不同紫罗兰品种对弱光胁迫的响应
2021-09-26张一丹周长娥郭绍霞
张一丹,周长娥,郭绍霞
(1.青岛农业大学园林与林学院,山东青岛 266109;2.青岛市农业技术推广中心,山东青岛 266071 )
光是影响植物生长发育的重要环境因子,光合作用将光能转化为有机物,为植物光合作用提供能量[1];同时,植物感受光信号因子调控形态建成,从而使植物达到最佳生长状态[2-3]。
紫罗兰(Matthiolaincana)为十字花科、紫罗兰属二年生植物,花大色艳,开花时间长,常用作冬季日光温室切花栽培植物[4-5]。在冬季北方地区,随着时间与空间的改变光照强度变化幅度较大,严重影响植物生长,降低其品质和产量。研究发现在弱光环境下,植物进行光合作用的能力减弱,电子传递速率减慢,促使植物体内活性氧产生,使细胞膜系统受到损伤及代谢过程受阻[6],降低了植物对矿质营养的摄入和同化产物的积累,导致植株矮小、干物质量减少等[7]。马铃薯(Solanumtuberosum)在持续弱光环境下生长后,发现其茎节细长、叶片变薄、叶面积减小,同时叶绿素含量降低、生长速度迟缓[8]。另外研究表明,在弱光胁迫下黄心菜(Brassicapekinensis)叶片的PSII实际光化学量子效率(φPSII)和PSII电子传递速率(ETR)与对照相比显著下降,说明弱光可以导致植物光合作用下降[9];关晓溪等[10]研究发现弱光胁迫下烟草(NicotianatabacumL.)幼苗的可溶性蛋白含量下降,丙二醛(MDA)含量升高,使膜系统受损;薛维纳等[11]研究表明,当植物受到严重弱光胁迫时,抗氧化酶活性受到抑制,活性氧不能被及时清除,从而对植物体形成氧化损伤。因此,测定不同品种紫罗兰的生长和生理指标,筛选出耐弱光的品种,应用于冬季温室栽培,提高其观赏品质和市场竞争力,是当前设施栽培生产亟待解决的问题之一。
1 材料与方法
1.1 试验材料
供试的3个紫罗兰品种分别为‘卡尼陀’‘辉煌’和‘贵族’,购买自浙江虹越花卉股份有限公司。试验地点为青岛农业大学人工气候室,育苗基质为园土∶蛭石∶草炭按2∶1∶1体积比混合。
1.2 试验方法
