基于小麦外引种质资源的粒质量基因分子标记检测及组合分析
2021-11-01吴晓军李奉原申云霞陈向东胡喜贵张雪宁李小利胡铁柱茹振钢
吴晓军,李奉原,申云霞,陈向东,胡喜贵,姜 豪,张雪宁,李小利,胡铁柱,茹振钢
(河南科技学院 小麦中心,河南省杂交小麦重点实验室,现代生物育种河南省协同创新中心,河南 新乡 453003)
小麦产量是穗数、穗粒数以及千粒质量三因素协调优化的结果,其中千粒质量的遗传力较大,较为稳定[1],增加千粒质量是小麦产量育种的主要目标之一。研究表明,粒质量是数量性状,受主效和微效多基因控制[2-3]。随着分子生物学和比较基因组学的发展,在水稻和小麦的粒质量形成分子机制方面取得了一些重要研究进展,许多粒质量相关数量性状位点被鉴定出来,一些粒质量基因被成功克隆,并开发了相关功能分子标记[4-5]。这些研究成果为分子标记辅助选择技术在小麦育种中的应用奠定了基础。
籽粒大小是粒质量形成的重要影响因素[6]。Su等[7]根据水稻粒宽基因OsGW2信息,在普通小麦中同源克隆得到TaGW2基因,该基因编码具有E3泛素连接酶功能的蛋白质,位于小麦第6同源染色体上并靠近着丝粒,在TaGW2-6A启动子区检测到的2个SNP,形成2种单倍型Hap-6A-A(-593A和-739G)和Hap-6A-G(-593G和-769A),以-593位A/G多态性为基础,设计了酶切扩增多态性序列(CAPS)标记以区分2个单倍型。对265个中国小麦品种进行基因型和关联分析发现,单倍型Hap-6A-A与小麦粒宽、千粒质量显著相关,能够使小麦千粒质量增加3 g以上,并且抽穗早、成熟早。利用不同研究群体对小麦粒宽和千粒质量相关QTL的定位结果也显示,在小麦染色体6AS的TaGW2-6A位点附近检测到了影响小麦粒宽和千粒质量的QTL[8-11],这些研究为TaGW2-6A对粒质量和千粒质量的调控功能提供了更多证据。
细胞壁转化酶(CWI)是库器官发育和碳分配的关键酶,与籽粒质量密切相关[12-15]。Ma等[16]在小麦2AL染色体上克隆得到细胞壁转化酶基因(TaCwi-A1),发现该基因有2种单倍型TaCwi-A1a和TaCwi-A1b,其中单倍型TaCwi-A1a与高千粒质量有关,单倍型TaCwi-A1b与低千粒质量有关。谷氨酰胺合成酶(GS)在谷类作物的生长、氮利用和产量潜力中起着关键作用[17-18]。Li等[19]从普通小麦中国春和小偃54的2A、2B和2D染色体上克隆了谷氨酰胺合成酶基因(TaGS2),对2D染色体上TaGS2-D1基因的编码区序列分析发现,有2种单倍型TaGS2-D1a和TaGS2-D1b,关联分析发现,TaGS2-D1a能够提高小麦的产量和氮素利用效率。小麦蔗糖合成酶基因TaSus-2B参与小麦淀粉合成及胚乳形成,并最终影响籽粒质量[20]。Jiang等[21]从小麦2B染色体上克隆到TaSus2-2B基因,其有2种单倍型TaSus2-2Ba和TaSus2-2Bb,关联分析表明TaSus2-2Ba能够显著增加小麦千粒质量。
