一定价值,能为SLE的发病机制找到新的解释,并有助于发现SLE致病过程中更特异且更可靠的生物标志物。文中涉及的IFN信号通路及表观遗传修饰已尽可能地呈现在Fig 1中。
1 IFN家族介绍
IFN是一种广泛表达的细胞因子,最早是在鸡胚胎流感病毒实验中作为一种强烈干扰而被发现并因此得名。目前已知的IFN根据序列同源性可分为3个家族:Ⅰ型、Ⅱ型和Ⅲ型。人类Ⅰ型IFN家族主要成员是IFN-α和IFN-β,Ⅱ型IFN家族包括单一成员IFN-γ,Ⅲ型IFN家族的代表是IFN-λ[3]。经过长期研究,除了抗病毒活性,人们还发现IFN具有调节先天免疫和适应性免疫反应、抗肿瘤以及参与自身免疫性疾病中免疫应答失控的功能[3],而IFN-γ更为独特,它在促进病理性炎症和免疫调节中有着与Ⅰ型和Ⅲ型IFN不同的作用[4]。
Ⅰ型和Ⅲ型IFN可由几乎所有的有核细胞产生,尤其是巨噬细胞和树突状细胞(dendritic cell,DC)。大多数细胞都表达Ⅰ型IFN受体IFNAR,IFNAR又包括IFNAR1和IFNAR2,而Ⅲ型IFN的受体IFNLR主要分布在上皮细胞和浆细胞样DC(plasmacytoid DC,pDC)上[5]。Ⅱ型IFN主要由自然杀伤细胞(natural killer cells,NK)、NK T细胞和T细胞产生,IFN-γ通过独特的受体IFNGR传递信号,IFNGR在大多数类型的细胞上都有表达[5]。在模式识别受体(PRR)检测到微生物产物后,激活IFN调节因子(interferon regulatory factors,IRF)家族转录因子,后者又和NF-κB一起促进多种免疫细胞和非免疫细胞分泌IFN,之后,产生的IFN与细胞表面的受体结合,通过经典的Janus激酶(Janus kinase,JAK)/信号转导和转录激活因子(signal transducer and activators of transcription,STAT)通路启动信号级联反应,进而驱动ISG的转录[6]。由于3种类型的IFN诱导基因信号途径之间有显著的重叠,因此,我们很难通过基因信号来辨认具体是哪种IFN在发挥作用。
2 SLE中的IFN
以Ⅰ型IFN为例,在SLE中,由自身核酸过度激活的pDC是Ⅰ型IFN的主要来源,自身抗体和核酸结合蛋白组成的免疫复合物被pDC上的FcyRIIA吞噬后转运到内体,在那里免疫复合物的核酸部分与Toll样受体(toll-like receptor,TLR)7或TLR9结合,随后激活转录因子并产生IFN-α,这是SLE中Ⅰ型IFN持续产生的原因之一,另外,其他免疫细胞和pDC之间的相互作用也可促进IFN的持续产生[2],持续产生的IFN可以导致持续的自身免疫反应。IFN在SLE中通过多种方式发挥作用,包括通过IFN信号通路表达ISG、促使其他细胞发挥功能以及与其他细胞因子形成相互作用的调控网络等。以Ⅰ型IFN为例,IFN-α可促进Th17细胞的分化,抑制调节性T细胞(regulatory cell,Treg)的功能,使自身反应性T细胞扩增,它还刺激B细胞产生抗体。特别的是,IFN-α可增加NK细胞的细胞毒作用并促进IFN-γ产生,这说明Ⅰ型IFN和Ⅱ型IFN之间有所联系[2]。
