青冈栎林分空间结构对林下更新的影响
2021-10-14朱光玉王琢玙周根苗边更战张娅妮刘文剑
易 烜,朱光玉,王琢玙,周根苗,边更战,张娅妮,黄 靓,刘文剑
(1湖南省青羊湖国有林场,长沙410627;2中南林业科技大学林学院,长沙410004;3湖南省农林工业勘察设计研究总院,长沙410007;4湖南景辉农林生态科技有限公司,长沙410000)
0 引言
林分结构是指林分在没有受到毁坏的前提下,林分内部保留的一些较为稳定的结构规律[1],它可以分为空间结构和非空间结构2个方面。林分的空间结构是指林木的分布格局及其属性在空间上的排列方式,它是对森林内林分采伐、更新以及干扰活动状况的一种体现,在森林经营管理中是不能缺少的部分[2]。林下更新作为一种在森林中广泛存在的更新形式,在森林的更新及群落稳定方面有着十分重要的作用。林下更新不仅使物种丰富度和林分多样性更高,还使森林混交的程度加大;林下更新的幼苗生长令现存林分逐渐发育为复层、异龄、混交的近自然的状态[3]。研究林分空间结构对林下更新的影响,对天然次生林结构调整有实际意义。
青冈栎(Cyclobalanopsis glauca)在中亚热带常绿阔叶林中是比较常见的树种,在国内亚热带地区常被当作造林树种进行经营[4],是常绿阔叶林生态系统的重要组成部分,经济、社会及生态效益显著,科学研究价值较高。但由于受到环境和人为方面的干扰,现有的青冈栎林大多为次生林,天然更新速度减慢,如果次生林持续受到破坏,会朝着退化的方向发展。但假如可以按照其结构特点开展合理经营,就可使其在一定时间内得到快速恢复[5]。笔者以青羊湖国有林场13块固定样地的青冈栎林数据作为研究对象,通过Excel 2016软件分样地计算相关林分空间结构指标和林下更新指标,运用灰色关联度分析法分析对林下植被更新有显著影响的林分空间结构因子,根据分析结果对促进林下更新提出合理建议,旨在为青冈栎林的合理经营、充分发挥其生态服务功能提供理论参考。
1 研究区概况
研究区域位于湖南省青羊湖国有林场境内。林场位于宁乡市西部黄材镇(111°58'07″—112°05'18″E,28°08'02 "—28°14'20" N),属雪峰山余脉,由中低山向丘陵过渡的地带,海拔最高点927.4 m,最低点166 m。林场总面积1083.9 hm2,森林覆盖率96.99%,活立木蓄积115743 m3[6]。林场地处中亚热带向北亚热带过渡的大陆季风性湿润气候区,年平均气温16.2℃,最高40℃,最低气温为-11.5℃,年平均日照为1737.6 h,平均降雨量1438.8 mm,相对湿度85%,无霜期259~273天。
