柴达木盆地唐古特白刺的表型多样性
2021-11-02吉德娟张得芳
吉德娟,张得芳,于 倩
(1.青海大学 农林科学院,青海 西宁 810016;2.青海高原林木遗传育种实验室,青海 西宁 810016)
唐古特白刺Nitraria tanguticumBobr.属蒺藜科白刺属落叶灌木,又名酸胖、唐古特泡泡刺,是多年生的旱生植物,主要分布于陕西、内蒙古、宁夏、甘肃、青海、新疆及西藏的荒漠和半荒漠,生于湖盆沙地、盐碱地和戈壁滩[1-2],青海省的白刺资源主要分布在德令哈市的尕海镇、怀头他拉镇、柯鲁克湖、大柴旦、都兰县的诺木洪、香日德农场、格尔木市的大格勒等地方,总面积10×104hm2,年产鲜果34×104t[3-4],是荒漠和半荒漠地区的建群树种,也是我国的特有灌木种,具有抗旱、抗寒、抗热、抗风沙、耐盐碱、耐贫瘠等生态适应特性,在稳定沙漠、治沙造林、保护绿洲中起到比较明显的生态作用[5-6]。唐古特白刺的果实是沙漠中罕见的野生浆果,有“沙漠樱桃”之称,果实中含有丰富的维生素、胡萝卜素、氨基酸、果胶、黄酮素、花青甙、多糖类、萜类、生物碱和多种微量元素等[7],具有健脾胃、滋补强壮、调经活血、抗氧化的功效,还具有降血压和降血糖的作用[8-10],对机体免疫、细胞增殖以及核酸、蛋白质的合成等都有直接或间接的积极作用[11]。白刺的叶片含有丰富的蛋白质和氨基酸[11],可作为饲养牲畜的优良饲料。白刺枝条可作为燃料和防风固沙的材料。白刺籽油含有丰富的人体必需的不饱和脂肪酸(EFA),含量高达96%以上,可开发制成保健品[12],另外白刺根部寄生的锁阳,在防癌、抗癌、延缓衰老、防治心血管疾病等方面也具有重要医疗价值和经济价值[13-15]。
近年来,随着白刺产品给人们带来越来越多的经济效益,人类对资源开始掠夺性的开采,另一方面由于植被退化、地下水资源枯竭、土壤盐渍化加重、沙漠化[16]等一系列问题使得野生唐古特白刺退化严重、生长不良,造成资源量大幅下降,野生白刺资源亟待保护。而对其资源的多样性研究是科学地加以保护的前提。
表型多样性的研究有着简单直观的特点,被广泛的应用于遗传多样性的研究中。随着对白刺资源开发程度的加大,近年来对白刺的研究随之增加,但是对唐古特白刺的表型多样性的研究报道还比较少,通过对唐古特白刺的叶片、果实、种子等形态学表型特征的系统研究,了解其表型分化程度,地理变异规律以及居群间和居群内的变异特征,对唐古特白刺资源的合理开发利用提供指导。