关于SLE患者中IFN信号通路的研究一直是一个热点。据报道,三型IFN水平在SLE患者中均高于在健康对照组中,并且与SLE疾病活动指数(SLE disease activity index,SLEDAI)相关[7]。SLE患者的组织和器官,包括皮肤、关节、肾脏和中枢神经系统都有不同程度的IFN表达增加[2]。另外IFN还与部分SLE临床指标有一定相关性,例如,IFN-α水平与C3、IgG、ANA半定量、抗ds-DNA抗体半定量等临床指标的差异就具有统计学意义[8],但它们之间的相互作用的具体机制尚未可知。
随着对IFN信号的了解越来越深入,研究人员们尝试通过下调SLE患者中的IFN表达来治疗SLE,例如靶向IFN-α、IFNAR或IFNAR下游信号成分的治疗。其中,一种抗IFNAR的单克隆抗体anifrolumab刚刚完成了SLE患者的第三阶段临床试验,并显示出了令人满意的疗效和安全性[9],这表明在不影响抗病毒能力的前提下,选择性抑制IFN信号或许可以成为SLE患者个性化治疗的关键一环。
3 SLE中IFN的表观遗传调控
表观遗传修饰是指在不改变DNA序列的情况下基因表达的可遗传变化,主要包括DNA甲基化、组蛋白修饰和非编码RNA(non-coding RNA,ncRNA)等。目前SLE的估计遗传率为43.9%,然而同卵双胞胎SLE符合率仅为24%[1],这种SLE个体差异可以用表观遗传学解释,甚至,我们可以认为,表观遗传学因素大大增加了SLE发病机制的复杂性。部分异常表观遗传修饰参与调节SLE中的IFN信号,控制IFN相关基因的表达或抑制,进而参与调控自身免疫反应可能是表观遗传学因素在SLE中发挥作用的途径之一。
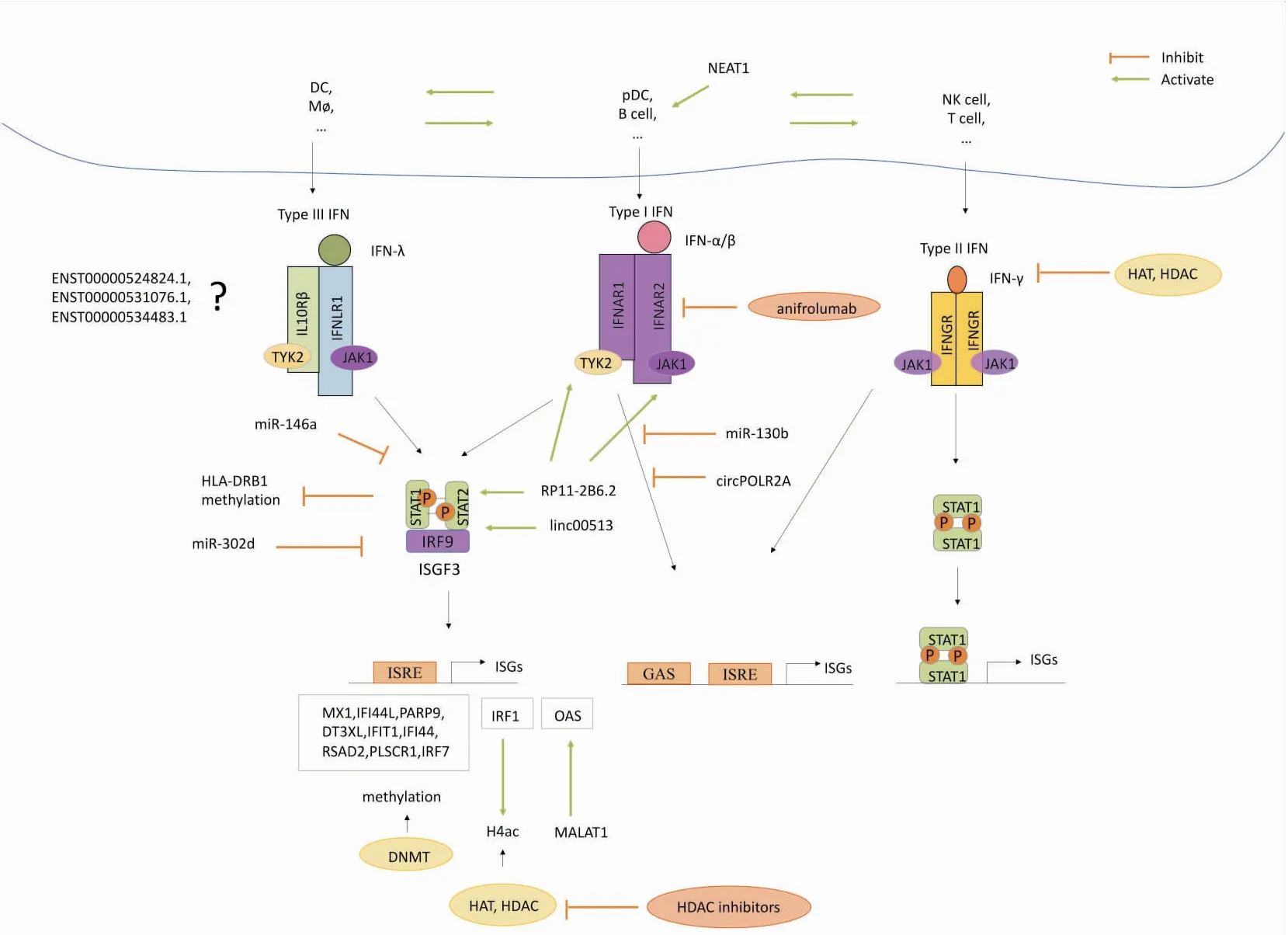
Fig 1 Potential mechanisms of IFN signaling pathway and epigenetic modification in SLE
3.1 DNA甲基化DNA甲基化是一种非常关键的表观遗传修饰,通常发生在CpG二核苷酸的胞嘧啶碱基的5位碳原子上,以S-腺苷甲硫氨酸作为甲基化的供体,由DNA甲基化酶(DNA methyltransferase,DNMT)催化完成。研究人员通过对比分析大量SLE患者和健康人的DNA甲基化图谱发现,SLE患者的结果表现出显著的低甲基化,这种低甲基化在活动期患者中尤为突出,值得注意的是,其中最显著的甲基化差异出现在Ⅰ型IFN相关基因上[10]。最近的一项研究显示,在SLE患者中,MX1、IFI44L、PARP9、DT3XL、IFIT1、IFI44、RSAD2、PLSCR1和IRF7等多个与Ⅰ型IFN通路有关的IFN相关基因存在显著低甲基化,且甲基化程度还与SLEDAI评分有关[11]。