挑选籽粒饱满的种子浸种催芽后于2018年12月将紫罗兰种子播种于128孔的穴盘内,正常光温管理,光照强度(500 μmol·m-2·s-1),温度18 ℃/11 ℃(昼/夜),光周期12 h/12 h(昼/夜),空气相对湿度保持在60%~80%。每周浇3次水,浇水均匀而充足。每7 d浇1次Hoagland营养液,以保证养分供给,管理期间注意温度、光照和通气条件等的控制。26 d后(约2~4片真叶)将生长健壮且长势一致的幼苗移栽至盆口直径15 cm、高18 cm的塑料盆内,每盆2株苗,待长出6~8片真叶后在人工气候室内进行弱光(100 μmol·m-2·s-1)处理,其他生长条件均不变。采用随机分组设计,在弱光处理后的0 d、1 d、3 d、5 d、7 d时分别选取功能叶(从上往下数第3~4叶)进行指标测定,重复3次。
1.3 指标测定
1.3.1 生长指标的测定
用尺子(0.1 mm)测量紫罗兰根茎部到植株生长点的距离即为株高;用清水将紫罗兰根部冲洗干净后将表面水分吸干,称量紫罗兰地上部和地下部鲜重;将紫罗兰幼苗放入105 ℃烘箱中杀青20 min,在85 ℃下烘干至恒重,用万分之一电子天平称量地上部干重和地下部干重。
1.3.2 生理指标的测定
超氧化物歧化酶(SOD)活性、可溶性蛋白含量、丙二醛(MDA)含量以及叶绿素含量的测定均采用王学奎[12]描述的方法。
叶绿素荧光测定:选择晴朗少云的天气(9:30至11:30),叶片暗适应30 min后用Pocket PEA(Hansatech Instruments Ltd.,UK)测定叶片的叶绿素荧光参数,根据最小荧光值(Fo)、最大荧光值(Fm)、可变荧光值(Fv)及稳态荧光(Fs)等计算PSII实际光化学量子效率(φPSII)和PSII电子传递速率(ETR)[13-14]。
1.3.3 耐弱光性综合评价
采用隶属函数法[15]进行综合比较。与抗弱光性呈正相关的指标诸如超氧化物歧化酶(SOD)活性、可溶性蛋白质含量等采用公式:
U(Xijk)=(Xijk-Xmin)/(Xmax-Xmin)
与抗弱光性呈负相关的指标MDA含量等采用公式:
U(Xijk)=1-(Xijk-Xmin)/(Xmax-Xmin)