1 材料与方法
1.1 试验材料
通过2020年8月对柴达木盆地的唐古特白刺野生天然资源的调查,进行样品采集。采样点自然地理情况如表1所示,样点的 6 个群体基本覆盖了柴达木盆地野生唐古特白刺的分布区,在选定的居群内,每个居群随机选取15~30 株生长健壮、无明显病虫害的植株个体,距离间隔30 m以上,以确保采样的科学性,在选取的采样植株的东南西北4 个方向各取一定数量的叶片(统一选取相同部位的叶片),果实带回实验室。

表1 柴达木盆地唐古特白刺天然居群地理分布Table 1 Geographical distribution of Nitraria tanguticum Bobr. natural population in Qaidam basin
1.2 测定指标及方法
随机选取没有病虫害和机械损伤的完整叶片、果实以及种子的各性状进行测定。每个样本植株随机选取30 片叶片、30 个果实以及30 粒种子进行测定。
1)叶片性状测定:用游标卡尺(GB/T 21389—2008)测定叶长、叶宽、叶厚,用电子天平(0.001g)称取叶片鲜质量,并计算叶形指数(叶形指数=叶长/叶宽)。
2)果实性状测定:用游标卡尺测定果实横径、果实纵径,用电子天平称取鲜果单果质量,晾晒后称取干果单果质量,并计算果形指数(果形指数=果长/果径)。
3)种子性状测定:对采集的样本进行脱粒、净种后,随机抽取种子测定种子千粒重。
1.3 数据分析
利用Excel 2019 和SPSS 23.0 软件对数据进行方差分析、变异系数分析、相关性分析、主成分分析、聚类分析等。
2 结果与分析
2.1 唐古特白刺天然居群表型性状变异特征分析
对所调查的植株表型性状进行多重比较以及计算各性状平均值和标准差(表2),由表2可知,居群中14 个表型性状间存在显著的差异。株高平均值最大的是居群5,最小的是居群1,居群2、3、6 与居群2、4、5 之间没有显著差异;南北冠幅最大的也是居群5,最小的是居群1,居群5、6与居群1、2、3、4 之间存在显著差异;东西冠幅的多重比较以及最大、最小值,与南北冠幅之间的结果大体相同;
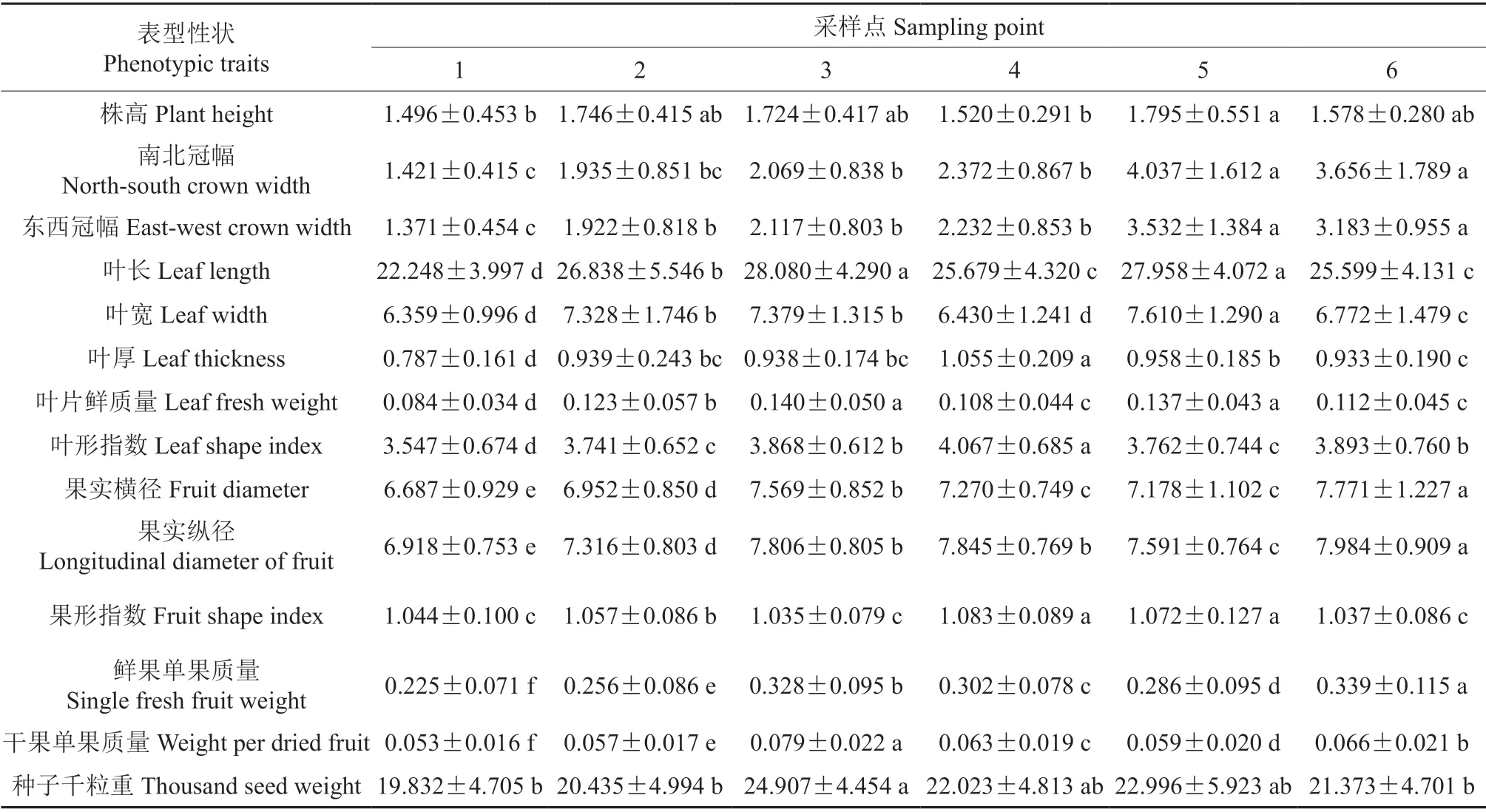
