欧洲黑杨高生长相关基因PnIAA9的克隆及进化分析
2021-10-28杨成超
杨成超
(辽宁省杨树研究所,辽宁 盖州 115213)
杨树Populusspp.属于被子植物门Angiosperms双子叶植物纲Dicots杨柳科Salicaceae杨属,是多年生木本植物,无居间分生组织。它分布广泛,易繁殖,是人工用材林、生态防护林及四旁绿化的重要树种,具有重要的经济和生态价值。树高是与杨树木材产量密切相关的重要性状之一。由于多年生杨树树体高大,树高不易精确测定,故1年生苗的高生长研究在杨树高生长形态建成研究中占据重要地位[1]。
高等植物通过调节顶端分生组织和侧生分生组织的活性建立地上株型系统,分生组织的活性受环境信号、发育阶段和遗传因素的综合调控,植物激素参与这些信号的整合。顶端优势是植物分枝调控的核心问题,而生长素对顶端优势的形成和维持发挥关键作用。Auxin/indole-3-acetic acid(Aux/IAA)蛋白是调节许多对生长素反应现象的转录调节因子,大部分Aux/IAA的功能已经通过拟南芥获得性突变体确定了。Aux/IAA和ARF转录因子是植物中生长素响应的关键调节因子,杨树基因组中总计有35个Aux/IAA和39个ARF基因被识别[2]。Aux/IAA转录因子IAA9的功能涉及果实发育、叶片形态建成[3]和毛白杨P.tomentosa次生木质部发育[4]。沈韵[5]研究了毛白杨的IAA9在木质部、韧皮部、茎、根中的差异表达。目前未见到从欧洲黑杨P.nigra中克隆IAA9的报道。
为探讨杨树苗期高生长与内源IAA的关系,从分子角度研究高生长形态建成,在黑杨派树种高生长定位观测[1]和内源IAA及ABA含量分析[6]基础上,本文以欧洲黑杨为材料,采用RACE(Rapid Amplification of cDNA Ends)技术克隆了与高生长密切相关、与叶片发育有关的生长素基因家族的PnIAA9,并进行了杨树Aux/IAA基因家族的进化分析。
1 材料与方法
1.1 材 料
1.1.1 试验材料
4月上旬,采用硬枝扦插技术,在花盆(直径33 cm,高26 cm,底径20 cm)内扦插1段18 cm长欧洲黑杨1年生茎段。花盆中土壤为沙壤土与腐殖土混合物(体积比为3∶1)。温室温度控制范围15~25 ℃,苗木常规水肥管理。7月中旬,剪取欧洲黑杨茎尖并置于冻存管中,立即投入液氮,取出后-80 ℃保存。
1.1.2 主要试剂
RNA 提取使用Trizol试剂(Invitrogen,USA),ExTaq DNA Polymerase(TaKaRa,Japan),RQ-DNase(Promega,USA),RNase Inhibitor(TaKaRa,Japan),MM-LV反转录试剂盒(Promega,USA),dNTP(华绿源生物,中国),pGEM-T easy试剂盒(Promega,USA),PCR扩增产物回收和纯化试剂盒、质粒提取试剂盒(AxyGen,USA),SMARTTM RACE cDNA Amplification Kit(Clontech,USA),引物合成(北京奥科生物,中国)。
