2株肺炎克雷伯菌肌尾噬菌体的鉴定
2021-10-25高云航张红伟马红霞么乃全徐凤宇
高云航,张红伟,马红霞,么乃全,徐凤宇
(1.吉林农业大学动物科学技术学院,吉林 长春 130118;2.动物生产及产品质量安全教育部重点实验室,吉林 长春 130118)
肺炎克雷伯菌(Klebsiellapneumoniae)是人和动物的共生菌,部分菌株致肺炎、败血症、子宫内膜炎等疾病;高毒力肺炎克雷伯菌(HypervirulentK.pneumoniae,hvKp)除引起社区获得性肺炎(Community-acquired pneumonia,CAP)外[1],也引起伴有转移脓毒性病变和其他化脓性脓肿和肝脓肿[2],其所致医院获得性肺炎(Hospital-acquired pneumonia,HAP)的28 d病死率达39.6%[1]。荚膜多糖是肺炎克雷伯菌的重要毒力因子,K1、K2、K5、K20、K54和K57等荚膜型与社区化脓性感染相关[3]。随着抗生素使用量的增长,肺炎克雷伯菌呈现明显耐药现象[4],加剧了控制该菌感染的难度。为缓解细菌耐药性蔓延,开发补充或替代抗生素的抗菌剂用于防治细菌感染已成当务之急。
毒性噬菌体除复制后具有特异杀菌作用外[5],其产生的解聚酶可清除荚膜、生物被膜、减弱细菌的毒力[6-7]。Wang等表达并纯化了vB_KpnP_IME321株噬菌体的荚膜解聚酶Dp4,用其预处理实验小鼠,在致死量宿主菌Kp409株K.pneumoniae感染后,实验小鼠全部存活,感染宿主菌后口服该酶,实验小鼠存活率显著提高[6]。噬菌体或脂质体携带的鸡尾酒噬菌体可保护所有肺炎克雷伯菌感染的BALB/c小鼠[8]。更可喜的是,肺炎克雷伯菌K7株噬菌体GH-K3的抗性突变株K7RR毒力远低于K7株[9]。可见噬菌体及其相关有效成分非常有希望成为潜在的新型抗菌剂。本研究以分离自猪子宫内膜炎的肺炎克雷伯菌为宿主,分离毒性噬菌体,在鉴定基础上检测了其主要生物学特性,为更深入研究和应用肺炎克雷伯菌噬菌体提供资料。
1 材料与方法
1.1 菌株 猪源肺炎克雷伯菌KL1株、KL2株、KL3株、KL4株、KL5株、KL6株、KM1株和KM2株分离自肺炎患猪,414株、KE1株、KE2株、KE3株、KE4株和KE5株分离自子宫内膜炎患猪,共14株,均由本实验室鉴定并保存。
1.2 主要试剂与仪器 上层LB半固体培养基、下层LB固体培养基、2×LB培养液、SM液等依标准方法配制。CP100MX超速离心机为日本日立公司产品;TEM-100CX透射电镜为日本TEOLLTD公司产品。
1.3 方法
1.3.1 宿主菌筛选 复苏分离自子宫内膜炎患猪的6株肺炎克雷伯菌,挑单菌落于LB液体培养基中培养过夜,分别取150 μL涂于LB平板,静置后将头孢噻肟等12种药敏纸片贴于平板上,大肠杆菌ATCC25922株作质控菌株,培养16 h后观察,根据动物源及人医临床实验室标准化研究所(CLSI)细菌药敏试验标准[10-11]评估。
1.3.2 噬菌体分离、纯化 以筛选的多重耐药肺炎克雷伯菌为宿主,将采自长春市某猪场、牛场健康猪、牛的新鲜粪便的新用生理盐水悬浮,静置后取上清8 000 r/min离心20 min,取0.22 μm滤器过滤后滤液10 mL加10 mL 2×LB液体培养基中,加入新培养的宿主菌、CaCl2溶液(终浓度1 mmol/L),100 r/min培养8 h。