上述公式中:U(Xijk)为第i个品种第j个取样阶段第k项指标的隶属度,且U(Xijk)∈[0,1];Xijk表示第i个品种第j个取样阶段第k个指标测定值;Xmax、Xmin为所有参试品种中第k项指标的最大值和最小值。用以上公式求出不同品种的各项指标参数的隶属函数值,再求各值和的平均值得其综合评价值。
1.4 数据处理
采用Microsoft Excel 2003和DPS 7.5 (LSD法进行方差分析)软件进行数据统计与分析。
2 结果与分析
2.1 弱光胁迫对不同紫罗兰品种生长的影响
由表1可以看出,3个紫罗兰品种弱光胁迫处理后7 d与0 d相比,其株高、地上部鲜重、地下部鲜重、地上部干重和地下部干重均有不同程度的增长,‘卡尼陀’的地上部鲜重、地上部干重和地下部干重增加最显著,分别增加了136.8%、67.7%、40.0%。
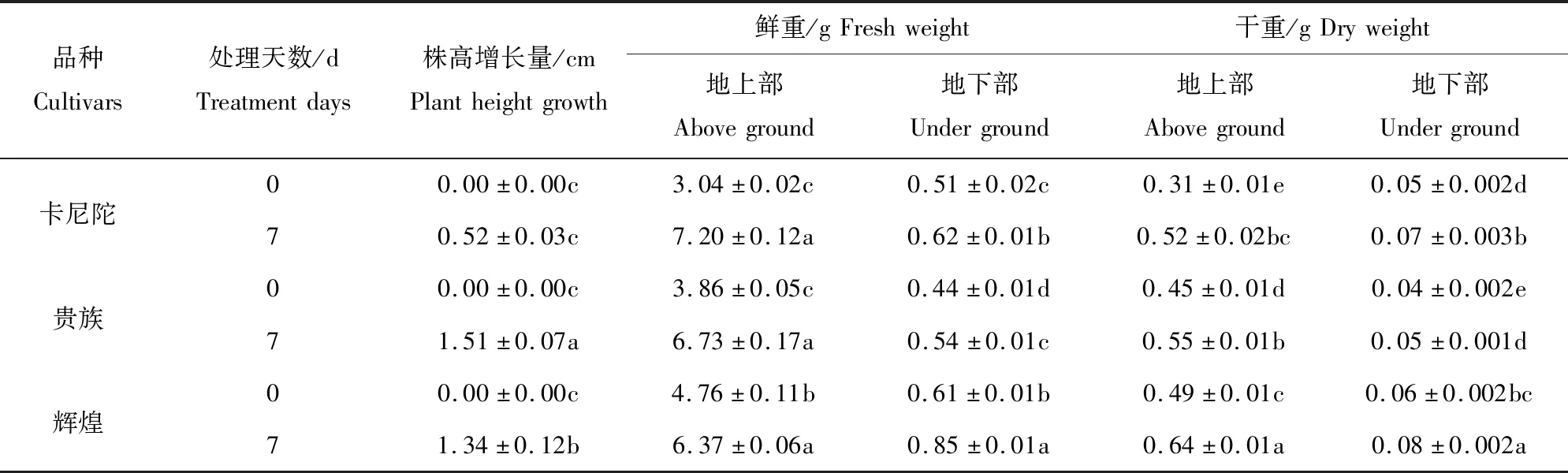
表1 弱光胁迫对不同紫罗兰品种生长的影响Table 1 Effects of low light stress on the growth of different violet cultivars
2.2 弱光胁迫对不同紫罗兰品种抗氧化酶(SOD)活性的影响
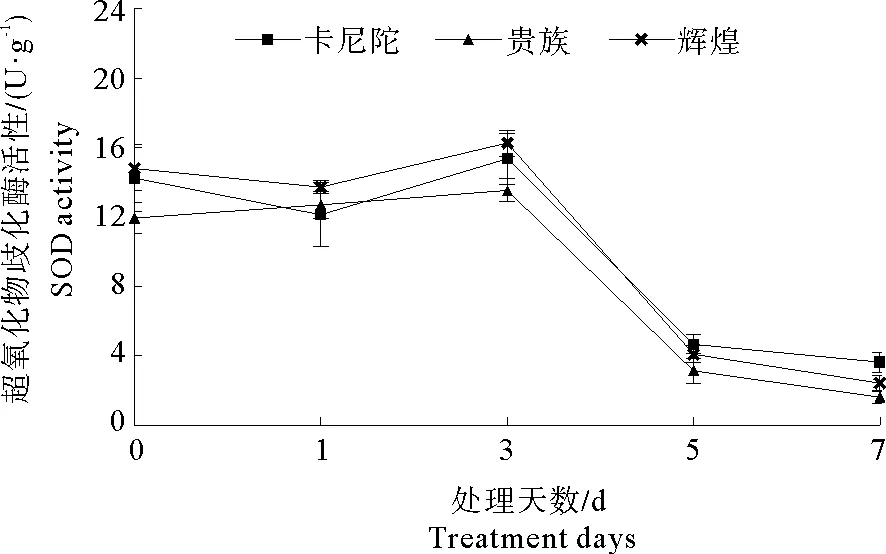
图 1 弱光胁迫对不同紫罗兰品种SOD活性的影响Fig.1 Effects of low light stress on SOD activity of different violet cultivars
2.3 弱光胁迫对不同紫罗兰品种可溶性蛋白含量的影响
可溶性蛋白是植物体内一种重要的渗透调节物质,对生物膜起到保护作用,在逆境胁迫下,植物体为抵御逆境胁迫会增加自身可溶性蛋白含量,以减缓逆境对其产生的不良影响[17]。由图2可以看出,弱光胁迫处理0~3 d,‘卡尼陀’和‘贵族’可溶性蛋白含量先上升后下降,而‘辉煌’可溶性蛋白含量一直上升。处理3~7 d时,3个紫罗兰品种的可溶性蛋白均呈下降趋势。总体来看,紫罗兰叶片中可溶性蛋白的含量先升高后下降,这说明,在一定程度的弱光胁迫下,植物体能够通过自身适应反应增加可溶性蛋白含量,减轻弱光胁迫对其产生的不良影响,但随着弱光胁迫时间的延长,植物自身无法进行应激调节,对植物造成了不可逆的伤害。
