光对高等真菌次级代谢影响的研究进展
2021-10-25张宗源潘梦诗郭文阳岳丹丹张英涛亓兰达
张宗源,潘梦诗,郭文阳,岳丹丹,张英涛,亓兰达
(河南省科学院 生物研究所,郑州 450008)
真菌物种丰富,被认为是陆地和水生生物中最大的生物多样性资源之一。真菌在长期演化过程中形成了可以产生多类型、多功能的次级代谢物代谢机制,这些次级代谢产物有利于真菌争夺营养物质,可抵御恶劣的生存环境,实现自我保护[1],完成细胞增殖、分化和整个生命周期,并可作为与环境中生物交流的信号分子[2]。不仅如此,丰富的真菌次级代谢物结构新颖,骨架丰富,活性独特,是现代新药研发的重要来源,在疾病治疗中发挥巨大作用[3]。目前在临床上有重要应用价值的抗生素、免疫调节剂、降血脂、胆固醇等药物多数源于真菌的次级代谢物[4]。由于真菌次级代谢物具有重要应用价值,所以次级代谢研究一直是研究热点。
环境因素对真菌次级代谢生物合成起着重要的作用。光是最重要的环境因素之一,它对生物体的生理和行为产生决定性影响,真菌也不例外。在长期进化过程中,真菌形成精细且复杂的光感应系统,它们通过光敏蛋白和特异的信号网络传导途径将感受到的光信号(有光或无光、光的方向、光质、光强等)转变为化学和生物信号,并做出反应以更好地适应环境[5]。本文对近年真菌光生物学研究进行概述,介绍光对真菌天然活性次级代谢物合成的影响及其分子调控机制的研究。
1 真菌的光生物学研究
光以一种被称为光子的粒子形式存在,具有波粒二象性,根据不同的波长将其分为紫外辐射区(<380 nm)、可见光(380~780 nm)、红外辐射区(>780 nm)[6]。光为真菌提供重要的环境信息,真菌的光反应接近于人类视觉,即从蓝/近紫外区(450 nm)到近红外区(700 nm)。在红曲霉(Monascus)光生物学研究中,黑暗条件促进其营养生长,且分生孢子数量以及闭囊壳的数量明显大于光照条件[7]。光质对真菌也有不同程度的影响,蓝光和(或)红光会抑制构巢曲霉(Aspergillusnidulans)[8]、烟曲霉(Aspergillusfumigatus)[9]、禾柄锈菌(Pucciniagraminis)[10]等真菌孢子的萌发,除了蓝光和红光之外,最近的研究表明,绿光可以抑制灰葡萄孢(Botrytiscinerea)的孢子萌发,并在该反应中发现了一个功能未知的视蛋白[11]。在灵芝菌丝体培养过程中,蓝光促进灵芝菌丝体生长,绿光生长的最慢,同时不同光质对灵芝菌丝体生长形态也有显著影响,蓝光处理菌丝体生长密实,红光和黄光处理菌丝体生长蓬松,绿光处理菌丝体生长则为前期较稀薄,后期密实[12]。
光在大型真菌由营养生长阶段转变为生殖生长阶段的发育过程中起着至关重要的作用。在人工栽培食药用菌中,光照对香菇菌棒的转色起到重要作用,相比黑暗环境,光照条件下香菇菌丝体转色正常[13]。光作为环境因子之一,对食药用菌生长具有调控作用,钱磊等[14]的文章中详细阐述了光质、光强、光周期对菌丝体生长、原基形成和子实体形态的影响。
2 光对真菌次级代谢产物生物合成的影响
真菌次级代谢物种类丰富,是天然药物的重要来源之一,丝状真菌例如半知菌类曲霉科青霉属(Penicillium)、曲霉属(Aspergillus)和头孢菌属(Cephalosporium)等菌株产生的次级代谢物广泛应用到医药生产中。大型真菌如灵芝、香菇、虫草等被称为食药用真菌,也是重要的天然活性物质来源[15]。真菌天然活性产物按照其合成途径归类为萜类,如灵芝酸、β-胡萝卜素等;多糖,如灵芝胞外多糖等;非核糖体多肽,如青霉素等;聚酮类,如红曲素、美伐他汀、竹红菌甲素等;生物碱,如虫草素等(图1)。
