温度和空间对大草蛉捕食梨瘿蚊的影响
2021-10-15凤舞剑胡长效朱强
凤舞剑,胡长效,朱强
(1.徐州生物工程职业技术学院/徐州市现代农业生物技术重点实验室,江苏 徐州 221006;2.江苏海洋大学食品科学与工程学院,江苏 连云港 222005)
梨瘿蚊[Dasumeirapyri(Bouch)]属双翅目瘿蚊科(Diptera:Cecidomyiidae),在欧洲、北美洲、大洋洲等地均有分布,国内已遍布江苏、辽宁等15个省,并已成为我国梨树生产上的主要害虫。梨瘿蚊以幼虫危害梨树新梢芽叶,导致叶片纵卷成苞,变硬、变脆、变褐,干枯,提早脱落,严重影响梨果产量[1,2]。梨瘿蚊幼虫在梨树上的空间分布属于聚集分布,且具有隐蔽性[2],国内尚无防治用药登记,利用天敌昆虫防治梨瘿蚊显得尤为重要。
目前,已报道的梨瘿蚊天敌主要有草蛉、花蝽、瓢虫、食蚜蝇、蜘蛛、瘿蚊广腹细蜂等[3,4],其中微小花蝽(Oriusminutes)对梨瘿蚊3~4龄幼虫有较好的捕食反应,1~5龄若虫及雌成虫最大捕食量分别达17.30、17.99、22.37、40.82、116.28、64.94头/d[1],但其饲养条件要求高。大草蛉(Chrysopapallens)属脉翅目草蛉科(Neuroptera:Chrysopidae),异名Chrysopaseptempunctata,东洋、古北区系共有种,国内除西藏未见采集记录外,其余各省均有分布[5,6]。大草蛉具有食谱广、繁殖能力强、易于规模化饲养等特点,对夹竹桃蚜(Aphisnerii)、烟粉虱(Bemisiatabaci)、草地贪夜蛾(Spodopterafrugiperda)、二斑叶螨(Tetranychusurticae)、豆大蓟马(Megalurothripsusitatus)、棉铃虫(Helicoverpaarmigera)等多种害虫均具有较强的控害潜能[6,7]。迄今为止,未见有大草蛉对梨瘿蚊捕食功能反应的研究报道。因此本试验利用HollingⅡ等模型研究温度和空间对大草蛉捕食梨瘿蚊的功能反应和搜寻效应影响,并对大草蛉防控梨瘿蚊的潜在能力进行评估。
1 材料与方法
1.1 供试虫源
大草蛉(Chrysopapallens)采自江苏精良农业科技有限公司徐州市丰县科技示范小麦田,室内用桃蚜饲养,繁殖2代以上,选择健康3龄幼虫和成虫,饥饿24 h后用于试验。梨瘿蚊[Dasumeira pyri(Bouch)]于2019年3月采自徐州市丰县谢大为家庭农场梨园,室内以盆栽无虫梨为寄主饲养2代以上,选择健康3龄幼虫用于试验。各供试昆虫均使用PRX-250A型人工气候箱饲养,相对湿度为(65±5)%,光周期L∶D=16 h∶8 h。