表2 各居群表型性状均值、标准差及多重比较†Table 2 Mean value,standard deviation and multiple comparison of phenotypic traits in each population
叶片各性状中,叶长均值最大的是居群3,最小的是居群1,居群3 和5、居群4 和6 之间的差异不显著;叶宽均值最大的是居群5,最小的是居群1,叶宽均值分布在6.359~7.61 mm 之间;叶片厚度的差异在各群体中表现比较明显,叶厚平均值最大的是居群4,最小的是居群1,居群4、居群1 与其他居群间均存在显著差异;叶片鲜质量在居群3 中的平均值最大,但与居群5 之间没有显著差异,与其他居群之间差异显著,叶片鲜质量平均值最小的是居群1。
果实的各性状在居群间表现出较大的差异,果实纵、横径均值最大的都是居群6,最小的都是居群5,居群间差异显著;果形指数平均值最大的是居群4,但与居群5之间没有显著差异,居群1、3、6 的果形指数之间也没有显著的差异性,但与其他居群间的差异显著;在鲜果与干果的单果质量方面,每个居群两两之间都存在显著的差异,鲜果单果质量最大的是居群6,而干果单果质量最大的是居群居群3,最小的都是居群1,其他居群的干果和鲜果单果质量呈对应关系。
种子千粒重最大的是居群3,最小的是居群1,居群1、2、6 与居群3 之间的差异达到了显著程度,与居群4 和居群5 之间的差异不显著。
综上结果发现,居群1 的各个指标均为最小值或与最小值之间没有显著差异,因此与其他居群在表型上产生明显的差异。
柴达木盆地唐古特白刺14 个表型性状在居群间、居群内两个层次上的变异均方及F 值见表3。经F 检验,株高、南北冠幅、东西冠幅、叶长、叶宽、叶厚、叶片鲜质量、叶形指数、果实横径、果实纵径、果形指数、鲜果单果质量和干果单果质量、种子千粒重在居群间都存在显著差异(P<0.05)。F值的大小是在一定程度上反映了不同表型性状在居群间差异程度的不同。居群间不同性状的F值由大到小依次为果实纵径、果形指数、干果单果质量、叶长、叶片鲜质量、叶厚、果实横径、叶宽、叶形指数、鲜果单果质量、南北冠幅、东西冠幅、种子千粒重、株高。由此可见,唐古特白刺居群间表型性状存在广泛差异,其中差异程度最大的分别是果实纵径和果形指数。
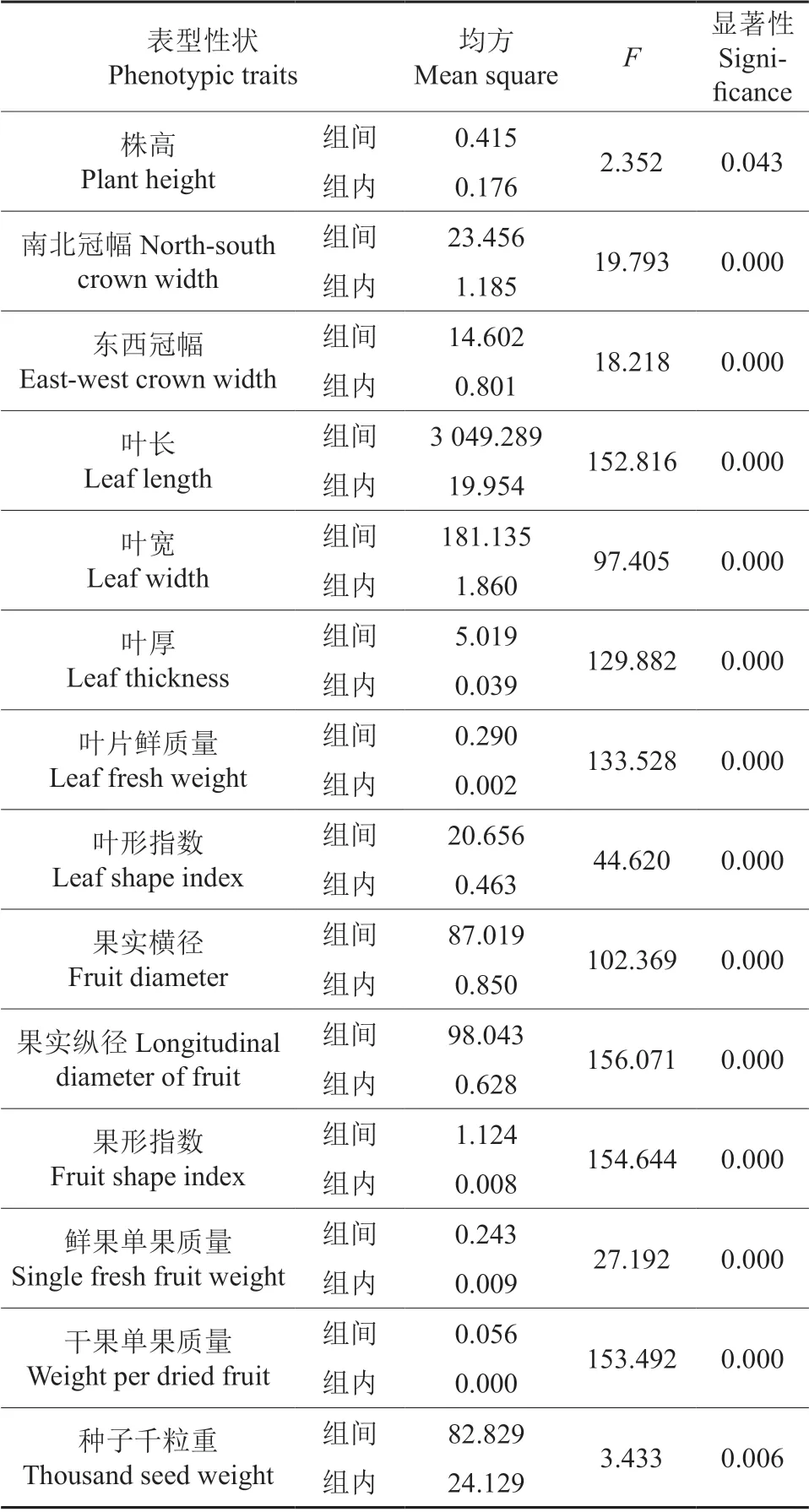