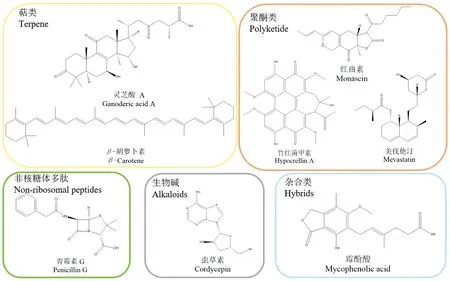
图1 真菌中具有代表性天然产物Figure 1 Representatives of fungal natural products
2.1 光对萜类化合物生物合成的影响
萜类化合物(萜烯类或者类异戊二烯)大多具有抗氧化、抗肿瘤、提高免疫力等功效。蛇孢菌素(ophiobolin A)是玉米小斑病菌(Bipolarismaydis)产生的具有抗肿瘤活性的二倍半萜类化合物,白光处理可以促进其生物合成,蓝光、绿光、红光、黄光则起到抑制作用[16]。类胡萝卜素是一类基本结构单位为异戊二烯的天然产物。蓝光能诱导蛹虫草菌丝体以及子实体类胡萝卜素的积累,失活蛹虫草中蓝光受体基因(Cmwc-1)导致类胡萝卜素产量降低[17]。通过控制光照和(或)黑暗能够获得四萜类化合物β-类胡萝卜素高产菌(MucorcircinelloidesMU218)[18]。蓝光对促进灵芝菌丝体三萜类化合物灵芝酸的积累优于其他光质,绿光和蓝光处理对提高灵芝酸种类有明显优势[12]。可见不同光质对同种萜类化合物有不同的影响,蓝光在诱导类胡萝卜素、虫草素和灵芝三萜类化合物具有促进作用。
2.2 光对多糖生物合成的影响
灵芝出菇实验发现不同光质影响不同时期灵芝多糖含量:现蕾期黄、蓝光有利于灵芝多糖含量积累;开伞期蓝光有利于灵芝多糖含量积累;弹孢后期绿光、蓝光、红光均能促进灵芝多糖积累[19],而红、蓝光照射后有利于液体发酵灵芝胞内多糖的积累[20]。液体发酵条件下,蓝光抑制胞内多糖积累[21],但是促进胞外多糖积累[22]。光不仅影响真菌多糖的积累,还影响多糖中单糖组分的含量。Puchkova 等[23]发现红、蓝光照射蛹虫草菌丝体后液体发酵,胞内多糖中葡萄糖组分百分含量增加,甘露糖组分和半乳糖组分含量降低;红、蓝光照射后,灵芝液体发酵中胞内多糖中甘露糖百分含量升高,葡萄糖含量降低[20]。由此可见,在不同生长时期,光对真菌多糖合成具有不同影响。此外,光质还影响真菌多糖单糖组分比例。
2.3 光对非核糖体多肽类化合物生物合成的影响
真菌可通过非核糖体途径合成一系列低分子量多肽类次级代谢产物,这种多肽类化合物统称为非核糖体肽(non-ribosomal peptides,NRPs),其通常具有抗菌、抗病毒、抗癌等生物活性。常见的非核糖体多肽有青霉素G、环孢菌素A和博来霉素。Spröte等[24]证明了光调控蛋白VeA的缺失会抑制青霉素的生物合成。Wang等通过构建△CsVosA(VeA组件)禾旋孢腔菌(Cochliobolussativus)突变株,发现两个非核糖体多肽合成酶(NRPs)基因表达下调[25]。
2.4 光对聚酮类化合物生物合成的影响
光质对聚酮类活性物质合成的影响与种属有关,光照强度对聚酮类化合物一定影响。Zhang等[26]研究表明红光、蓝光、绿光处理Haloroselliniasp.(No.1403)1403R和1403C产量增加76%。红曲色素是由Monascus产生的聚酮类色素,其生物合成与光照(蓝光)强度和时间有关[27-28]。发酵前期光处理会抑制橘青霉(Penicilliumcitrinum)产聚酮类化合物美伐他汀(Mevastatin)[29]。竹红菌素(hypocrellin)是竹黄菌(ShiraiabambusicolaP.Hennigs)主要有效成分,是一种苝醌类色素,属聚酮类化合物,红光可促进竹红菌素合成,蓝光则会抑制[30]。
2.5 光对生物碱类化合物生物合成的影响