青羊湖林场属于中亚热带常绿阔叶林,据调查有维管植物189科651属1086种[7],现保存天然林684.2hm2[6],生长着青冈栎、钩栲、米槠林原生群落是迄今发现同类型森林中结构最为完好的森林[7]。林场现为湖南青羊湖生态保护与森林经营国家长期科研基地。
2 研究方法
2.1 样地设置与调查
采用典型取样的方法,在湖南省青羊湖国有林场的青冈栎林中随机选取13处20 m×30 m的样地(图1),每块样地内建立直角坐标系,对于胸径>5 cm的林木记录其树种、树高、胸径、坐标等因子,对于胸径≤5 cm的幼树记录其树种、地径、树高、冠幅。
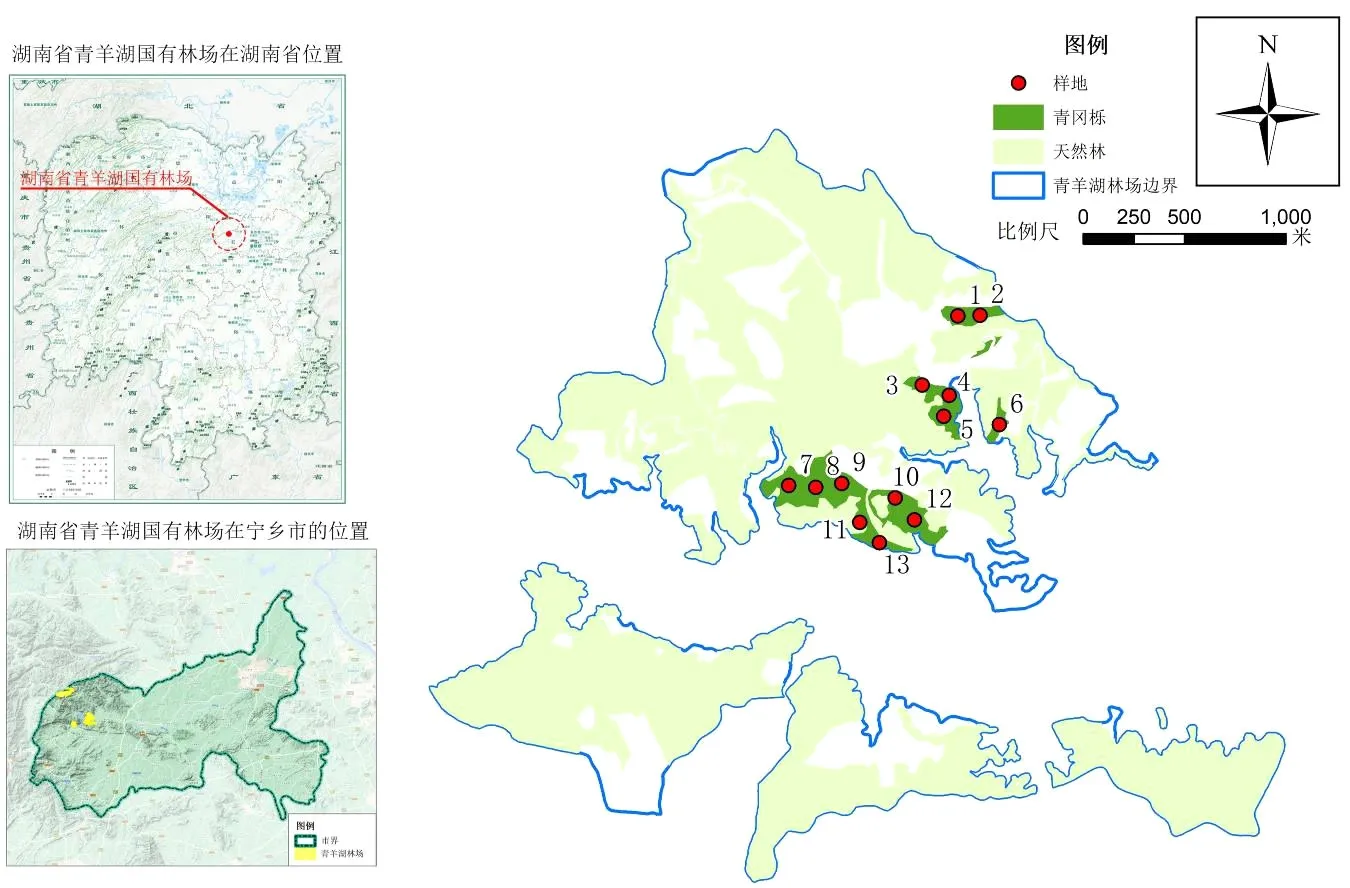
图1 青冈栎天然林样地分布图
2.2 林分空间结构研究
森林功能的充分发挥在相当大程度上是受到林分空间结构的影响,因为其影响着经营空间的大小及林分的稳定性。这些年,越来越多的学者对林分空间结构开展研究,相继提出空间结构参数体系,主要包括描述林分的空间分布格局、树种隔离程度、林木大小分化程度、开阔程度等结构参数。
2.2.1 林分空间分布格局的研究 林分空间分布格局可以分为随机分布、聚集分布和均匀分布,通过角尺度、聚集指数等能够对其进行分析。为描述结构单元内林木分布的均匀性,惠刚盈等[8-9]于1999年提出角尺度的概念,在此基础上又推出当n=4时,标准角为72°,而且通过计算机模拟林分所验证;聚集指数是由Clark和Evans[10]最早提出的最近邻体空间分析方法,能够对林分中最近相邻木距离上的空间分布格局进行描述。安慧君等[11]通过聚集指数对吉林省蛟河实验自然保护区的混交阔叶红松(Pinus koraiensis)林做出研究后表明,在计算聚集指数时,应结合数据的特点对边缘效应进行修正。
2.2.2 树种隔离程度的研究 混交度是用于表达林分中树种相互隔离程度的重要指数,由Gadow于1992年提出,在描述树种空间分布多样性上具有显著性意义。惠刚盈等[12]于2001年将混交度的概念引入国内,并对其进行了相关研究,结果表明,当使用4株邻近木计算混交度最为合理。汤孟平等[13]在对结构单元内树种差异和树种隔离关系进行总体分析后,提出全混交度指数,使不同混交结构中存在相同混交度的问题得到很好的解决。