表3 唐古特白刺居群间和居群内方差分析Table 3 Analysis of variance between and within populations of Nitraria tanguticum Bobr.
2.2 唐古特白刺天然居群表型性状变异系数分析
唐古特白刺6 个天然居群表型性状的变异系数见表4,从表4中可以看出,唐古特白刺14 个表型性状的平均变异系数为23.69%,变异幅度在7.67%~48.95%之间,变幅比较大。同一个性状在不同生态区内的变异系数不尽相同,平均变异系数较大的为植株的南北冠幅和叶片鲜质量的变异系数,均值达到了39%以上,平均变异系数最小的是果形指数(CV=8.97%),说明果形指数较其它性状相对稳定。由表4可以看出变异系数由大到小的性状排列顺序为:南北冠幅、叶片鲜质量、东西冠幅、鲜果单果质量、干果单果质量、株高、种子千粒重、叶厚、叶形指数、果形指数。株高变异系数最大的是居群5,最小的是居群6,二者相差12.95%;南北冠幅在6 个居群间的变异范围为29.29%~48.95%;东西冠幅在各居群间的变异系数也较大,都在30%以上,最大的是居群2,最小的是居群6;叶厚的变异较植株的冠幅的变异程度要小,变异系数的范围在18.56%~25.83%之间;叶片鲜质量变异系数最大的是居群2,最小的居群5;叶形指数变异系数主要分布在18.07%上下,各居群之间的变异系数差别不是很大;果形指数的变异系数分布在7.67%~11.87%之间;鲜果单果质量和干果单果质量的变异系数分布范围分别为25.89%~33.89%、27.47%~34.26%;种子千粒重变异系数最大的是居群5,最小的是居群3。
唐古特白刺同一居群间各性状的变异也存在较大的差别,由表4可知,在居群2 中各性状的表型性状平均变异系数最大,居群4 表型性状平均变异系数最小。在各居群中,变异系数最小的性状都是果形指数,在居群1、2、4 中,变异系数最大的性状是叶片鲜质量;居群3、5、6 中,变异系数最大性状是南北冠幅。

表4 唐古特白刺不同居群表型性状变异系数分析Table 4 Coefficient of variation of phenotypic traits in different populations of Nitraria tanguticum Bobr.
