ω-3多不饱和脂肪酸调控Wnt/β-catenin通路预防神经管畸形的机制研究
2021-10-06游雪云刘艳秋吴敏
游雪云,刘艳秋,吴敏
(1.江西省妇幼保健院产前诊断中心,江西 南昌 330006;2.临川区第一人民医院检验科,江西 抚州 344100)
神经管畸形(NTD)是胚胎神经管无法完成正常闭合而导致的一种先天畸形,其中多不饱和脂肪酸(PUFA)代谢调节紊乱是导致NTD 发生的最重要原因,长链多不饱和脂肪酸(LCPUFA)是神经系统的重要结构组分,尤其是ω-3多不饱和脂肪酸(ω-3-PUFA),但其功能目前尚不清楚。我们的前期研究发现:ω-3-PUFA 代谢产生的类花生酸TXA2、PGl2 能够参与调控营养物质由母亲经胎盘到胎儿的转运,同时促进Hcy 代谢中一些关键基因;Hcy升高可以激活Wnt/β-catenin 信号通路,可能在神经管发育过程中发挥作用。鉴于此,笔者团队于2019年1 月至2020年9 月在江西省妇幼保健院中心实验室以ω-3-PUFA 为切入点,通过不同饲料喂养的孕鼠腹腔注射VPA 建立起VPA 鼠模型,探讨ω-3-PUFA在Hcy 协同下调控Wnt/β-catenin信号通路预防神经管畸形中的功能和作用。
1 材料与方法
1.1 材料 清洁级6 周龄ICR 小鼠(雌鼠50 只,雄鼠25 只)由南昌乐悠生物科技有限公司提供,孕期所需的橄榄油强化饲料由江苏美迪森生物医药有限公司加工。VPA:≥98%,Sigma-Aldrich Co.LLC.总蛋白测定试剂盒:A045-2,由南京建成生物工程研究所提供。Hcy 代谢通路中关键基因(CASC2c、MTHFR、CSE和CSCA2)mRNA表达检测所需试剂为UNIQ-10 柱式Trizol 总RNA 抽提试剂盒,PCR引物由生工生物工程(上海)股份有限公司合成。
1.2 实验方法
1.2.1 qRT-PCR 检测目的基因mRNA的表达通过Primer bank 搜索目的基因的引物,使用NCBI数据库进行blast 比对后,由上海生物有限公司合成。取新鲜的胎盘组织约50mg,Trizol 法提取总RNA 并反转录为cDNA,以表1 操作方法进行反转录,以表2 引物序列和表3 定量PCR 反应体系进行扩增。950C 预变性5min,循环扩增阶段950C 变性15s,580C 退火30s,720C 延伸69s,32 个循环,720C 保温10min。

表1 反转录反应体系

表2 qPCR 引物序列

表3 SYBR Premix Ex Taq 配制荧光定量荧光定量PCR 反应体系
1.2.2 Western blot 法检测目的蛋白的表达 取新鲜胎盘组织约50mg,用冷PBS 洗涤三次,加入500μl 裂解液,匀浆后12000rmp 离心10 min,收集上清液(总蛋白溶液)用BCA 法测定蛋白浓度,于-80℃冰箱保存。10% 分离胶SDS-PAGE 电泳,120V转膜75min,丽春红染色,室温下封闭1h,将转印膜用预先稀释好的—抗于40C 孵育过夜。然后用TBST 于室温下脱色,摇床上洗三次,每次5min。用二抗(用TBST 稀释3000 倍)孵育转印膜30min(室温),然后在脱色摇床上用TBST 洗三次(室温),每次5min。ECL 化学发光1min,曝光显示目的条带,扫描仪扫描,保存数据。
1.3 统计学处理 采用SPSS16.0 软件进行数据分析,并用GraphPad Prism 5 软件作图。定量数据以表示,各组均数间比较,方差齐时采用t 检验,方差不齐时采用近似t 检验,P<0.05 为差异有统计学意义。
2 结果