2.2.3 林木大小分化程度的研究 大小分化程度是用来描述林分大小分化程度的一个空间参数,由Gadow等通过研究林木树种的直径和树高的空间分布提出的。后来,惠刚盈等[14]以Gadow的研究为基础,在1999年提出了大小比数的概念,它可以准确比较出参照树与其相邻木大小,能够描述林分在胸径、树高、冠幅等参数上的空间优势度。通过对杉木生态公益林不同龄组的大小比数等空间结构参数进行分析,曹小玉等[15]提出了杉木生态公益林经营管理的可靠方案。
2.2.4 开敞度的研究 开敞度是用来描述林下光条件状况的指标,它不仅反映光照强度,而且也反映了林木生长营养空间的大小。沈海龙等[16]曾研究红松林地上生物量和径高生长量受开敞度调控的影响。林存学[17]研究认为,次生林林冠下人工栽培红松的生长发育受开敞度的影响。
2.3 林下更新研究
在20世纪50年代,中国林业部综合队曾对兴安落叶松的更新条件进行过调查,表明以林分为基本单位,只反映了林分更新数量与总体状况之间的关系。国内这些年也开展了很多关于影响林下更新幼苗生长的深入研究,如徐化成等[18]综合探究了更新树种的生物特性、种子生产与扩散、幼苗的发生时期及幼苗的生长和存活率的季节变化。刘传照等[19]在东北对红松林幼树更新进行了研究。王贺新等[20]通过探究树种天然更新受枯枝落叶层的阻碍作用,发现了枯枝落叶层物理阻断对天然更新影响规律。徐庆祥等[21]对不同间伐强度下蒙古栎林下乔木幼树更新状况进行了研究,认为在采取中度间伐强度的情况下能够促进蒙古栎林下幼树的更新。毛磊[22]就樟子松林分空间结构对其植被更新影响做出研究,结果表明,通过对樟子松林分空间结构做一定调整,可以促进林下更新,改善竞争关系,这给研究林下更新提供了思路。
2.4 林分空间结构指数计算方法
通常采用大小比数、混交度、聚集指数和开敞度作为表达林分空间结构的参数[23]。为减少林分结构受边缘效应的干扰,设置5 m缓冲区。本研究空间结构单元数n取4。
大小比数U用来表达相邻木之间的大小关系,该指数被定义为大于参照树的相邻木个数占其空间结构单元内所有相邻木的比例。计算如式(1)。