噬菌体富集:离心培养液取上清过滤,如前述方法富集4次后再离心,过滤得噬菌体原液。
点滴法验证原液:将80 μL新培养宿主菌涂于LB平板,静置20 min,滴5 μL噬菌体原液,培养12 h观察结果。
纯化噬菌体:取100 μL稀释噬菌体原液,与100 μL新培养的宿主菌液、CaCl2浓缩液混匀,室温静置15 min,加入5 mL 50 ℃上层LB培养基,混匀后速倾于下层LB平板上,37 ℃培养12 h。用灭菌牙签挑取相对大而透明的单斑于1 mL SM液中,4 ℃ 冰箱中静置12 h,离心、稀释再铺双层平板,重复多次,纯化噬菌体。
1.3.3 噬菌体裂解谱测定 以宿主菌外的13株肺炎克雷伯菌为受试菌,参考1.3.2中点滴法操作。
1.3.4 透射电镜观察 取稀释的纯化噬菌体,培养成噬菌斑网,加入3 mL SM液,4 ℃冰箱中过夜,吸出噬菌体,12 000 r/min离心20 min,取上清滴于铜网上,静置1 min,2%磷钨酸负染90 s,透射电镜观察。
1.3.5 氯仿、乙醚敏感性试验 (1)氯仿敏感性试验:将950 μL噬菌体与50 μL氯仿混匀,设对照组,4 ℃冰箱过夜,测效价。(2)乙醚敏感性试验:将800 μL噬菌体与200 μL乙醚混匀,设对照组,冰浴振荡2 h,3 000 r/min离心15 min,取水相测效价。
1.3.6 热及酸碱稳定性试验 设7个水浴温度(40~ 70 ℃,间隔5 ℃),分别放入盛1 mL噬菌体的EP管,作用20、40 min和60 min后分别取300 μL,冷却测效价。
分装900 μL pH 3~13的液体LB,37 ℃预热,分别加入100 μL噬菌体,混匀,37 ℃培养3 h,测效价。
1.3.7 最佳感染复数测定 将活菌计数的宿主菌与已知效价的噬菌体按表1稀释并等量混匀,于37 ℃摇床中培养5 h,4 ℃、12 000 r/min离心30 min,取上清过滤测感染复数(Multiplicity of infection,MOI)。
1.3.8 一步生长曲线绘制 按最佳MOI混匀噬菌体与宿主菌,37 ℃水浴中静置15 min,4 ℃、12 000 r/min 离心1 min,弃上清,用液体LB悬浮、洗涤沉淀2次,加37 ℃预热液体LB,37 ℃ 50 r/min培养,每10 min取500 μL,4 ℃、12 000 r/min离心20 min,取上清测效价,以效价的对数为纵坐标,以感染时间为横坐标绘生长曲线。
2 结果
2.1 宿主菌筛选 由药敏试验知肺炎克雷伯菌414株仅对头孢唑啉、头孢噻肟、新霉素和阿奇霉素敏感,对诺氟沙星、阿米卡星、庆大霉素、卡那霉素、四环素、多西环素等耐药,为6株试验菌中抗菌谱最宽者,选为宿主菌。
2.2 噬菌体分离纯化 经多次分离,从新鲜健康猪粪、牛粪中各得到1株噬菌体,分别经4次挑单斑纯化,得边缘整齐、透亮的均一噬菌斑(封底彩版图1),命名为KP1、KP2。

图1 噬菌体KP1、KP2纯化后噬菌斑Fig.1 The plaques of phage KP1,KP2 after purification