1.2 试验方法
1.2.1 不同温度下大草蛉对梨瘿蚊的捕食功能反应 试验于2019年7月进行,设19、21、23、25、27、29、31℃共7个温度梯度,2.6 L(22.5 cm×15.3 cm×12.0 cm)盒盖带透气阀的饲养盒底部铺湿润滤纸和5张梨树嫩叶。各试验条件下梨瘿蚊3龄幼虫密度分别设25、40、55、75、95、115头/盒,每处理接入1头大草蛉3龄幼虫或成虫,以不接入大草蛉3龄幼虫或成虫作对照。每处理重复5次,置入PRX-250A型人工气候箱中,24 h后记录剩余梨瘿蚊数量,计算大草蛉捕食量。
1.2.2 不同空间下大草蛉对梨瘿蚊的捕食功能反应 试验于2019年7月进行,设温度为(25±1)℃,4.6 L(26.5 cm×18.5cm×13.5 cm)和1.3 L(18.5 cm×12.3 cm×9.5 cm)两种饲养盒底部分别铺湿润滤纸和5张梨树嫩叶。各试验条件下梨瘿蚊3龄幼虫密度分别设25、40、55、75、95、115头/盒,每处理接入1头大草蛉3龄幼虫或成虫,以不接入大草蛉3龄幼虫或成虫作对照。每处理重复5次,置入PRX-250A型人工气候箱中,24 h后记录剩余梨瘿蚊数量,计算大草蛉捕食量。
1.3 数据分析
功能反应用HollingⅡ圆盘方程Na=a′TN/(1+a′ThN)拟合,搜寻效应用Holling(1959)方法S=a′/(1+a′ThN)拟合[8,9]。式中,Na为大草蛉3龄幼虫或成虫捕食量,N为梨瘿蚊3龄幼虫初始密度,a′为瞬时攻击率,T为搜寻总时间(本试验为1 d),Th为处理时间(即大草蛉幼虫或成虫捕食1头梨瘿蚊3龄幼虫所用时间),S为搜寻效应。
利用Microsoft Excel 2016和DPS 9.5进行数据处理。
2 结果与分析
2.1 温度对大草蛉捕食梨瘿蚊3龄幼虫的影响
2.1.1 温度对大草蛉捕食量的影响 在19、21、23、25、27、29、31℃共7个试验温度下,同一虫态大草蛉(3龄幼虫或成虫)在梨瘿蚊3龄幼虫同一密度下,19~25℃条件下捕食量随温度升高而升高,27~31℃条件下捕食量随温度的升高而降低。捕食量在25℃条件下最高,27℃次之,但两者差异不显著(表1)。