2.3 唐古特白刺天然居群相关性分析
2.3.1 唐古特白刺天然居群表型性状相关性分析
对唐古特白刺的14 个表型性状进行双变量相关性分析,结果如表5所示。从表5可以看出在唐古特白刺各表型性状中,株高与叶片的各性状之间呈显著的相关关系;南北冠幅和东西冠幅彼此之间存在极显著关系,与其他表型性状之间的相关性都不显著;叶长、叶宽、叶厚也仅与叶片的部分性状呈显著的相关关系;叶形指数与果实纵径间存在显著的相关关系;果实横径和纵径都与鲜果的单果质量之间的相关性达到了极显著水平;果实的鲜果质量和干果质量之间存在显著的相关关系,干果质量还与种子千粒重之间呈显著的相关性。
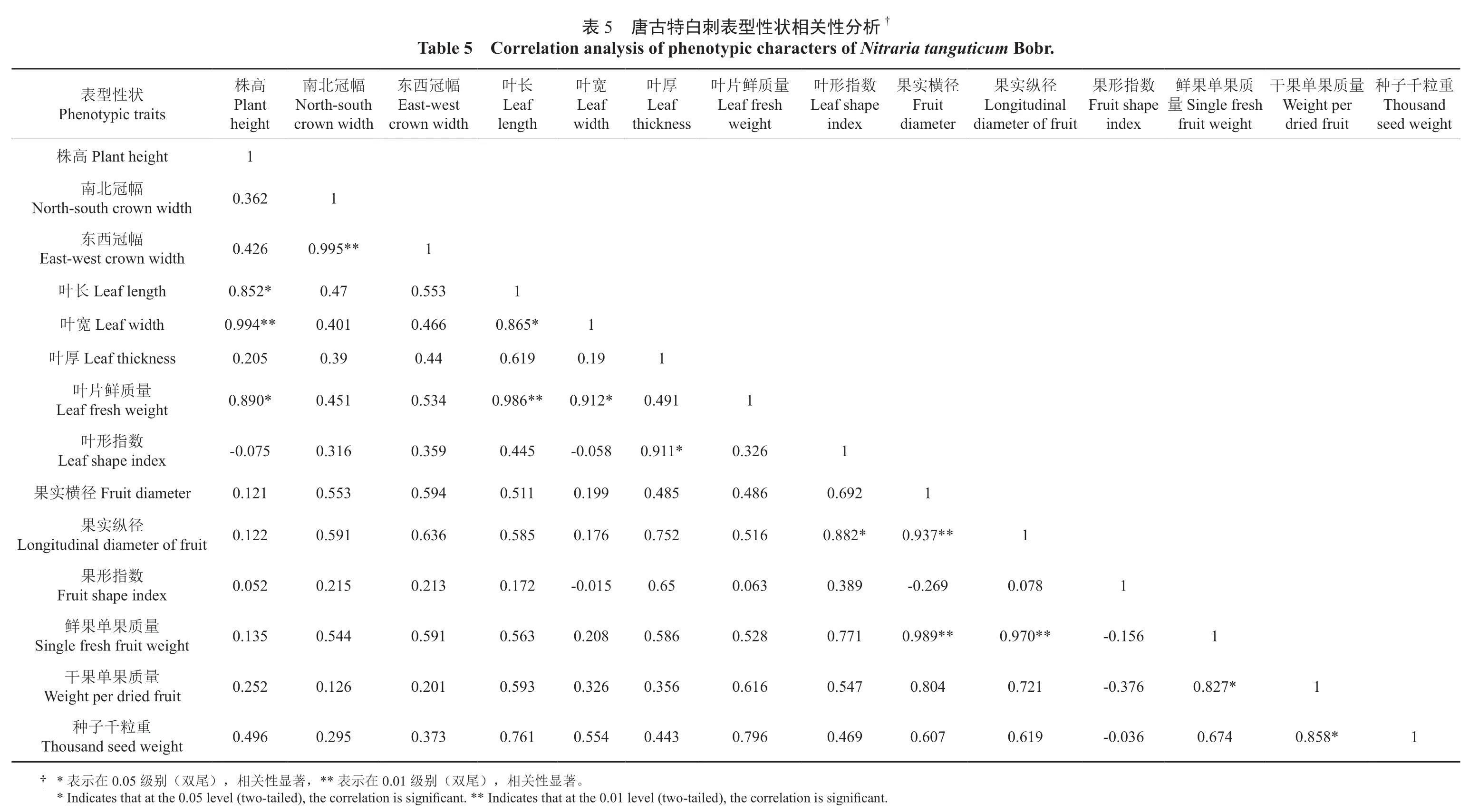
†析分性关相状性型表刺白特古5 唐表Table 5 Correlation analysis of phenotypic characters of Nitraria tanguticum Bobr.重子Thousand粒千种seed weight量果Weight per质果单dried fruit干果Single fresh质果单鲜量fruit weight果Fruit shape数指形index径纵实果Longitudinal diameter of fruit径实Fruit横果diameter叶Leaf shape数指形index叶Leaf fresh量质鲜片weight厚Leaf叶thickness宽Leaf叶width长Leaf叶length东East-west幅冠西crown width南North-south幅冠北crown width株Plant高height 1状性型表Phenotypic traits Plant height 高株1 0.362幅冠北南North-south crown width 1 0.995**0.426幅冠西东East-west crown width 1 0.553 0.47 0.852*Leaf length 长叶1 0.865*0.466 0.401 0.994**Leaf width 宽叶1 0.19 0.619 0.44 0.39 0.205 Leaf thickness 厚叶1 0.491 0.912*0.986**0.534 0.451 0.890*量质鲜片叶Leaf fresh weight 1 0.326 0.911*-0.058 0.445 0.359 0.316-0.075数指形叶Leaf shape index 1 0.692 0.486 0.485 0.199 0.511 0.594 0.553 0.121 Fruit diameter 径横实果1 0.937**0.882*0.516 0.752 0.176 0.585 0.636 0.591 0.122径纵实果Longitudinal diameter of fruit 1 0.078-0.269 0.389 0.063 0.65-0.015 0.172 0.213 0.215 0.052数指形果Fruit shape index 1-0.156 0.970**0.989**0.771 0.528 0.586 0.208 0.563 0.591 0.544 0.135量质果单果鲜Single fresh fruit weight 1 0.827*-0.376 0.721 0.804 0.547 0.616 0.356 0.326 0.593 0.201 0.126 0.252量质果单果干Weight per dried fruit 1 0.858*0.674-0.036 0.619 0.607 0.469 0.796 0.443 0.554 0.761 0.373 0.295 0.496重粒千子种Thousand seed weight。