2.3 噬菌体裂解谱测定 检测表明,KP1不裂解13株受试菌,KP2可裂解KL6株,裂解率分别为7%、14%,说明2株噬菌体特异性较强。
2.4 噬菌体电镜观察 透射电镜观察到KP1、KP2均呈蝌蚪状复合对称,有正二十面体头部、可收缩尾,可见尾板,属肌尾噬菌体科;KP1头长宽(78±1.5)nm,尾长(109±2)nm;KP2头长宽(72.92±3.81)nm,尾长(96.68±5.22)nm(封底彩版图2)。
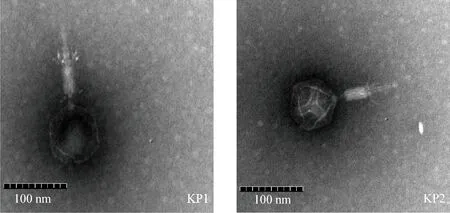
图2 噬菌体KP1、KP2电镜图(40.0 k×)Fig.2 Electron micrograph of phage KP1,KP2 (40.0 k×)
2.5 氯仿、乙醚敏感性试验 KP1、KP2经氯仿作用后,效价分别为6×108PFU/mL、1.06×109PFU/mL,经乙醚作用后效价分别为1.69×108PFU/mL、1.56×108PFU/mL,与对照组效价(KP1为9.2×108PFU/mL、KP2为5×108PFU/mL)差异不显著。说明噬菌体KP1、KP2对乙醚和氯仿不敏感,推测其无囊膜。
2.6 热及酸碱稳定性试验 热稳定性结果见封底彩版图3,KP1、KP2在40~60 ℃时,效价变化不明显,70 ℃作用20 min以上时,二者被灭活。
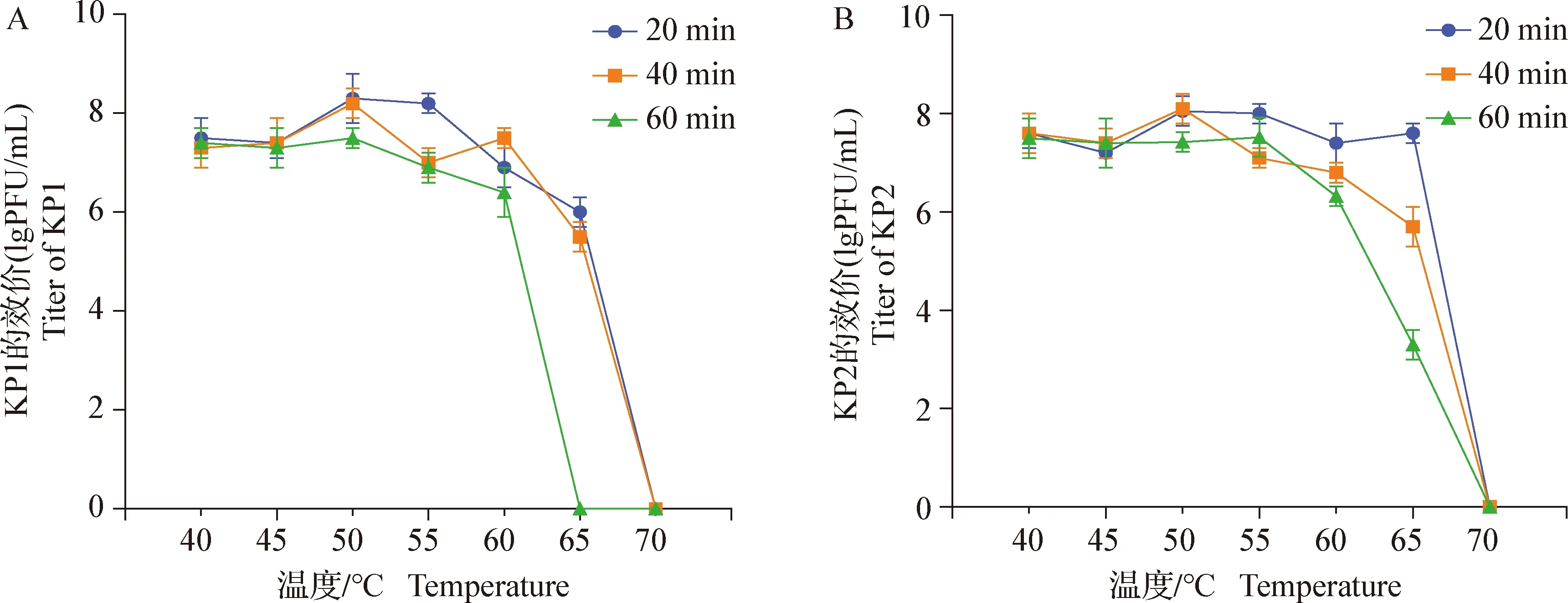
图3 噬菌体KP1、KP2的热稳定性Fig.3 Thermal stability of phage KP1,KP2A:KP1;B:KP2
KP1在pH 5~11条件下效价变化不明显,对pH 4和12的环境也有一定耐受性,在强酸或强碱环境中(pH≤3或≥13时)被灭活;KP2在pH 4~10的环境下效价基本稳定,对pH 3和11的环境有较好耐受性,在强酸或碱环境下(pH≤2或≥12时)被灭活(图4)。

图4 噬菌体KP1、KP2的pH稳定性Fig.4 pH stability of phage KP1,KP2A:KP1;B:KP2
2.7 噬菌体的最佳MOI测定 当MOI为0.1时,噬菌体KP1效价为1.6×109PFU/mL;MOI为0.01时,KP2效价为1.05×109PFU/mL,分别为7个MOI中最高值,为最佳MOI(表1)。
2.8 噬菌体的一步生长曲线 绘制噬菌体KP1、KP2的一步生长曲线(图5),发现其繁殖规律图像分别基本符合四次函数y=0.000 4x4-0.016x3+0.212 9x2-0.606 5x+4.770 8和三次函数y=-0.003 8x3+0.100 1x2-0.400 8x+4.731 1。KP1、KP2的潜伏期分别为20 min、30 min,爆发期分别约60 min、80 min,爆发量分别约为15 PFU/cell、11 PFU/cell。
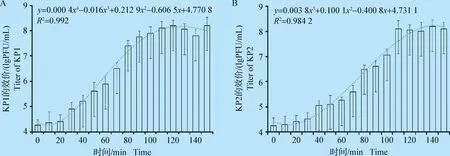
图5 噬菌体KP1、KP2的一步生长曲线Fig.5 One-step growth curve of phage KP1,KP2A:KP1;B:KP2
3 讨论