式中,kij为离散型变量,当参照树i比第j株相邻木小时kij=1,反之kij=0。
聚集指数R用来表达林木空间分布格局,即最近邻单株聚集的平均值与随机分布下的期望平均距离之比,计算如式(2)。

式中,ri为第i株林木与其最近邻木之间的距离,N为样地中林木株数,F为样地面积。
混交度M用来表达相异树种空间隔离程度的指数,其被定义为在和参照树构成空间结构单元的相邻木中与参照树树种不同的林木占有的比例。

式中,在参照树和相邻木并非同一树种情况下Vij=1,相反Vij=0。
开敞度B被定义为参照木i到最近4株相邻上层林木的水平距离与该株上层林木高度的比值之和。

式中,Dij为参照树i到第j株相邻木的距离,Hij为第j株相邻木的高度。
2.5 林下更新指标计算方法
采用株数密度、树高、地径、冠幅作为表达林下更新的参数。单位面积株数是指每公顷幼树幼苗的株数,计算如式(5)。

式中,Ai为株数密度(株/hm2),Bi为各样地株数,Si为样地面积(m2),i为样地号。
2.6 幼树物种重要值分析法
重要值是一个用来衡量某个种在群落中的地位和作用的综合数量指标,以综合数值来表示植物物种在群落中的相对重要值,计算如式(6)。重要值取值范围0%~100%。
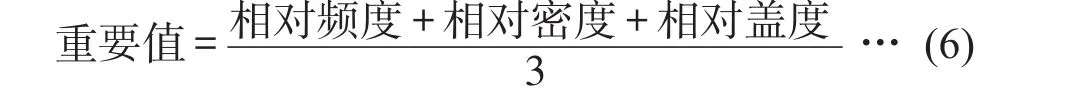
2.7 变异系数法
变异系数也称离散系数、差异系数,是衡量样本资料中各观测值变异程度的统计量。在比较1个或多个资料的变异程度时,常采用标准差与平均数的比值来比较,称为变异系数(CV),计算如式(7)。

变异系数具有直观简洁的优点,可以消除对2个或多个资料变异程度比较时平均数不同的影响。笔者将13块固定样地的林分空间结构和林下更新指数作为分析对象,通过平均数、标准差与变异系数3个统计指标对数据进行处理,分析不同样地的林分空间结构和林下更新指数变异性,变异程度越小,则代表该指数越稳定。
2.8 灰色关联度分析法
灰色关联度分析是源于灰色理论中的一种分析方法,它可以基于所研究的因素间变化的相似性来判断关联度。灰色关联度分析是按照各因素数列曲线形状的相近程度来对发展态势进行分析,简单来说就是指在系统发展过程中,若2个因素变化的态势是趋于一致,就可以认为两者关联较大;相反则较低。这种方法既可以用来分析不同因素之间的关联度,又可以评价多层次综合指标系统所描述的整体优劣程度。
2.8.1 数据无量纲化处理 按照灰色关联度分析的要求将青冈栎林下更新指数与4个林分空间结构指数视为一个灰色系统。由于各因子的物理意义不同,使得数据的量纲也有所区别,不方便进行对比,所以对数据进行无量纲化处理,即把各性状的平均值除该因子的全部数据,这样就获得了一个新的数据列。
2.8.2 计算关联系数 以无量纲化后的青冈栎林下更新指数为参考数列X0,林木大小比数、混交度、聚集指数、开敞度分别作为比较数列Xi,i=1,2,…,n,且X0=[X0(1),X0(2),…,X0(k)],Xi=[Xi(1),Xi(2),…,Xi(k)],n为样地数,k是参考系数,在此次研究中n=13、k=4,则关联系数如式(8)。

式中,△i(k)=|X0(k)-Xi(k)|为X0数列与Xi数列在第k点的绝对差值;在所有绝对差值中,△min和△max分别用来表示所有比较数列绝对差值中最小值和最大值;ρ为分辨系数,为提高显著性差异,取值[0,1],一般取0.5,这样能够减少由于最大绝对差值过大而导致的失真。
2.8.3 求关联度 各因子关联系数的均值即为关联度,计算如式(9)。关联度ri越接近于1,说明参考数列与比较数列的相关性越好。