著显性关相),尾双(别0.01 级在示,** 表著显性关相),尾双(别0.05 级在示† * 表* Indicates that at the 0.05 level (two-tailed), the correlation is significant.** Indicates that at the 0.01 level (two-tailed), the correlation is significant.
2.3.2 唐古特白刺表型性状与地理因子间的相关性分析
对唐古特白刺的14 个表型性状和5 个地理因子的均值进行相关性分析,结果如表6所示。经度仅与叶形指数之间存在显著的相关关系;纬度与植株的南北、东西冠幅之间存在显著的相关关系;海拔与叶片的厚度之间的相关性达到极显著水平,与叶形指数、果实纵径之间的相关性达到显著水平;土壤pH 值与叶片厚度呈显著的负相关关系,与其他表型特征之间的相关性不显著;总的来说,海拔对唐古特白刺表型的影响略大于经纬度以及土壤的pH 值。从表型性状方面来看,环境因子对唐古特白刺表型性状影响最大的性状叶厚和叶形指数。

表6 唐古特白刺表型性状与地理因子间的相关性分析†Table 6 Correlation analysis between phenotypic traits and geographical factors of Nitraria tanguticum Bobr.
2.4 唐古特白刺表型性状的主成分分析
对唐古特白刺的14 个表型性状进行协方差主成分分析,结果如表7所示。对14 个表型性状进行降维,最终得到3 个主成分,其贡献率分别为75.602%、13.940%、8.379%,总贡献率累积达到97.921%,从图1也可以看出,从第4 个成分开始,特征值逐渐趋于平缓,说明前3 个主成分因子完全可以代表原始因子所代表的大部分信息。在第1个主成分中,公因子与原始变量之间的相关系数较大的有叶长(2.075)和种子千粒重(1.639),说明主成分1 的主要反映的是叶片和种子的性状,在第2 个主成分中,影响较大的原始变量主要是南北冠幅(0.81)、种子千粒重(-0.64)、东西冠幅(0.598),说明主成分2 主要由唐古特白刺的冠幅来体现,第3 个主成分中,与原始变量之间的相关系数较大的是叶长和种子千粒重,但是主成分3 与叶长之间呈反比。总体来看,叶长、种子千粒重以及植株的冠幅是对主成分分析影响较大的几个表型性状。
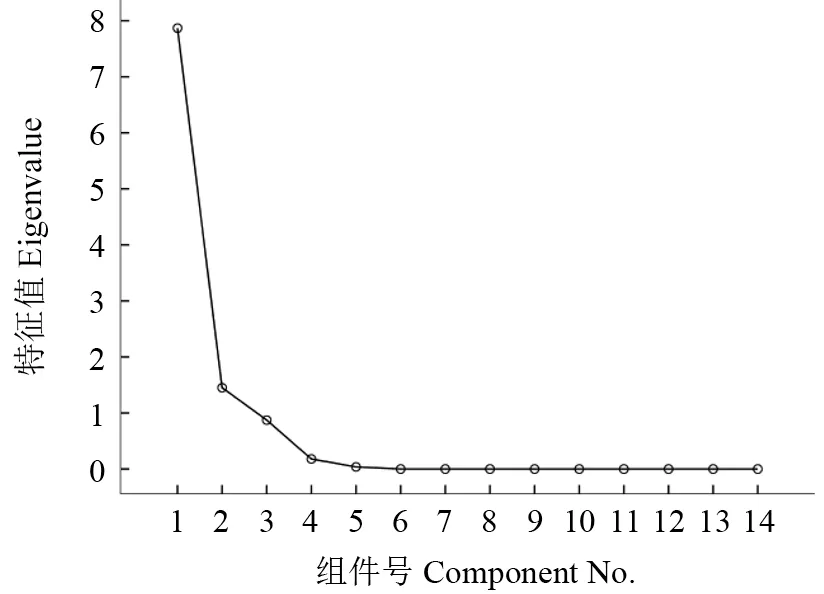
图1 唐古特白刺表型性状主成分分析组件图Fig.1 Component diagram of principal component analysis for phenotypic traits of Nitraria tanguticum Bobr.