当前对作物产量相关基因的发掘研究进展较快,而利用价值评估方面,尤其是多个粒质量相关基因的综合效应分析研究报道较少[4]。为此,本研究对48份国外引进小麦种质进行了千粒质量表型鉴定,并利用4个粒质量相关功能分子标记TaCwi-A1、TaGW2-6A、TaGS2-D1、TaSus2-2B进行粒质量基因单倍型类型和分布检测,以评价粒质量基因功能分子标记的有效性及4个粒质量基因不同单倍型组合的综合遗传效应,筛选具有较好粒质量性状遗传改良潜力的小麦种质,为粒质量基因功能分子标记和外引种质在国内小麦育种中的利用提供一些科学依据。
1 材料和方法
1.1 试验材料
参试材料包括48份国外小麦种质,分别引自智利、日本、墨西哥、法国等10多个国家,其中Kanto107、Manital、Opata、Pastor、Norin61、Salmone、Glenlea、RL6077、Madsen、Attila、Pavon共11份材料,由山东省农业科学院作物研究所资源库提供,其他材料由河南科技学院小麦中心保存和提供。
1.2 粒质量相关性状测定
小麦品种(系)于2019-2020年种植在河南科技学院朗公庙试验基地,随机区组设计,2个重复,行长4 m,行距25 cm,2行区,采用常规标准试验田管理方法,适时收获,脱粒。从每个品种(系)中数取500粒小麦种子,3次重复,分别称质量,并计算千粒质量平均值。
1.3 DNA提取和分子标记检测
在每个品种(系)中取3株小麦的叶片混合,采用Lagudah等[22]方法提取其基因组DNA,并用1%的琼脂糖凝胶进行电泳检测。将DNA原液稀释至100 ng/μL工作液浓度,置于4 ℃冰箱保存备用。
利用Su等[7]开发的CAPS标记(Hap-6A-P1和Hap-6A-P2)检测TaGW2-6A基因的单倍型;利用Ma等[16]开发的一对显性互补功能标记(CWI21和CWI22)检测TaCwi-A1基因的单倍型;利用Li等[19]等开发的功能分子标记检测TaGS2-D1基因的单倍型;利用Jiang等[21]开发的一对显性互补功能标记(Sus2-1和Sus2-2)检测TaSus2-2B基因的单倍型。以上分子标记引物均由生工生物工程(上海)股份有限公司合成,引物详细信息见表1。PCR反应体系为20 μL,内含2×TaqMaster Mix(北京康为世纪生物有限公司)10 μL,上下游引物(10 μmol/L)各0.8 μL,模板DNA(100 ng/μL)1 μL,ddH2O 7.4 μL。PCR扩增反应程序:95 ℃ 预变性4 min;94 ℃变性30 s,55.0~61.5 ℃ 退火30 s,72 ℃延伸50 s,32~35个循环;72 ℃延伸5 min。TaGW2-6A(Hap-6A-P2)分子标记扩增产物在2%的琼脂糖凝胶上进行电泳检测,其余TaGW2-6A(Hap-6A-P1)、TaCwi-A1、TaGS2-D1、TaSus2-2B等分子标记扩增产物的电泳检测均使用1%的琼脂糖凝胶,电泳后在凝胶成像系统中扫描成像,拍照保存。

表1 分子标记的引物信息Tab.1 Primer information of molecular markers
1.4 统计分析