1.2 试验方法
1.2.1 Total RNA提取及检测
按照Trizol试剂盒说明书提取欧洲黑杨总 RNA,使用Agilent 2100 Bioanalyzer(Agilent RNA 6000 Nano Kit)检测总 RNA的浓度、RIN值、28S/18S和片断大小。样本的纯度使用紫外分光光度计NanoDropTM进行测定。
1.2.2 基因的克隆
RACE引物设计:根据PopulusDB EST数据库中IAA9序列设计RACE引物如下:3IAA9:5′-CCCTTTGCATCCCTCAAATGATGGTC-3′;5IAA9:5′-GCGGAGCTGTTACGAAGTGGTCTGGT-3′。其中,3IAA9从基因的3′ 端克隆;5IAA9从基因的5′ 端克隆。
欧洲黑杨IAA9基因的克隆采用SMART RACE技术,详细过程参照Clontech生物公司生产的SMARTTM RACE cDNA Amplification Kit说明书。
反转录:分别向标记5′和3′ 的200 μL PCR反应管内加2 μL RNA;向标记3′ 的PCR反应管内加1 μL 3′RACE CDS,1 μL Extension Buffer,1 μL H2O混匀后72 ℃温浴2 min,立即冰浴冷却;向标记5′ PCR反应管内加1 μL 5′ RACE CDS,1 μL Extension Buffer,1 μL 5′ RACE SMART Oligo混匀后72 ℃温浴2 min,立即冰浴冷却;分别向上述PCR反应管内加1μL DTT,2μL 5×First strand buffer,1 μL dNTP和1 μL SuperScriptase II,混匀后42 ℃温浴60 min,48 ℃温浴30 min。反转录完毕,分别向各反应管内加100 μL灭菌水,混匀后-20 ℃保存。
3′ RACE/5′ RACE扩增:分别向标记5′和3′两个200 μL PCR反应管内加2.5μL相应的第一链合成产物;向5′ 标记的PCR反应管内分别加1μL ycc5IAA92(10 μmol·L-1)引物,向3′ 标记的PCR反应管内分别加1 μL ycc3IAA92(10 μmol·L-1)引物;向上述PCR反应管内分别加47.5 μL按照说明书配制的反应混合液,混匀后立即按照下列反应体系进行PCR反应:先96 ℃预变性1 min,然后 96 ℃ 10 s、72 ℃ 2 min进行10个循环,再96 ℃ 10 s、68 ℃ 30 s、72 ℃ 2 min 15个循环。反应完毕后,取5 μL在1%琼脂糖凝胶电泳检测扩增片段的特异性和大小。