3 结果与分析
3.1 幼树物种组成及重要值分析
林下更新幼树有6种,分属于4科5属,分别是青冈栎、苦槠、多脉青冈、南酸枣、黄檀、马尾松(表1)。青冈栎在更新幼树中占明显优势,是优势树种,重要值为63.09%;其次为苦槠和多脉青冈,重要值均大于10%。
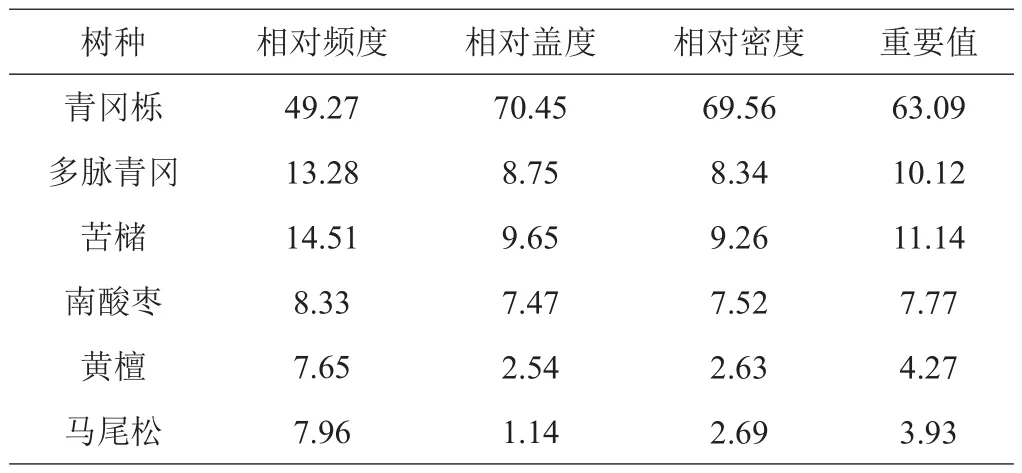
表1 林下更新幼树物种组成及重要值 %
3.2 各项指数的计算结果与分异特征
由13块样地的林分空间结构与林下更新指数统计信息(表2)及其变异系数(表3)可以看出,青羊湖国有林场青冈栎天然次生林各个样地间混交度、幼树的株数密度和树高的变化较大,变异系数分别为0.5281、0.3197、0.2626。大小比数变化较小,变异系数为0.0563。
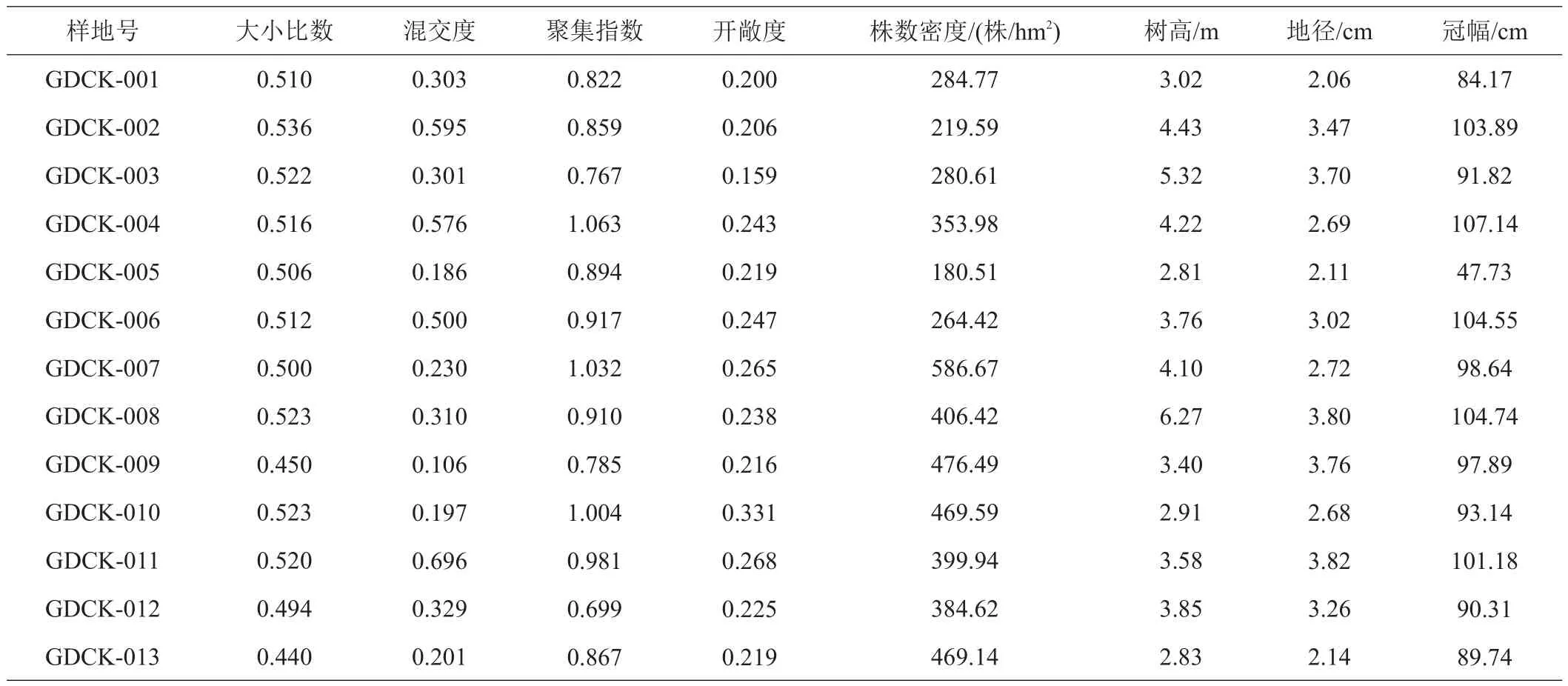
表2 13块样地林分空间结构与林下更新指数的统计信息