试验数据整理使用Excel 2016软件,不同粒质量基因及组合间千粒质量的差异显著性分析使用SPSS 25.0软件。
2 结果与分析
2.1 TaCwi-A1基因在供试小麦种质中的单倍型类型和分布
利用TaCwi-A1基因的一对显性互补功能标记(CWI21和CWI22)对48份小麦外引种质进行检测(图1),高千粒质量TaCwi-A1a单倍型的扩增条带为402 bp,低千粒质量TaCwi-A1b单倍型的扩增条带为404 bp,检测结果显示,1份小麦种质KPL-1未扩增出条带,37份小麦种质酶切得到402 bp特异性条带,占供试材料的77.1%,7份小麦种质酶切得到404 bp特异性条带,占供试材料的14.6%,3份小麦种质(Kanto107、Salmone和硬质博莱特)均扩增出402,404 bp 2个特异性条带,占供试材料的6.3%,为位点杂合型(表2)。

表2 48份外引种质中粒质量基因单倍型类型及千粒质量Tab.2 Haplotypes of grain weight genes and thousand kernels weight of 48 introduced germplasms
2.2 TaGW2-6A基因在供试小麦种质中的单倍型类型和分布
利用TaGW2-6A基因的CAPS分子标记(Hap-6A-P1和Hap-6A-P2)对48份外引小麦种质进行检
测(图2),首先用Hap-6A-P1引物扩增出949 bp条带,以之为模板,再用Hap-6A-P2引物进行扩增,扩增的产物经TaqⅠ限制性内切酶酶切,可切出167,218 bp条带,分别对应于高千粒质量、低千粒质量单倍型Hap-6A-A和Hap-6A-G。检测结果显示,1份小麦种质09智引2号未扩增出条带,27份小麦种质酶切出167 bp特异性条带,占供试材料的56.3%,20份小麦种质酶切得到218 bp特异性条带,占供试材料的41.7%(表2)。
2.3 TaGS2-D1基因在供试小麦种质中的单倍型类型和分布
利用TaGS2-D1基因特异性引物对48份小麦种质进行检测(图3),结果显示,1份小麦种质野猫未扩增出条带,32份小麦种质,为高千粒质量TaGS2-D1a单倍型,占供试材料的66.7%,15份小麦种质,为低千粒质量TaGS2-D1b单倍型,占供试材料的31.3%(表2)。
2.4 TaSus2-2B基因在供试小麦种质中的单倍型类型和分布
利用TaSus2-2B基因的一对显性互补功能标记(Sus2-1和Sus2-2)对48份小麦种质进行检测(图4),Sus2-1引物扩增出423 bp条带,Sus2-2引物扩增出381 bp条带,分别对应于高千粒质量、低千粒质量单倍型。检测结果显示,1份小麦种质CL0438未扩增出特异性条带,8份小麦种质扩增出423 bp条带,占供试材料的16.7%,为高千粒质量TaSus2-2Ba单倍型,39份小麦种质扩增出381 bp条带,占供试材料的81.3%,为低千粒质量TaSus2-2Bb单倍型(表2)。
2.5 基因单倍型及其组合对小麦千粒质量的影响