蛇足石杉中的石杉碱,是吡啶酮类生物碱,研究表明黑暗处理比光照更有利于蛇足石杉内生菌石衫碱的积累[31]。虫草素属嘌呤类生物碱,具有抑制微生物、抗肿瘤、免疫调节等功效,液体发酵蛹虫草,蓝光处理蛹虫草产量最高[32],通过失活蛹虫草中蓝光受体基因(Cmwc-1),虫草素产量降低[17]。
2.6 光对其他活性物质生物合成的影响
二十碳五烯酸(eicosapentaenoic acid,EPA)是具有促进脑部发育和抗动脉粥样化功效的不饱和脂肪酸。粉核油球藻(Pinguiococcuspyrenoidosus)在光照强度为100 μmol /(m2·s)时产生的EPA比例最高[33]。霉酚酸(mycophenolic acid,MPA)是从短密青霉(Penicilliumbrevicompactum)发酵液中分离的一种具有多种活性的酶抑制剂。光照强度以及光质对其合成有不同影响,光强为1 400 Lx时MPA浓度最高,红光对MPA的合成有促进作用,蓝光产生抑制[34]。在A.nidulans中,白光可以促进亚铁血红素的合成,可能跟VeA互作蛋白(VipA)有关[35]。
3 光调控真菌次级代谢分子机制研究
在光响应研究中首要任务是要了解真菌如何感受到光。真菌感知光的能力与感光蛋白有关,不同的光受体蛋白能感应特定波段的光质。光受体蛋白的种类、结构和功能已经有了较为全面的研究[5],光调控真菌次级代谢及其分子机制研究也越来越深入(表1)。真菌次级代谢的分子调控是在转录、转录后、翻译以及表观遗传等多个层次上,有大量蛋白参与的复杂过程,其中不乏光作为重要的环境因子通过影响表观遗传因子(DNA甲基化酶Dnmt,组蛋白乙酰化酶HDA等)、全局调控因子(LaeA,VeA,VelB等)和途径特异性调控因子调控真菌次级代谢(图2)。

表1 几种主要活性物质及光调控分子机制Table1 Several main bioactive compounds and molecular mechanisms of light regulation

图2 光调控真菌次级代谢的分子机制Figure 2 Molecular mechanism of light regulation of fungal secondary metabolism
表观遗传对真菌次级代谢物合成具有调控作用,其主要研究内容为DNA甲基化和组蛋白乙酰化。DNA甲基化是指由S-腺苷甲硫氨酸提供甲基基团,在DNA甲基转移酶催化作用下,将甲基基团转移到DNA分子的碱基中,在真菌中,DNA甲基化发生在胞嘧啶上,而形成5-甲基胞嘧啶主要是在CpG二核苷酸上,DNA甲基化通过改变DNA稳定性、DNA构象、染色质结构等控制基因的表达,其中甲基化抑制基因表达,去甲基化促进基因的活化和表达。光照可诱导生物体基因组的甲基化和去甲基化[38];肖冬来等[39]研究表明光照可诱导广叶绣球菌(Sparassislatifolia)菌丝去甲基化,表明在光照条件下其更多基因表达。组蛋白乙酰化是组蛋白的乙酰化水平,是通过组蛋白乙酰化酶(HAT)和组蛋白去乙酰化酶(HDAC)来调控,根据组蛋白乙酰化与去乙酰化相互转变,就犹如基因的开关,可以控制基因的表达与沉默。组蛋白乙酰化可以影响真菌次级代谢物的生物合成。Osorio-concepción等[40]的研究中发现深绿木霉(Trichodermaatroviride)的组蛋白去乙酰化酶HDA-2是由光诱导的。以上结果均表明光可通过表观遗传调控真菌次级代谢。