表3 样地林分空间结构与林下更新指数的变异系数
3.3 灰色关联度分析
对表2中的13块样地林分空间结构与林下更新指数进行无量纲化处理后,得到新的数据列如表4。
依据无量纲处理取得的数据(表4),得到各比较数列与参考数列的绝对差值△i(k),将△minXi(k)和△maxXi(k)的值代入关联系数公式,取分辨系数为ρ=0.5,得到各林下更新指数与林分空间结构指数的关联系数ξi(k),具体数据见表5~8。

表4 无量纲化林分空间结构与林下更新指数

表5 株数密度关联系数
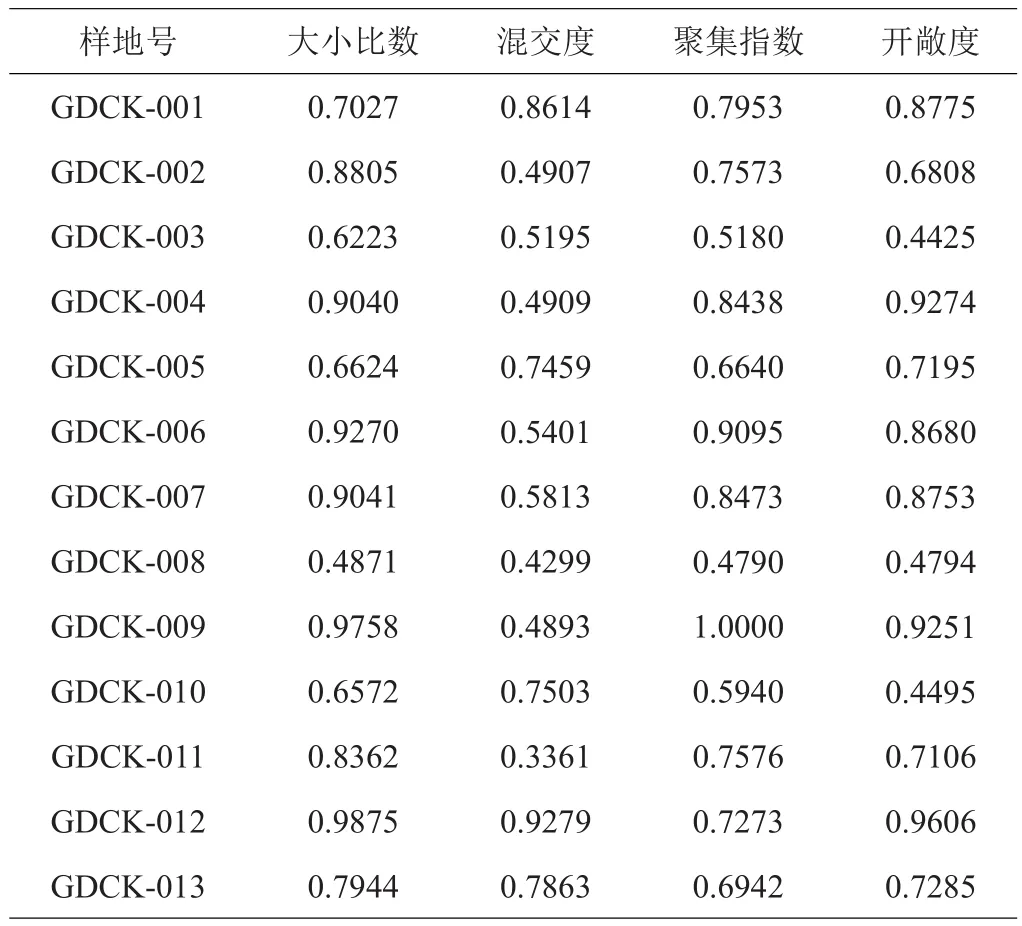
表6 树高关联系数

表7 地径关联系数

表8 冠幅关联系数
由于关联系数是对4个比较数列在某时刻的紧密程度的反映,所以关联度取4个比较数列在各个时刻的关联系数平均值,具体数据见表9。
由表9可知,各林分空间结构指数与林下更新指数间的关联度较高(均大于0.5)。当以株数密度为参考数列时,林分空间结构指数与其关联度表现为开敞度>聚集指数>大小比数>混交度;以树高作为参考数列时,表现为大小比数>开敞度>聚集指数>混交度;以地径作为参考数列时,表现为大小比数>开敞度>聚集指数>混交度;以冠幅作为参考数列时,表现为大小比数>聚集指数>开敞度>混交度。综合所有林分空间结构指数对林下更新指数关联度的平均值,与林下更新关系最为密切的应为林木大小比数(0.7744),说明在青冈栎林中,林木大小比数对林下幼树更新的影响最大,其次为开敞度(0.7468)、聚集指数(0.7396),林木混交度影响相对较小(0.6029)。