这些IFN相关基因在SLE中有各自的作用,筛选出这些基因的异常甲基化或许是有意义的,例如MX1和IFI44L,MX1水平的升高可以诱导适应性免疫反应,从而导致自身抗原的耐受性崩溃[11], 而IFI44L启动子甲基化更被认为是诊断SLE高度敏感且特异的标志物,可以用来鉴别SLE患者和健康人群与其他自身免疫性疾病如类风湿关节炎和原发性干燥综合征的患者,尽管IFI44L的具体功能还未可知,但可以确定的是表达增加的IFI44L是Ⅰ型IFN表达特征和免疫应答的组成部分之一[12]。另外,研究人员发现在狼疮患者CD8+T细胞中,HLA-DRB1也存在显著低甲基化,HLA-DRB1的表达依赖于CIITA和STAT1的信号转导,IFN-α处理可诱导体外狼疮CD8+T细胞表达CIITA和STAT1,因此狼疮中HLA-DRB1低甲基化发挥致病作用或许与狼疮患者体内高Ⅰ型IFN水平有关[13]。
3.2 组蛋白修饰组蛋白是一组与DNA链相关的保守蛋白质,它通过形成核小体来维持DNA链的稳定、复制和转录,以及改变基因的表达。组蛋白复合体的N端尾部延伸至核小体外,可以接受不同的化学修饰,主要包括甲基化、乙酰化、泛素化和磷酸化[14],其中关于甲基化和乙酰化的研究相对较多。组蛋白修饰水平在SLE中有所增加,并以不同的方式调节基因转录,异常组蛋白修饰的产生有赖于IFN过表达环境。研究人员在SLE患者单核细胞中发现H4赖氨酸残基乙酰化水平升高,他们认为可能的机制是IRF1直接或间接调节组蛋白乙酰化,另外体外细胞模型经IFN-α处理实验证明IFN-α的存在可以使IRF1被持续激活[15],所以SLE患者体内高IFN水平可能通过诱导IRF1过表达导致了H4高乙酰化。
组蛋白乙酰化由组蛋白乙酰化酶(histone acetylase,HAT)和组蛋白去乙酰化酶(histone deacetylase,HDAC)的相反作用共同控制,HAT和HDAC能够有效催化组蛋白赖氨酸残基上乙酰基的添加或去除[14]。其中HDAC可通过调控TLR、IFN信号通路和调控抗原提呈过程等途径参与调控固有免疫和适应性免疫过程,HDAC抑制剂是一类可抑制HDAC活性的化合物,被认为可以通过调控免疫细胞功能和抑制促炎细胞因子与趋化因子产生在SLE中发挥改善疾病严重程度和减少肾脏损伤的作用[16]。
3.3 非编码RNA转录组测序技术的发展推动了人类发现大量功能未知的非编码转录本,其中不具备编码蛋白质潜力的RNA被称为ncRNA,ncRNA主要包括微小RNA(microRNA,miRNA)、长链非编码RNA(long non-coding RNA,lncRNA)和环状RNA(circular RNA,circRNA)等。事实上,ncRNA占据了RNA的大多数,并且越来越能肯定的是,在各种细胞的生理功能中,ncRNA扮演着关键角色。例如,ncRNA参与调节Th1、Th17、滤泡辅助性T细胞( follicular helper T cell,Tfh)、Treg等细胞的分化和功能[17],而这些细胞可以分泌包括IFN在内的多种效应细胞因子,所以ncRNA同样可能是一种通过调节IFN信号参与SLE致病过程的表观遗传学因素。