表7 唐古特白刺表型性状前3 个主成分特征值及相关系数分析表Table 7 Characteristic values and correlation coefficients of the first three principal components of phenotypic traits of Nitraria tanguticum Bobr.
2.5 唐古特白刺野生居群表型性状聚类分析
利用平方欧式距离法对6 个野生唐古特白刺居群进行聚类分析,结果如图2所示,当平方欧式距离为5 时,将6 个天然居群分为三类,居群1为一类,居群3 和5 为一类,其他居群为一类;当欧式距离为15 时,居群1 为一类,其他居群聚为一类。

图2 基于14 个表型性状的唐古特白刺野生居群聚类图Fig.2 Cluster diagram of Nitraria tanguticum Bobr. wild population based on 14 phenotypic traits
3 讨论与结论
3.1 讨 论
通过表型性状特征来评价植物遗传多样性是一种简单、便捷的方式,且便于野外资源的调查与收集[17],表型性状反映了基因在长期环境作用下产生的不可逆的变化,这种变化经过稳定遗传后产生新性状[18],对植物的进化和适应多变环境具有重要意义[19]。因此很多学者都利用形态学标记研究植物的多样性,陈霞等[20]、唐燕等[21]、童跃伟等[22]都利用果实和叶片的性状分别对川梨、枸杞和红松的表型多样性进行了分析,也有学者通过枝、果、花、叶和种子[23]等部位的形态指标对植物的表型多样性进行了研究。将表型指标与分子标记相结合进行优良品种选育,在很大程度上缩短育种周期,同时也可用于资源保护,为植物资源鉴定和保护研究提供科学依据[24]。唐古特白刺是分布于荒漠和半荒漠地区重要的防沙治沙树种,由于其生长环境的多样性,在基于果实、叶片、种子的表型性状多样性研究中再加入植株冠幅和树高等性状来研究唐古特白刺的表型多样性,使研究结果更加准确可靠。
天然分布下的野生居群一般都具备丰富的遗传变异,并且对生长地方特殊的生境条件更加适应[25]。生境的变化会通过一系列级联反应进而影响生物多样性[26],探究植物和环境之间的相互作用,有助于了解植物进化过程和适应环境机制。本研究中,分布于不同地理环境的唐古特白刺在居群间和居群内的性状表型值,变异系数之间存在显著的差异性,这是唐古特白刺自身的遗传因素以及长期的环境异质性共同作用的结果。一般认为,当变异系数大于10%时,说明植物的性状差异很大[27],变异系数越大,植物对环境的适应广度也就越大[28]。在本研究中,表型性状变异系数的范围在7.67%~48.95%之间,说明唐古特白刺居群间存在丰富的表型多样性,且具有在多变的环境中适应的能力。实验所调查的唐古特白刺多分布于荒漠半荒漠以及河滩等地,生长于荒漠半荒漠的唐古特白刺长期受风沙侵蚀的影响,而生于河滩等地的唐古特白刺所受到风沙的影响较小,风沙会影响植物的光合速率和蒸腾速率[29],进而影响植物的生长。本研究中,分布于不同生境下的唐古特白刺的东西冠幅、南北冠幅、叶片鲜质量以及单果重等性状在不同居群之间存在显著差异,主要是受风沙大小以及土壤理化性质的影响,这些性状也是唐古特白刺重要的表型性状,这与张亚琦等[30]研究青海西伯利亚白刺的表型多样性中的结论具有一致性。