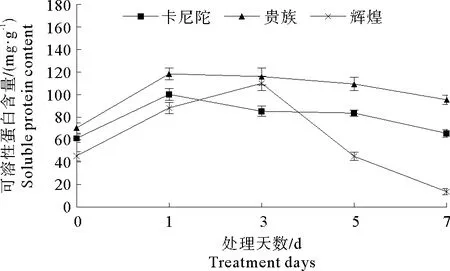
图2 弱光胁迫对不同紫罗兰品种可溶性蛋白含量的影响Fig.2 Effects of low light stress on soluble protein content of different violet cultivars
2.4 弱光胁迫对不同紫罗兰品种丙二醛(MDA)含量的影响
从图3可以看出,随着弱光处理时间的延长,从总体趋势来看,3个紫罗兰品种在弱光胁迫下MDA含量都呈现上升的趋势,但在程度上存在差异。在处理5~7 d时,‘辉煌’MDA含量急速上升,‘贵族’和‘卡尼陀’MDA含量缓慢增加;处理7 d与0 d相比,3个紫罗兰品种的MDA含量显著增加,‘卡尼陀’‘贵族’‘辉煌’比对照分别增加了70.9%、36.7%,42.4%,说明弱光胁迫对‘辉煌’的细胞膜伤害最大。
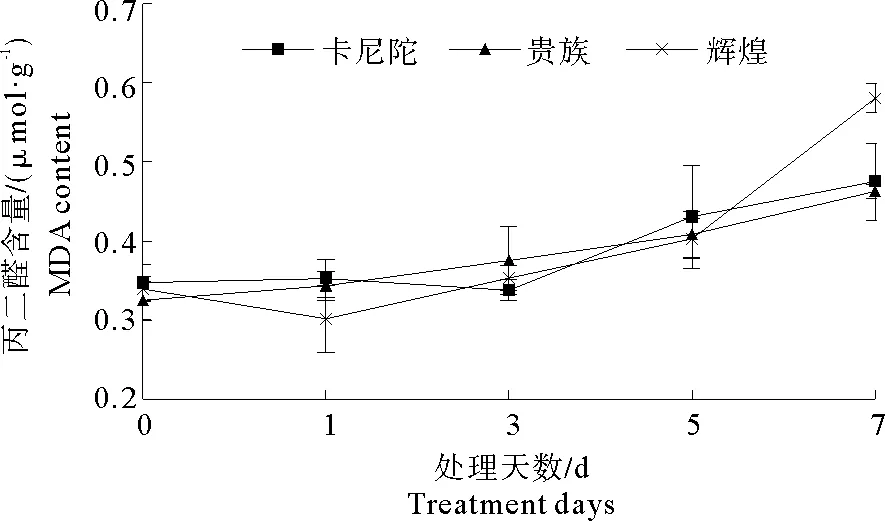
图3 弱光胁迫对不同紫罗兰品种丙二醛含量的影响Fig.3 Effects of low light stress on MDA content of different violet cultivars
2.5 弱光胁迫对不同紫罗兰品种总叶绿素含量的影响
随着弱光胁迫处理时间的延长,3个紫罗兰品种的总叶绿素含量均呈下降趋势(图4)。其中,与0 d相比,处理1~3 d时‘卡尼陀’和‘贵族’总叶绿素含量显著下降,分别下降了43.2%和40.5%,而‘辉煌’总叶绿素含量下降不显著;处理7 d与0 d相比‘辉煌’‘卡尼陀’和‘贵族’的总叶绿素含量均显著降低,分别降低了30.9%、61.5%和50.1%。
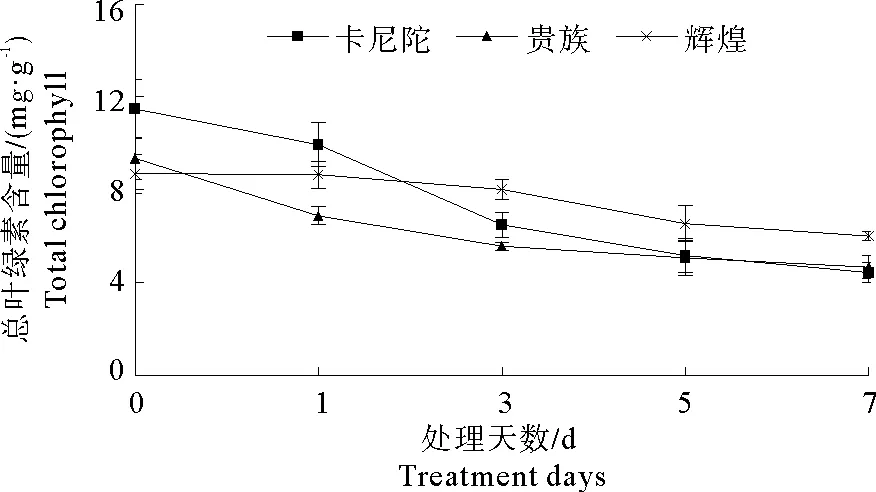
图4 弱光胁迫对不同紫罗兰品种总叶绿素含量的影响Fig.4 Effects of low light stress on total chlorophyll content of different violet cultivars