近年来,随着对肺炎克雷伯菌耐药现象的关注,能裂解该菌的噬菌体也被广泛关注和研究。Gao等[12]从北京某医院废水中分离到KN2荚膜型肺炎克雷伯菌的毒性、短尾噬菌体vB_KpnP_IME337,潜伏期为10 min,基因组为44 266 bp;Morozova等[13]报道了多重耐药肺炎克雷伯菌短尾噬菌体KP8基因组为73 679 bp;Tan等[14]从上海某医院分离了ST11型产碳青霉烯酶肺炎克雷伯菌(Carbapenem-resistantKlebsiellapneumoniae,CRKP)的短尾噬菌体117和31;Shi等[15]分离CRKP的毒性、短尾噬菌体kpssk3,潜伏期也为10 min,可裂解受试27株CRKP临床分离株,在温度和pH改变时稳定性较好,基因组为40 539 bp;短尾噬菌体vB_KpnP_KL106-ULIP47 和vB_KpnP_KL106-ULIP54由Thiry等[16]分离;Cai等[17]从医院污水分离的长尾噬菌体vB_KpnS_GH-K3(简称GH-K3)的爆发量为291 PFU/cell,在pH 6~10和50 ℃以下稳定,基因组为49 427 bp;长尾噬菌体TSK1有狭窄宿主谱,在pH 7和37 ℃下稳定,基因组为49 836 bp[18];ZCKP1是从埃及Giza市的淡水中分离的肌尾噬菌体,基因组为150.9 kb[19];噬菌体vB_Kpn_F48也属于肌尾噬菌体科,对温度和pH变化高度稳定,基因组为171 kb[20]。本试验从健康猪、牛粪中各分离到1株 毒性、无囊膜肌尾噬菌体,裂解谱较窄,在40~60 ℃、pH 4~11的条件下稳定性较好,与kpssk3、TSK1、vB_Kpn_F48、GH-K3稳定性相近,KP1、KP2最佳MOI分别为0.1、0.01,潜伏期分别为20 min、30 min,爆发期分别约60 min、80 min,较前述报道的噬菌体潜伏期略长,爆发量约分别为15 PFU/cell、11 PFU/cell,约为GH-K3的1/20、1/30。
对肺炎克雷伯菌噬菌体功能研究也有相关报道,Domingo等[21]分析了所分离的4株感染K22荚膜型的肺炎克雷伯菌短尾噬菌体序列,表明其有假定的编码解聚酶基因;ZCKP1在25 ℃时可使宿主菌K.pneumoniaeKP/01的浓度降低超过2log10CFU/mL,50 PFU/cell的ZCKP1可使宿主菌生物被膜量降低50%[19];TSK1在感染之初的14 h也可减慢肺炎克雷伯菌繁殖,用TSK1处理后可使培养的肺炎克雷伯菌生物被膜量减少85%~100%,而用TSK1前处理在培养的最初24 h使宿主菌减少比例大于99%[18];117株噬菌体治疗Kp36所致尿路感染模型初始也表现出很强裂解能力[14];Thiry等[16]以梅隆线虫幼虫为模型,发现用噬菌体vB_KpnP_KL106-ULIP47 和vB_KpnP_KL106-ULIP54治疗肺炎克雷伯菌ST258株感染,用vB_KpnP_K1-ULIP33治疗ST23株感染,可使死亡率从70%降至20%。kpssk3、vB_Kpn_F48无抗生素抗性基因、毒力因子基因及整合酶基因[15,20]。本试验结果虽为噬菌体用于防控耐药性肺炎克雷伯菌感染提供了资料,但2株噬菌体是否携带抗生素抗性、毒力、整合酶基因及对可裂解菌所致疾病的防治效果均有待深入探究。