表1 不同虫态大草蛉在不同温度下对梨瘿蚊3龄幼虫的捕食量(头/d)
2.1.2 温度对大草蛉捕食功能反应的影响 HollingⅡ圆盘方程较好地描述了大草蛉3龄幼虫和成虫在不同温度下对梨瘿蚊3龄幼虫的捕食效应。在2.6 L饲养盒中,同一温度下大草蛉3龄幼虫对梨瘿蚊3龄幼虫的瞬时攻击率(a′)和捕食上限(1/Th)均高于成虫;同一虫态下大草蛉对梨瘿蚊3龄幼虫的瞬时攻击率和捕食上限最大值出现在25℃,其次为27℃,25℃和27℃大草蛉3龄幼虫对梨瘿蚊3龄幼虫的a′和1/Th分别为0.8105和0.7907、144.93头/d和140.85头/d,大草蛉成虫对梨瘿蚊3龄幼虫的a′和1/Th分别为0.3904和0.3902、43.86头/d和39.22头/d(表2)。捕食上限(1/Th)与温度(t)的关系为:1/Th(幼虫)=-1470.4760+128.5989t-2.5712t2(r=-0.9388*,*表示P<0.05水平上显著相关,下同),1/Th(成虫)=-444.0060+37.8733t-0.7455t2(r=-0.9388*)。
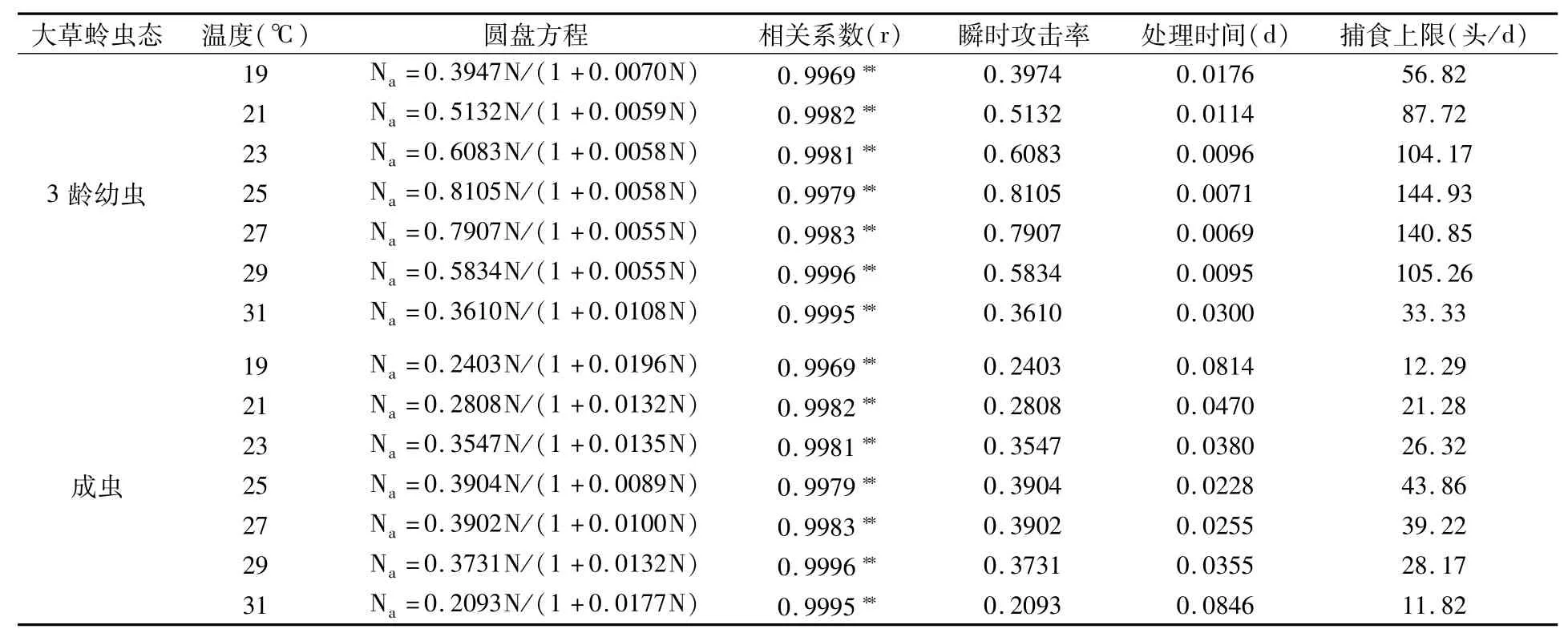
表2 不同虫态大草蛉在不同温度下对梨瘿蚊3龄幼虫的捕食功能反应
2.1.3 温度对大草蛉搜寻效应的影响 同一密度下,大草蛉3龄幼虫和成虫对梨瘿蚊3龄幼虫搜寻效应(S)与温度(t)均呈二次函数关系,高峰值出现在25℃和27℃(两者数值接近)(表3)。在温度、梨瘿蚊密度相同条件下,大草蛉成虫的搜寻效应均小于大草蛉3龄幼虫(表1)。

表3 不同虫态大草蛉对梨瘿蚊3龄幼虫搜寻效应与温度二次函数拟合模型
2.2 空间对大草蛉捕食梨瘿蚊3龄幼虫的影响
2.2.1 空间对大草蛉捕食量的影响 同一虫态大草蛉在梨瘿蚊3龄幼虫同一密度下,在1.3 L饲养盒中捕食量高于4.6 L,但在梨瘿蚊3龄幼虫25、40头/盒时,大草蛉成虫在1.3、4.6 L饲养盒捕食量差异不显著,在梨瘿蚊3龄幼虫25头/盒时,大草蛉3龄幼虫在1.3、4.6 L饲养盒捕食量差异不显著(表4)。

表4 不同虫态大草蛉在不同空间下对梨瘿蚊3龄幼虫的捕食量(头/d)
2.2.2 空间对大草蛉捕食功能反应的影响 大草蛉3龄幼虫和成虫在不同空间下对梨瘿蚊3龄幼虫的捕食功能反应模型均为HollingⅡ型方程。空间越小,大草蛉(成虫和3龄幼虫)对梨瘿蚊3龄幼虫的瞬时攻击率(a′)和捕食上限(1/Th)越大,处理时间(Th)越短。同一空间下大草蛉成虫a′、1/Th小于大草蛉3龄幼虫,而Th大于大草蛉3龄幼虫(表5)。