3.3.1miRNA miRNA是一类长度约为22个核苷酸的ncRNA,主要通过沉默特定的mRNA来调节基因表达。miRNA在先天免疫和获得性免疫的发育、稳态功能中发挥作用,这些作用的失调将导致自身免疫反应和炎症。如前所述,SLE是一种Ⅰ型IFN依赖型自身免疫性疾病,pDC过度激活是其重要特点之一。一项研究发现SLE患者血浆中外泌体传递的miRNA是潜在的新型TLR7内源性配体,可以诱导pDC激活并促使其持续分泌Ⅰ型IFN,表明miRNA可能是启动自身免疫反应的新的致病介质和治疗Ⅰ型IFN介导疾病的潜在靶点[18]。
国内外研究显示[19-21],在SLE患者中,部分miRNA是IFN信号通路潜在的负性调节因子,例如miR-146a,miR-130b和miR-302d。其中,miR-146a缺陷可以诱导外周血单个核细胞(peripheral blood mononuclear cell,PBMC)分泌Ⅰ型IFN,此外,miR-146a靶向TLR7、IRF5和STAT1等多个Ⅰ型IFN通路中的关键信号蛋白[19],并且靶点还在不断更新中。而miR-302d是研究人员在SLE患者单核细胞中鉴定出的另一种靶向IRF9的miRNA,IRF9是调控ISG表达的转录复合物的关键成分,miR-302d表达降低可以使IRF9表达增加,进而导致MX1和OAS1在内的一些ISG的表达也增加,这种变化在疾病活动性增强的患者中尤为明显[21]。
3.3.2lncRNA lncRNA指的是一类长度超过200个核苷酸的ncRNA,其中包括在相邻蛋白编码基因间隙中转录而来的基因间长链非编码RNA (long intergenic non-coding RNA,lincRNA)。与miRNA相比,lncRNA可以通过更复杂的分子机制在多个水平上调节细胞内的基因表达[22]。研究人员们通过转录组测序等技术发现了大量在狼疮患者和健康人中差异表达的lncRNA,借助生物信息学技术,他们从中筛选出了可能参与IFN信号通路的lncRNA,例如ENST00000524824.1, ENST00000531076.1和ENST00000534483.1,这3种lincRNA可能通过调控IFN家族及其免疫调节通路在SLE的发病机制和疾病活动性中发挥重要作用[23]。而另一个lincRNA linc00513在SLE患者体内明显过表达,它是Ⅰ型IFN通路的正性调节因子,通过促进关键转录因子STAT1和STAT2的磷酸化放大IFN信号[24]。