表型性状的相关性的研究中,各性状间存在一定的相关性,在后续的研究中,可以通过一种性状间接选择另一种性状,间接的进行优良单株的早期筛选。在本研究中,共存在14 对显著的相关关系,多数为叶片与叶片、种实与种实之间的相关性,叶片与种实之间的相关性较少,这与王宁等[31]研究的元宝枫表型多样性的结果相同。本研究中仅有叶形指数与果实纵径呈显著正相关,推测可能与居群的气候环境复杂多样有关,使得唐古特白刺的生殖生长受营养生长的限制性较小。植物长期对环境的适应以及响应,使其与环境之间产生密切的联系,研究他们之间的联系,有利于揭示植物的变异格局,植物叶片大小受温度、水分以及风速的影响,有研究表明,灰木莲的叶形指数和叶周长随着经度的增加而增大[32],在本研究中也得到了相同的结果。叶形指数和叶厚还与海拔之间存在显著的正相关关系,说明海拔越高,叶片将越窄越长,叶片也越厚,这可能与唐古特白刺长期适应寒冷、风沙的气候特征有关。冠幅与纬度呈显著正相关,这与刘明宣等[33]研究的枫香地理种源变异中的结果一致。
唐古特白刺的生物性状较多,因此在对其研究的过程中可能存在信息重叠的情况,主成分分析是对多个性状进行综合分析[34],以便得到较为简便的结果,同时可以排除由于原指标间的相关关系对分析结果产生的干扰[35],直观揭示群体间的遗传差异,有效指导目标亲本选育,有利于种质资源的分类[36]。在本研究中,唐古特白刺的14个性状中提取了3 个主成分,累积贡献率达到了97.921%,综合分析,对唐古特白刺表型多样性起主要作用的性状是冠幅、叶长以及种子的千粒重,这与前面所研究的环境影响变异的性状的结果有一定的相似性。张亚琦[37]对白刺属植物表型多样性的研究中发现,引起西伯利亚白刺表型变异的性状分别是叶形系数、每序核果数、果序重和果形系数,引起齿叶白刺表型变异的性状有种子千粒重、生长量、果序重和核形系数。
聚类分析可以探究不同种质间的亲缘关系和起源,同时为遗传多样性在分子水平上的研究提供一定的理论基础[38]。本研究中6 个唐古特白刺居群聚为2 类,居群1 为一类,其他居群为一类,分析发现,居群1 的土壤pH 值为8.38,高于其他几个居群的土壤pH 值,较高的土壤pH 值条件下植物受到离子胁迫、渗透压力胁迫以及高pH 值的影响,最直观的反应是影响其组织与器官。由于居群1 的土壤pH 值、土壤电导率较其他居群的要高,其叶长、叶宽、冠幅等指标均变小,并逐渐区别于其他群体中的植物个体,故居群1 最终单成一类,其他5 个居群的地理环境因子以及表型性状之间的都有一定的相近性,故为一类。
表型多样性在一定程度上可以反映遗传多样性的情况,但其受环境影响较大,因此在某些情况下不能准确的反映遗传多样性[39],本研究通过表型性状研究发现,柴达木盆地的唐古特白刺的种质资源丰富,后续研究中还需要结合分子层面进行更深一步的分析,以便更好地了解、开发利用和保护柴达木盆地唐古特白刺资源。
3.2 结 论
1)通过对唐古特白刺14 个表型性状的多重比较和方差分析表明,各个表型性状在不同居群间都表现出不同程度的差异性。在唐古特白刺的14 个表型性状中,植株的东西冠幅、南北冠幅、叶片鲜质量和果实的单果质量是变异较大的性状,各性状变异系数分布在了7.67%~48.95%之间。
2)唐古特白刺表型性状相关性分析表明,除东西冠幅、叶片鲜质量以及果形指数与其他性状间没有显著的相关关系外,其他性状之间都存在一定程度的相关性;表型性状与地理环境因子相关性分析中,地理环境因子主要影响唐古特白刺叶片各性状。
3)在唐古特白刺的14 个表型性状中,通过主成分分析,最终提取3 个主成分,累积贡献率达到97.921%,对主成分进行分析,结果显示东西冠幅、南北冠幅、叶长和种子千粒重是影响唐古特白刺的主要表型性状。
4)利用平方欧式距离将6 个唐古特白刺居群聚为2 类,居群1 单独为一类,居群2、3、4、5、6 为一类。