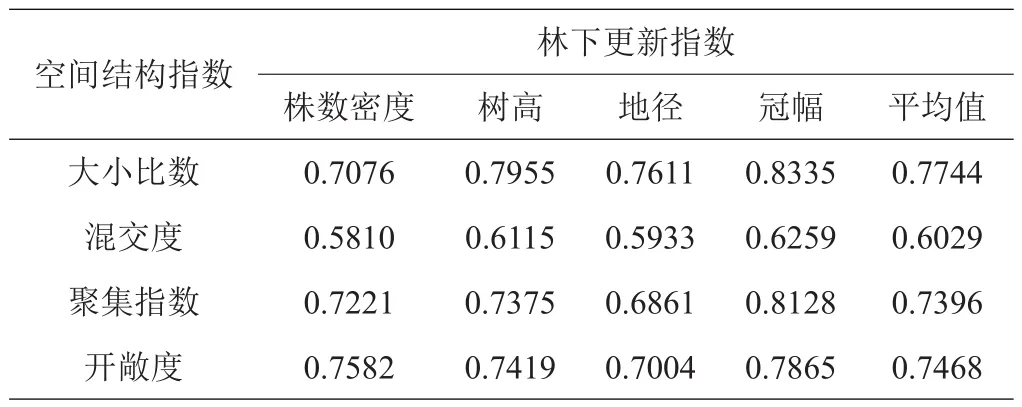
表9 空间结构指数-林下更新指数的关联度
4 结论与讨论
青羊湖林场青冈栎林林下更新幼树有6种,分属于4科5属,分别是青冈栎、苦槠、多脉青冈、南酸枣、黄檀、马尾松。青冈栎在更新幼树中占明显优势,是优势树种,幼树重要值为63.09%;其次为苦槠和多脉青冈,重要值均大于10%。
研究结果表明,青羊湖国有林场青冈栎林木大小比数、开敞度、聚集指数、混交度对其林下更新均有较大影响,其中林木的大小比数为主要因素,因此,在促进青冈栎天然次生林林下更新时,应以调整林木大小分化程度为主,综合考虑林木空间分布格局的调整方案。整个林分大小比数的变化源于乔木层林木个体间的竞争,而这种竞争自然会对林下幼树的更新产生一定的影响;开敞度通过控制林下光照条件,影响林下幼树的生长。有研究表明,开敞度值越大,其林下幼树受光照条件越好,生长越旺盛[24];聚集指数的变化能造成林下枯落物和光斑位置的变化,进而对林下更新幼树的均匀度产生影响;混交度的大小影响乔木层不同树种的种内竞争,有研究表明,混交度高会导致种内个体激烈竞争而形成的自疏和优胜劣汰现象,从而为林下幼树更新释放营养空间[25-26]。
笔者采用灰色关联度分析法量化并比较了每种青冈栎林分空间结构指数对林下更新的影响,但这种方法无法判断正负相关性,所以有一定的局限性。除林分空间结构外,林分非空间结构及环境因子也对林下更新有着较大的影响。因此,在今后研究中,可以将林分空间结构、非空间结构及环境因子一起进行灰色关联度分析,这样可以更加全面地探究影响林下幼树更新的关键因子。