本研究中48份小麦外引种质材料的平均千粒质量为30.15 g,变异范围为20.13~43.19 g,超平均千粒质量种质材料占45.8%(表2)。各粒质量相关基因均包含2种单倍型,就单个基因2种单倍型的平均千粒质量比较来看(表3),TaCwi-A1基因高千粒质量单倍型TaCwi-A1a的平均千粒质量高于低千粒质量单倍型TaCwi-A1b,差异达到显著水平,且TaCwi-A1a在供试小麦种质材料中占比较高,为优势单倍型;TaGW2-6A基因的高千粒质量单倍型Hap-6A-A的平均千粒质量高于低千粒质量单倍型Hap-6A-G,差异达到显著水平,并且Hap-6A-A在供试小麦种质材料中的占比也高于Hap-6A-G,基本属于优势单倍型;TaGS2-D1基因的高千粒质量单倍型TaGS2-D1a在供试小麦种质材料中占比虽高于低千粒质量单倍型TaGS2-D1b,但是平均千粒质量却显著低于TaGS2-D1;TaSus2-2B基因的高千粒质量单倍型TaSus2-2Ba在供试小麦种质材料中占比不仅低于低千粒质量单倍型TaSus2-2Bb,其平均千粒质量也显著低于TaSus2-2Bb单倍型。
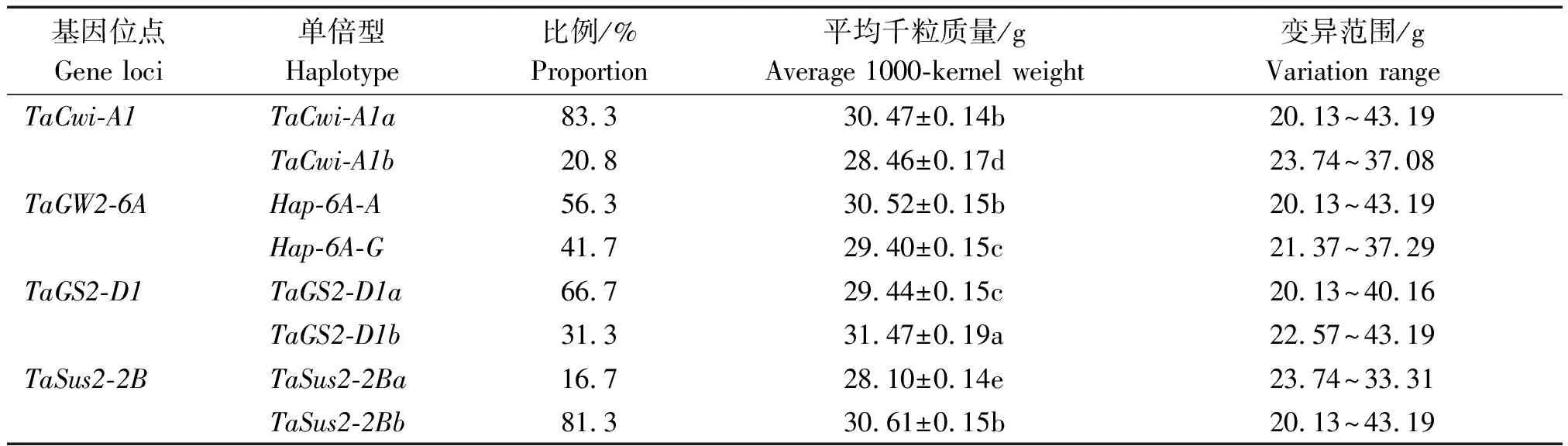
表3 不同单倍型对小麦种质千粒质量的影响Tab.3 Effects of different haplotypes on the 1000-kernel weight of wheat germplasms
4个千粒质量相关基因的单倍型组合中,以TaCwi-A1a/Hap-6A-A/TaGS2-D1a/TaSus2-2Bb在供试材料中占比最高,为33.3%,平均千粒质量为29.32 g,TaCwi-A1a/Hap-6A-G/TaGS2-D1b/TaSus2-2Ba在供试材料中占比最低,为4.2%,平均千粒质量为27.51 g(表4)。在单倍型组合中,TaCwi-A1a/Hap-6A-A/TaGS2-D1b/TaSus2-2Bb在供试材料中占比为10.4%,平均千粒质量最高,达到35.68 g,其次是TaCwi-A1a/Hap-6A-G/TaGS2-D1a/TaSus2-2Bb组合,在供试材料中占比为12.5%,平均千粒质量达到31.46 g。就总体趋势而言,含有相同单倍型组合的种质材料在供试材料中占比高的,其平均千粒质量也高,在供试材料中占比低的,其平均千粒质量也相对较低。
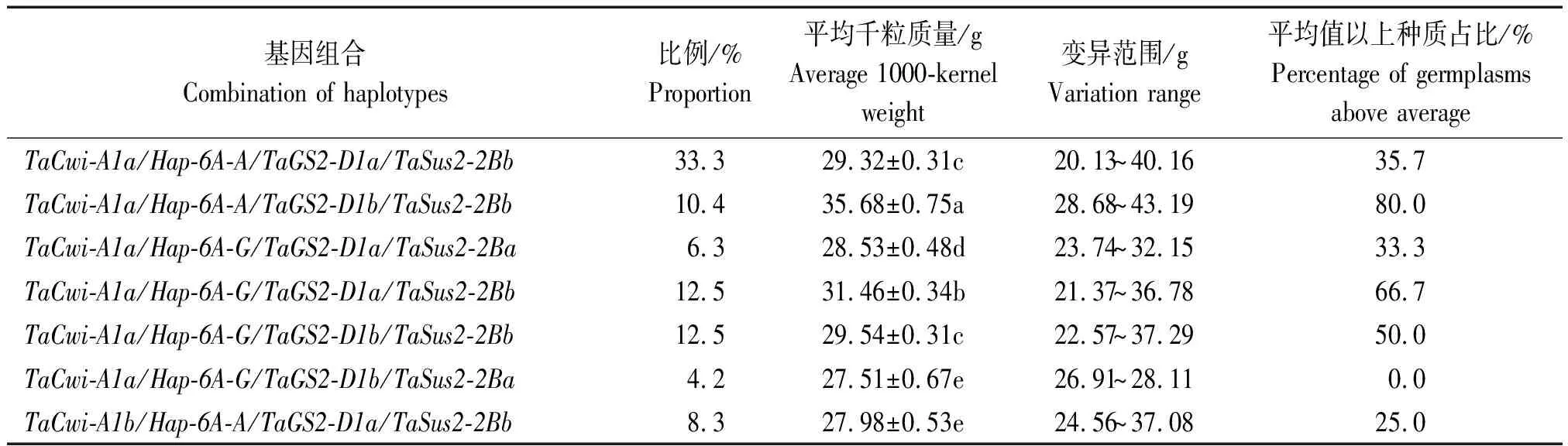
表4 部分单倍型组合对小麦种质千粒质量的影响Tab.4 Effects of some haplotype combinations on the 1000-kernel weight of wheat germplasms
3 结论与讨论
小麦粒质量在产量三因素中受遗传特性影响最大,与单位面积穗数和穗粒数无负相关关系,相对独立[23]。在稳定单位面积穗数和穗粒数的前提下,增加粒质量对提高小麦产量效果显著[24]。随着分子生物学和比较基因组学的发展,很多小麦粒质量相关基因被克隆出来,并基于基因序列开发出功能分子标记,能够大大提高小麦分子育种效率。然而,其在育种实践中的利用价值评价,尤其是综合遗传效应评价还较少[4,25]。