扩增产物切胶纯化:使用AxyGen公司生产的琼脂糖凝胶及PCR产物纯化试剂盒,对PCR扩增产物切胶纯化。
T-载体连接、转化和测序:将凝胶纯化的目的片段按照操作说明与pGEM-T Easy载体连接,连接产物转化E.coliDH5α 感受态细胞。在含有IPTG(异丙基硫代半乳糖苷)和X-gal(β-半乳糖苷酶)的LB/Amp平板筛选阳性植株,提出质粒并测序。测序引物是M13F和M13R,测序结果拼接后得基因的全长序列。
1.2.3 基因IAA9序列分析及进化树构建
应用DNAMAN软件对克隆的PnIAA9进行序列分析,应用美国国立生物技术信息中心(NCBI)Conserved Domains分析PnIAA9结构域,应用Culusterx软件进行氨基酸多序列比对。在文献分析基础上,从NCBI找到杨树中Aux/IAA家族基因并下载序列,应用MEGA7软件,通过NJ(Neighbor-joining)法构建系统进化树。
2 结果与分析
2.1 欧洲黑杨IAA9基因克隆
提取的欧洲黑杨茎尖的总RNA质量参数为OD260∶OD280=1.98,OD230∶OD260=2.01。总RNA 28S/18S条带亮度比值大于2,表明提取的总RNA经RNase-free DNase处理后完整性较好,没有降解和杂质污染现象。
图1为IAA9基因的5′ RACE和3′ RACE扩增图谱,从图1可以看出,IAA9 5′ RACE 克隆特异性比较好,为单一条带扩增的产物,大小为900 bp左右。而3′ RACE克隆特异性比较差,被扩增的产物大小为700 bp左右的特异条带,此外在1.0 Kb有一条非特异扩增条带。通过切胶回收5′ RACE产物和3′ RACE产物700 bp左右的条带,连接T载体,转化大肠杆菌感受态细胞,经测序分析,获得IAA9基因5′ 和3′ RACE序列。去掉重复序列,经拼接得到IAA9全长,命名为PnIAA9,长度为1 786 bp。全长DNA序列和氨基酸序列见图2。
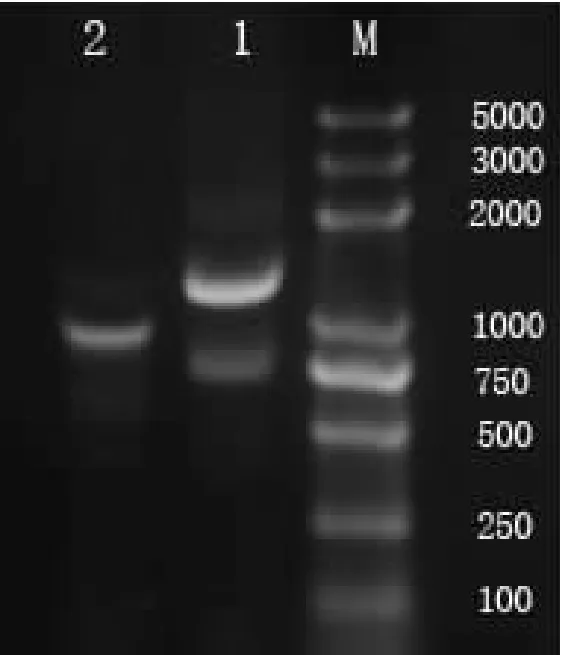
注:M为DL2000 Plus Marker;1为3′ RACE;2为5′ RACE。

图2 PnIAA9全长DNA序列和氨基酸序列
把PnIAA9 full length数据在NCBI中做Blast,得到的100个结果中,前14个结果的Per.Ident大于90%,其中第一位的P.deltoides值达到99.73%;排第2位的auxin-responsive proteinIAA9[P.trichocarpa]Per.Ident达到99.18%;排前11位的都是杨树的IAA9。
2.2 PnIAA9序列分析
利用DNAMAN软件进行分析,该基因编码一条由365个氨基酸组成的多肽链(图3)。