2.1 qRT-PCR检测CASC2c及β-catenin mRNA的表达水平 ω-3 PUFA 组CASC2c mRNA 相对表达量为对照组的1.166,P=0.0196,差异有统计学意义(P<0.05);ω-3 PUFA 组β-catenin mRNA的相对表达量为对照组的1.272,P=0.0110,差异有统计学意义(P<0.05)。见图1。
2.2 Western blot 法检测CASC2c和β-catenin蛋白表达水平 Western blot 分析可得,ω-3PUFAs 组CASC2c、β-catenin mRNA 相对表达量均较Control组显著增多,差异有统计学意义(P<0.05),见图1。CASC2c蛋白高表达组明显抑制β-catenin (*P<0.05)的表达,见图2。
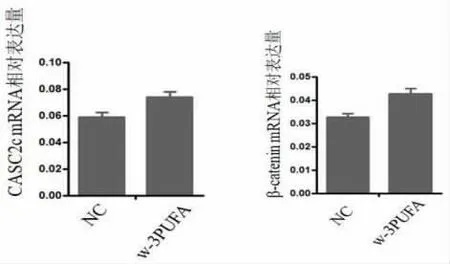
图1 CASC2c和β-catenin mRNA 相对表达量的比较

图2 CASC2c对Wnt/β-catenin 信号通路的影响
3 讨论
神经管畸形(NTD)为胚胎发育过程中出现的疾病,是一类严重的先天畸形,其发病率在新生儿中约占1/1000[1]。研究表明,NTD的发生与遗传、环境均有关系。其中,多不饱和脂肪酸代谢调节紊乱是导致NTD 发生的最重要原因,尤其是ω-3多不饱和脂肪酸(ω-3-PUFA),但其功能目前尚不清楚[2]。Huang 等动物实验表明,ω-3-PUFA的摄入促进了大鼠肝脏中Hcy 代谢通路关键酶的mRNA表达如CASC2c、MTHFR、MATla、CBS、GNMT、MTRR 等,并降低了血浆中Hcy的水平[3]。
本实验以ICR 小鼠为模型系统地研究了VPA及膳食脂肪酸的摄入对叶酸一Hcy 代谢通路中关键基因的表达的影响。以往的研究表明,VPA对叶酸一Hcy 代谢具有抑制作用:Ubeda 等人的动物实验表明VPA 能够升高血清中Hcy的水平,并能抑制MAT 酶(催化Met 转化为SAM)的活性。本实验发现VPA 注射完成6h 或(和)24h 以后抑制了孕鼠肝脏中叶酸一Hcy的代谢中关键基因CASC2cm-RNA表达和蛋白表达,这表明VPA的注射还可通过抑制叶酸-Hcy 代谢通路中以上基因的表达来阻碍Hcy的正常代谢。这些结果表明孕鼠腹腔注射VPA通过抑制孕鼠肝脏中叶酸Hcy 代谢通路中关键基因的表达升高了胚胎中Hcy的水平进而导致了胚胎NTD的发生。以往动物实验也表明PUFA的摄入能够显著降低血液中Hcy的水平[4]。在本实验中我们发现孕鼠橄榄油的摄入也能够有效地阻止VPA对胚胎中Hcy的升高作用,ω-3-PUFA的摄入能够升高叶酸-Hcy 代谢通路中关键基因(如CASC2c、CBS、MTRR 等)的mRNA和蛋白表达。以上研究结果表明ω-3-PUFA的摄入可能通过缓解VPA对孕鼠肝脏叶酸一Hcy 代谢通路中CASC2c、DHFR、MTRR、GAMT、CBS及CSE表达的抑制作用使胚胎中的Hcy水平维持正常水平进而降低了胚胎中VPA 诱导的NTD的发病率。
近年的研究表明Hcy与胚胎的先天畸形密切相关。Felkner 等人在德克萨斯州一墨西哥边境国家开展了一项病例对照实验发现血清中Hcy水平也与NTD的发病风险呈正相关[5]。中国以及非洲国家的研究也观察到了相似的结果[6,7]。然而关于Hcy是否能直接导致神经管畸形的发生尚存在争议。Greene 等人的动物实验表明用Hcy 硫代内酯培养小鼠胚胎并不能够升高其NTD的发病率,这表明Hcy 可能不是直接诱导NTD的发生,生育NTD 胎儿的母亲具有较高的Hcy水平可能只是Hcy 代谢紊乱的一种反映[8]。