全局性调控是指由全局性调控因子参与并对多个次级代谢物生物合成以及形态分化进行的调控。全局调控因子在真菌次级代谢调控中起着关键作用。其主要研究内容为Velvet蛋白和LaeA全局调控因子,而这两种蛋白都直接或间接受到光的调节。Velvet家族主要包含有VeA、VelB、VosA和VelC,其中VeA蛋白是光依赖性的无性/有性发育和次级代谢的全局调控因子,研究表明它与VelB、LaeA(loss of aflR expression A)形成异源三聚体系统调控构巢曲霉的生长发育和次级代谢[41]。光可以影响VeA蛋白在细胞质和细胞核的分布,从而影响其功能的发挥。在真菌体内,VeA为桥梁与其家族成员或LaeA及其家族成员形成复合物,直接或间接调控真菌有性、无性发育和次级代谢物的合成[42]。同时VeA蛋白还能够与红光受体蛋白相互作用,在构巢曲霉中其与红光受体蛋白FphA的组氨酸激酶结构域相互作用,且与LreA/LreB二聚体形成VeA-FphA-LreA-LreB光复合物[43],但目前还不清楚该复合物是如何调控真菌光依赖性的生长发育和次级代谢。
相对于全局性调控,光对真菌次级代谢还存在途径特异性调控,目前该方面研究较少。以光诱导类胡萝卜素合成为例,crgA基因是在研究丝状真菌光诱导类胡萝卜素生物合成过程中发现的一个关键负调控因子,其对类胡萝卜素结构基因及相关合成基因发挥着重要的调控作用。该基因受光的调控,其表达产物具有泛素连接酶活性,通过催化类胡萝卜素合成过程中其他蛋白泛素化来调控类胡萝卜素的合成[44]。由此推测在光的影响下,途径特异性转录因子基因表达受到调控,进而调控相关次级代谢基因簇的表达。
4 总结与展望
真菌因能生产有价值的天然化合物而受到高度重视,其次级代谢产物结构新颖,骨架丰富,活性独特,通常具有抗菌、抗肿瘤和抗氧化等特性,是现代新药研发的重要来源。然而这类化合物产率很低,不足以大规模提取与纯化。为了解决这一限制,通常通过发酵培养基优化、基因水平修饰和外源刺激等方法策略来提高该类化合物的生物合成。而光作为一种重要的环境信号,显然是一种有效、环保、清洁的能源和低成本的方法来调节真菌次级代谢物的生产。目前以LED为理想的光环境调控设施已经应用到食药用菌生产[45-46]。今后可以在工农业生产中加强LED的应用以提高药源真菌的活性成分。
随着光生物学研究的深入,越来越多的真菌光受体蛋白被发现,从光信号向生物信号转化出现了意想不到的复杂性。以目前的研究还不足以呈现出完整的真菌光响应信号通路图。虽然大多真菌有类似的光响应现象,但在不同的真菌种属间其调控机制存在差异。目前研究较为深入的为模式真菌N.crassa中的WCC蛋白复合体及其如何感知并传递光信号。然而为什么真菌可以感知不同的光色?推测其原因跟大自然环境竞争有很大关系。如在森林冠层环境中,绿光占主要优势[47],同样的已有研究表明红光比蓝光更容易穿透土壤[48]。同时,真菌对不同的光照强度做出不同反应,然而其影响机理需进一步研究。
真菌光响应被广泛研究,包括光生物学相关的转录因子和差异基因表达研究等方面也取得新进展。真菌次级代谢调控研究主要包括表观遗传调控、全局性调控、途径特异性调控。光在真菌发育和次级代谢过程中起着至关重要的作用,但其作用机制还不清楚。在模式真菌构巢曲霉中发现的光依赖全局调控因子Vet为我们提供了研究思路,光照抑制VeA-VelB复合蛋白进入细胞核,进而影响真菌的发育和次级代谢。而随后VelB-VeA-LaeA三聚复合物的发现更是将光信号调控真菌发育和次级代谢物生物合成与染色质的表观遗传修饰建立了桥梁。光照可以影响真菌DNA甲基化水平,推测可能跟具有甲基化功能的LaeA有关。真菌生物合成途径特异性转录因子主要包含Zn2Cys6和Cys2His2两种类型,研究表明60%的真菌次级代谢产物合成基因簇内含有途径特异性转录因子基因[49],真菌光响应与这两类调控因子的关系仍需进一步研究。