寇程等[26]利用开发的TaGW2基因的CAPS标记对国内外316份小麦品种(系)进行单倍型类型检测,发现Hap-6A-A和Hap-6A-G类型在粒质量、粒长和粒宽上差异显著。相吉山等[27]研究表明,TaCwi-A1基因的分子标记可以较好地区分小麦千粒质量大小,能够应用于小麦粒质量的分子标记辅助育种。本研究结果显示,TaGW2-6A基因单倍型Hap-6A-A、TaCwi-A1基因的单倍型TaCwi-A1a均为高千粒质量等位变异,且与低千粒质量单倍型在粒质量上的差异达到显著水平,所得结果与前人[26-27]研究一致。表明TaGW2-6A和TaCwi-A1基因在不同环境下的遗传效应都较为稳定,分子标记与平均千粒质量表型的一致性较好,可以用于小麦育种中优异种质千粒质量的鉴定和筛选。TaSus2-2B基因单倍型TaSus2-2Ba在外引小麦种质中的平均千粒质量显著低于单倍型TaSus2-2Bb,这与Jiang等[21]的研究不一致。时佳等[28]对219份新疆小麦品种(系)的TaSus2-2B基因检测也显示,单倍型TaSus2-2Bb的平均千粒质量显著高于单倍型TaSus2-2Ba。其原因可能在于,小麦的蔗糖合成酶基因对小麦千粒质量有着重要影响,然而该基因又对环境条件变化较为敏感,从而造成了上述研究结果不一致的情况[28]。另外,在48份外引种质中TaGS2-D1基因的单倍型TaGS2-D1a占比高于单倍型TaGS2-D1b,而平均千粒质量显著低于单倍型TaGS2-D1b,这与宋晓朋等[29]的研究结果并不一致。一直以来,对小麦千粒质量的研究所用群体多为重组自交系、DH群体或者回交群体,这些群体多基于千粒质量差异较大的亲本衍生而来,因此具有遗传背景狭窄的缺点[30],并且在不同环境和遗传背景中的遗传效应研究还较少。在本试验中,48份小麦种质材料来自国外10多个国家,整体的遗传多样性非常高,分析认为,单个基因对粒质量相关性状的影响可能会被其他籽粒表型相关基因影响所掩盖,从而导致研究结果的差异。以上结果也表明,粒质量性状由多基因控制,对多个粒质量相关基因进行综合遗传效应分析是十分必要的,有利于在小麦分子育种实践中的有效应用。
对外引种质中4个粒质量基因的单倍型部分组合研究结果显示,含有相同单倍型组合的种质材料在供试材料中占比高的,其平均千粒质量也相对较高。表明这些国外引进的小麦种质资源本身具有较好的遗传基础,聚合了一些优异的粒质量相关基因,这对于拓宽国内小麦的遗传基础有一定作用。
通过对48份国外引进小麦种质的4种粒质量相关基因等位变异类型检测和千粒质量性状测定,鉴定出一些含有粒质量相关优异等位变异基因的小麦种质资源。在较高遗传多样性的情况下,验证了TaCwi-A1基因和TaGW2-6A基因的优异等位变异类型遗传效应的稳定性,以及发现TaSus2-2B基因和TaGS2-D1基因容易受到环境和其他粒质量相关基因影响的特点,提出对于粒质量相关基因进行综合遗传效应分析的必要性。本研究为外引种质的有效利用和分子标记辅助选择技术在小麦粒质量相关性状改良中的应用提供了一些科学依据。