诱发氧化应激及细胞凋亡是高同型半胱氨酸血症主要的致病机制之一,但其中的具体机制还不是非常清楚,有报道认为高同型半胱氨酸血症可以激活Wnt/β-catenin 信号通路[9]。Wnt/β-catenin 信号转导通路是一条在生物进化中极为保守的通路。在正常的体细胞中,β-catenin 只是作为一种细胞骨架蛋白在胞膜处与E-cadherin 形成复合体对维持同型细胞的黏附、防止细胞的移动发挥作用。只有当细胞外Wnt 信号分子与细胞膜上特异性受体Frizzled蛋白结合激活胞内的dishevelled (散乱的)蛋白导致GSK3B 失活,使α-catenin 避免被磷酸化而遭降解的命运,β-catenin 才能在胞质中积累起来。当胞质中β-catenin的浓度达到一定水平时,就可向细胞核转移。在胞核中β-catenin与转录因子家族Tcf/Lefs 结合,可激活cyclinD1和cmyc 等原癌基因而导致细胞的增殖、分化、成熟。显然,在这条信号通路中β-catenin 发挥了重要作用,其在胞质中积累继而向胞核的转位,被认为是该信号通路被激活的标志。Wnt/β-catenin 信号通路在调节细胞增殖,细胞凋亡等活动中发挥重要作用,该信号通路的失衡会导致许多疾病,如神经管畸形,心脏发育畸形,神经退行性疾病等。神经管畸形是与高同型半胱氨酸血症相关的一类出生缺陷,已有文献报道高同型半胱氨酸血症导致神经管畸形的机制也诱发了氧化应激,从而激活了在发育中起到重要作用的Wnt/β-catenin 信号通路。近年研究发现,Wnt/β-catenin 信号通路是哺乳动物中枢神经系统(CNS)发育的关键信号通路之一。当Wnt/β-catenin 信号通路处于持续激活状态,脑组织中NSCs 一直处于有丝分裂周期之内,NSCs的数目也持续增加。Zhang 等对体外培养的小鼠NSCs(NE-4C)进行体外冲击波治疗(rESWT),后者激活了Wnt/β-catenin 信号通路,随后检测发现,经过电刺激的小鼠NSCs 不断增殖,并且一直处于未分化状态[10]。Qi 等从SD 大鼠中分离NSCs,并且在含有0.3%氧气的微氧培养系统中尽心培养,结果显示,在缺氧条件下,NSCs 内β-catenin水平升高与NSCs 持续增殖呈正相关[11]。以上研究均证实,Wnt/β-catenin 信号通路可促进NSCs 增殖,在NSCs 发育过程中扮演重要角色。最近又有研究报道提示该通路信号转导是作为神经元细胞形成的开关,因此对于神经元细胞的维持和增殖是重要的,特别是在神经管畸形(NTD),预防出生缺陷的研究中具有重要意义[12,13]。
ω-3多不饱和脂肪酸(ω-3-PUFA)属于长链脂肪酸的主要类别之一,是PPARs的潜在激活剂,Chang 等研究表明,肌肉中ω-3多不饱和脂肪酸富集增加了肌肉中的IMF 含量,影响脂肪生成相关基因的表达,还有证据表明ω-3多不饱和脂肪酸抑制细胞培养中的Wnt/β-catenin 信号通路,诱导PPARs表达和Wnt 信号传导[14,15]。本实验的研究结果表明,VPA 能够抑制孕鼠肝脏中PUFA的合成,并能造成肝脏及胚胎中叶酸一 Hcy 代谢通路中关键基因表达的紊乱,进而影响叶酸-Hcy的正常代谢。而ω-3-PUFA的摄入能有效缓解VPA 所引起的孕鼠肝脏及胚胎中叶酸-Hcy 代谢通路一系列基因mRNA和蛋白的表达紊乱,这表明PUFA的缺乏及叶酸一Hcy 代谢紊乱是导致NTD 发生的重要因素,而PUFA的摄入能够通过弥补机体PUFA的不足及调控叶酸-Hcy的正常代谢预防NTD的发生。