MALAT1是一个较为经典的lncRNA,它在肿瘤和众多疾病中的作用都有相关研究,而在SLE的研究中,它被证明可以上调OAS2、OAS3和OASL等下游效应分子的表达,而OAS2、OAS3和OASL在无病毒和细菌存在的情况下介导了IFN-α-2a的促炎作用,从而参与SLE的致病过程[25]。另一个经典lncRNA NEAT1主要在人单核细胞中表达,SLE患者体内NEAT1的表达明显上调,且与病情严重程度相关,最近,研究人员通过狼疮小鼠模型发现,NEAT1的过度表达会使髓源性抑制细胞G-MDSC分泌B细胞激活因子(BAFF),从而促使B细胞中Ⅰ型IFN信号激活,诱发SLE患者B细胞中的IFN表达特征,最终导致SLE自身抗体的产生[26]。
lncRNA还参与了狼疮患者的器官损伤,研究人员发现在狼疮肾炎患者肾活检组织中RP11-2B6.2的水平有所升高,并且与疾病活动度和IFN评分呈正相关,RP11-2B6.2可以提高染色质可及性并抑制SOCS1基因转录,通过激活JAK1、TYK2和STAT1的磷酸化对Ⅰ型IFN通路起正向调节作用[27]。
3.3.3circRNA 近几年circRNA的研究热度有增无减,它是一种具有闭合环状结构的内源性ncRNA。在SLE等自身免疫性疾病中,部分circRNA充当miRNA的分子海绵,通过与miRNA相互作用参与基因调控,例如一项研究发现,在SLE患者中circIBTK的表达有所下调,circIBTK可以通过与miR-29b结合来逆转miR-29b诱导的DNA去甲基化和AKT信号通路的激活,从而阻碍SLE疾病进展,所以circIBTK或许可以作为SLE的诊断和治疗靶点[28]。
与miRNA相比,circRNA有更强的稳定性,因此它被认为是疾病诊断、治疗及预后更理想的分子生物标志物。目前circRNA在SLE中的研究集中于其作为生物标志物的效价和充当miRNA海绵发挥调控基因表达作用,而关于它的表观遗传修饰功能和具体分子机制的研究仍十分空缺[29]。然而已有研究组人员通过体外结合实验发现,circPOLR2A过表达可以导致先天免疫因子PKR活性降低和EIF2α磷酸化,并抑制Ⅰ型IFN诱导基因表达,因此SLE患者体内circRNA的大量降解或许可以解释SLE中先天免疫通路的异常激活和Ⅰ型IFN表达特征的高表达[30]。所以我们有理由相信,circRNA同样可以通过调节IFN表达特征,在SLE的发生发展中发挥表观遗传学功能,只是还需要大量更深入的研究。
4 结语
SLE临床表现多样并具有潜在致残性,令患者们深受其害。复杂的发病机制和高度异质性使SLE的临床治疗效果因人而异,并且往往伴有明显的副作用,因此我们期待未来有更为安全有效的治疗方法面世。近十几年来,越来越多的研究认可了参与调控IFN信号通路的表观遗传修饰在SLE疾病发生发展中的作用,但是大多数异常表观遗传修饰仍未揭开面纱,它们所造成的异常分子机制和信号转导通路变化都还需要更深入的研究。筛选出SLE患者和健康人免疫细胞亚型或组织中差异表达的表观遗传修饰,或将有助于进一步探索SLE的发病机制和发现SLE更特异、更可靠的生物标志物和治疗靶点。