表5 不同虫态大草蛉在不同空间下对梨瘿蚊3龄幼虫的捕食功能反应
2.2.3 空间对大草蛉搜寻效应的影响 同一空间下同一虫态大草蛉搜寻效应随梨瘿蚊3龄幼虫密度的增大而减小,同一虫态大草蛉对同一密度梨瘿蚊3龄幼虫搜寻效应随试验空间的增大而减少,同一空间下大草蛉3龄幼虫对同一密度梨瘿蚊3龄幼虫的搜寻效应大于成虫(图1)。

图1 不同虫态大草蛉在不同空间下对梨瘿蚊3龄幼虫的搜寻效应
3 讨论与结论
在设定温度梯度下,大草蛉(3龄幼虫和成虫)对梨瘿蚊3龄幼虫的捕食功能反应均符合HollingⅡ型方程,在接近获得最大捕食量的密度时(115头/盒或95头/盒),捕食量出现反密度制约,这与大草蛉对二斑叶螨、草地贪夜蛾、桃蚜、夹竹桃蚜、烟粉虱、豆大蓟马、豆蚜等害虫的捕食功能反应研究结果一致[9-12]。
大草蛉捕食量受环境温度影响显著,张欣等[9]通过研究大草蛉对二班叶螨的捕食功能反应,发现大草蛉雌成虫对二斑叶螨的捕食效应在25℃时最高(a′、Na、1/Th、S最大)。本研究结果表明大草蛉3龄幼虫和成虫对梨瘿蚊3龄幼虫的捕食效应在25℃和27℃较高,可能与本试验设定的温度梯度、湿度与张欣等[9]设定的温度梯度(15、20、25、30、35℃)、湿度(60%)等不同有关。本试验2℃梯度差比张欣等的5℃梯度差更能准确地反映出大草蛉捕食效应随温度变化的差异。大草蛉对梨瘿蚊的最佳控制温度与梨瘿蚊的发生期相符,利用其防治梨瘿蚊具有较大的潜力。
同一温度同一空间下,大草蛉3龄幼虫对梨瘿蚊3龄幼虫的瞬时攻击率(a′)、捕食上限(1/Th)和搜寻效应(S)均高于大草蛉成虫,这与徐庆宣等[7]对大草蛉捕食草地贪夜蛾的研究结果一致。大草蛉3龄幼虫捕食量高于成虫,可能与食量大小和猎物(梨瘿蚊)的隐蔽习性有关。梨瘿蚊喜潜藏取食,大草蛉成虫翅的存在导致其搜寻能力降低。
空间越小,大草蛉(成虫和3龄幼虫)对梨瘿蚊3龄幼虫的瞬时攻击率(a′)、捕食上限(1/Th)和搜寻效应(S)越大,处理时间(Th)越短。这与胡长效等[8]研究龟纹瓢虫捕食褐软蚧的结果一致。本研究仅在室内比较了4.6 L和1.3 L两种规格饲养盒下对大草蛉捕食作用的影响,具体空间增或减至何种程度,大草蛉(成虫和3龄幼虫)对梨瘿蚊3龄幼虫捕食效能才不再变化,以及空间形状、空间内异质性等对捕食效能的影响尚有待进一步深入探讨。