图3 PnIAA9全长序列的开放阅读框
PnIAA9结构域分析结果表明,其属于Aux/IAA家族。氨基酸序列特异点区间212~357,E-value= 6.19e-110,E-value数值小说明结果很有意义。Aux/IAA基因的转录是由植物激素生长素快速诱导的,该家族的一些成员较长,并包含一个N末端DNA结合域。
选择PnIAA9和从NCBI下载的毛果杨IAA9、IAA20、IAA32、IAA33氨基酸序列,进行氨基酸多序列比对,结果见图4。典型的Aux/IAA蛋白具有4个保守结构域,分别称为Domain I、II、III和IV[7]。Domain I、II位于氨基端(N端),而Domain III和IV位于羧基端(C端)。Domain II是Aux/IAA蛋白被泛素化降解的靶位点,其核心序列VGWPP是经典蛋白降解序列。PnIAA9、IAA9和IAA20都具有VGWPP序列(图4中黑色方框),说明PnIAA9等3个蛋白易受到生长素的降解,是短命蛋白。
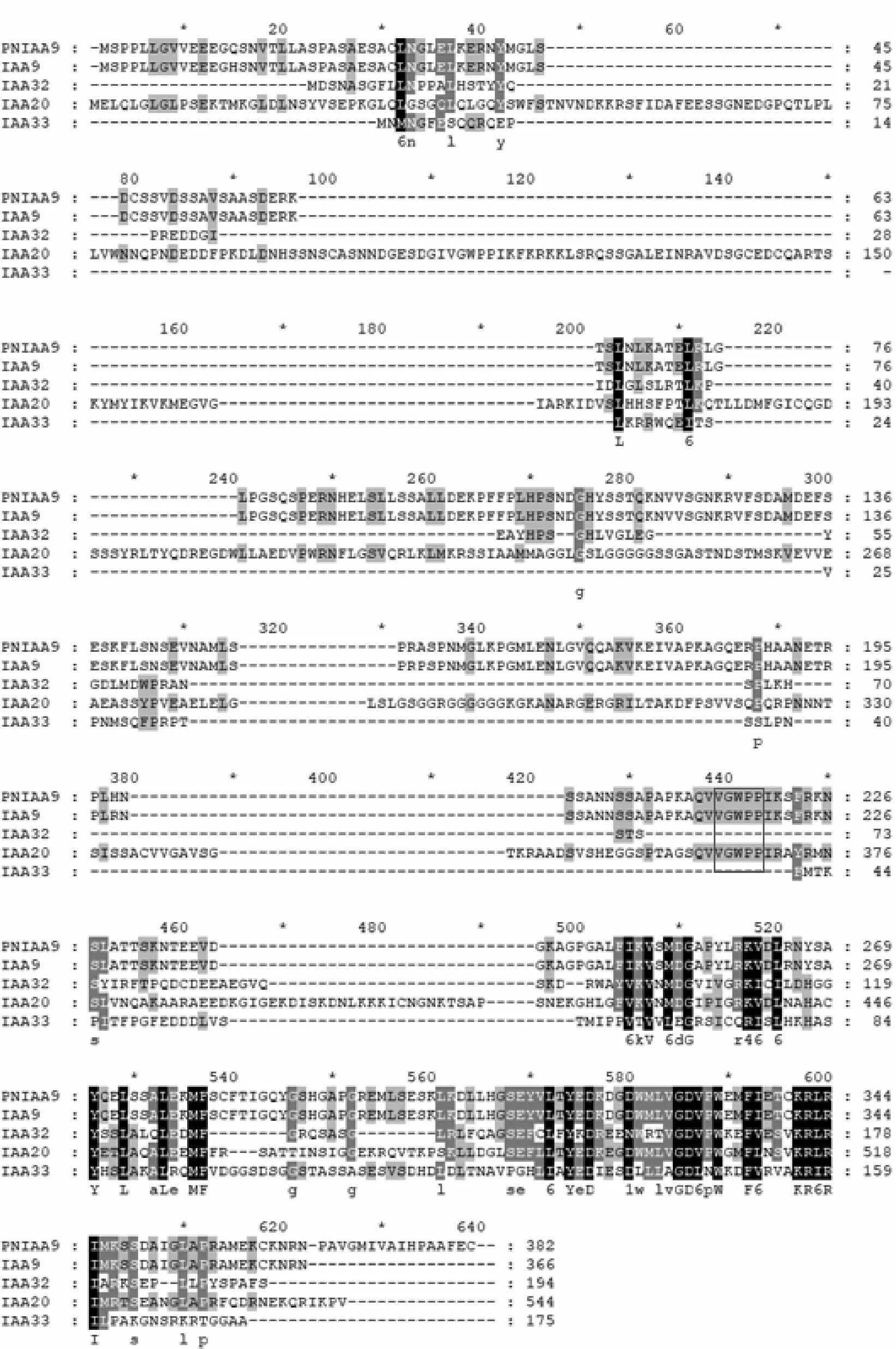
图4 PnIAA9等氨基酸多序列比对结果
2.3 PnIAA9 与杨树Aux/IAA进化树构建
综合文献分析结果,在NCBI上找到并下载18个杨树Aux/IAA生长素响应蛋白序列,多数属于毛果杨P.trichocarpa基因组,与从欧洲黑杨中克隆的PnIAA9构建进化树,结果见图5。
由图5可知,PnIAA9与毛果杨的IAA9同源性最高,与毛果杨的IAA33基因序列相似度最低,进化关系最远。

图5 PnIAA9与18个杨树Aux/IAA进化树构建
3 结论与讨论
杨树苗高由节间长和节间数构成,由于其叶互生,节间数量与叶片的数量密切相关。在枝条长度相等时,美洲黑杨节间相对较长但节间数量少,欧洲黑杨节间相对较短但数量较多。通过克隆欧洲黑杨节间数量基因,再通过转基因技术转化到美洲黑杨,可能培育出节间长且节间数量多的速生树种。
一般认为,IAA和叶的起始有关[8],阻断生长素的运输会阻止叶的起始,在拟南芥的生长素运输缺陷突变体中可以观察到这个现象[9]。高红兵等[10]应用GC-MS-SIM技术测定了毛白杨休眠芽中IAA和ABA含量,杨成超等[6]研究了IAA与ABA含量的比值与3个黑杨派杨树苗高的关系,结果表明苗高月增量与顶芽的IAA/ABA呈显著正相关。目前,在拟南芥中已经有通过Aux/IAA9调控来调节拟南芥生长的研究[11]。
本研究从欧洲黑杨茎尖中克隆的PnIAA9基因长度为1 786 bp,编码365个氨基酸,并进行杨树Aux/IAA家族的进化分析。将来可以利用基因编辑等技术提高或降低顶端分生组织IAA的含量来调控内源IAA/ABA含量的比值,也可以通过对IAA的调控来控制节间数量的多少实现苗高性状的分子调控和遗传改良,从而为培育矮化园艺植物和超级速生杨树打下基础。