2.6 弱光胁迫对不同紫罗兰品种叶绿素荧光参数的影响
如图5所示,弱光胁迫下3个紫罗兰品种的PSII实际光化学量子效率(φPSII)和PSII电子传递速率(ETR)显著下降(见图5A、5B),处理7 d与0 d相比‘卡尼陀’‘贵族’‘辉煌’的φPSII分别降低了27.8%、38.2%、44.6%;ETR分别降低了37.7%、51.4%、63.6%,说明弱光胁迫使紫罗兰叶片的PSII反应中心受到影响,电子传递速率下降,降低了植株对光能的利用效率。
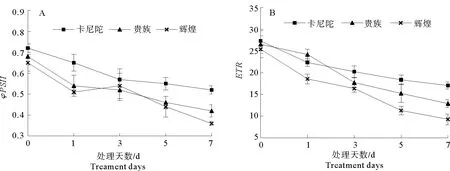
图5 弱光胁迫对不同紫罗兰品种叶绿素荧光参数φPSII和ETR的影响Fig.5 Effects of low light stress on chlorophyll fluorescence parameters φPSIIand ETR of different violet cultivars
2.7 耐弱光性综合评价
运用隶属函数法对3个品种进行耐弱光的综合评价(表2)。根据平均隶属函数值越大其耐弱光能力越强得出3个品种的耐弱光能力顺序为:‘卡尼陀’>‘贵族’>‘辉煌’。
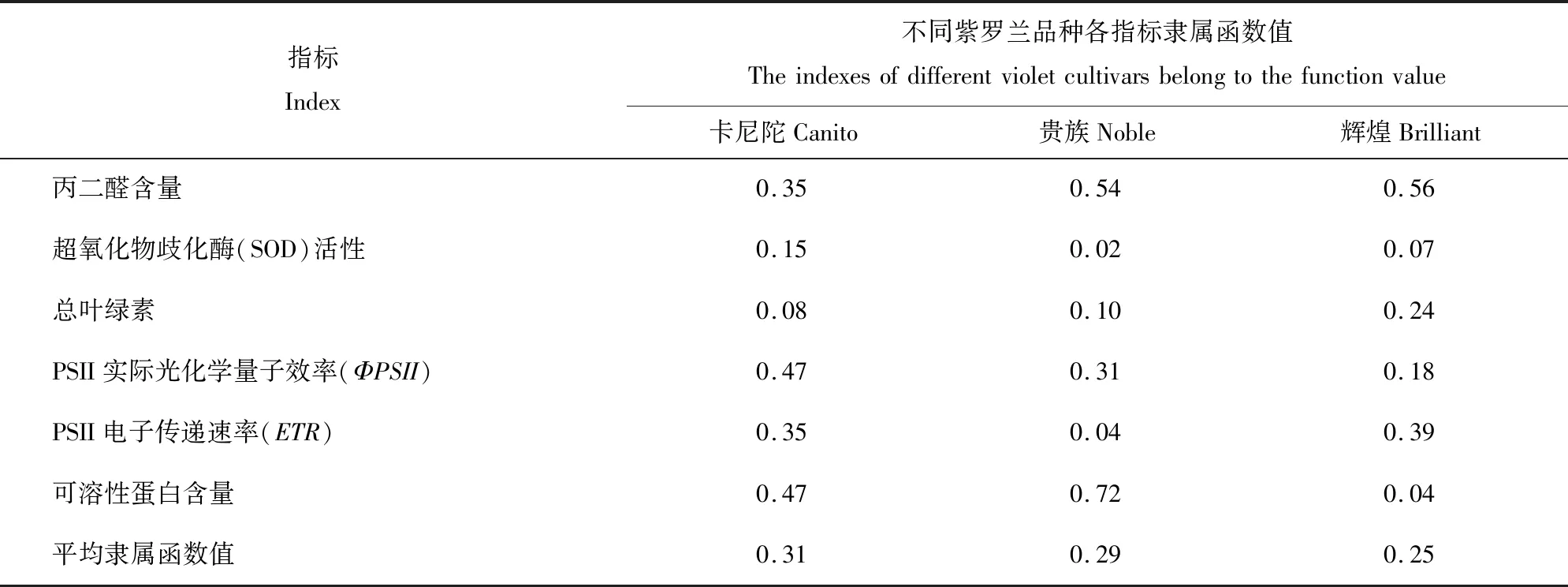
表2 紫罗兰品种的耐弱光性排名Table 2 Low light resistance of M. incana cultivars
3 讨论
在弱光胁迫下,植物为了适应环境,会调节自身形态特征[18],比如植物株高增加,叶片干重减少等都是植物为了适应弱光胁迫所采取的反应机制。干物质的积累与碳的净同化密切相关,因此在植物的生长过程中,弱光往往会导致植物干物质积累减少和产量下降[19]。杨柳等[20]对‘狭叶红景天’幼苗(Rhodiolakirilowii)的研究中发现,遮阴处理与对照相比,植株株高显著增加,总生物量、根干重、根冠比显著减少;刘科等[21]研究发现遮光使超级杂交稻干物质积累量比普通杂交稻品种和常规稻品种下降明显,说明超级杂交稻对弱光的抵抗能力较差。在本试验中,弱光胁迫下不同品种紫罗兰株高、地上部鲜重、地下部鲜重、地上部干重和地下部干重较0 d都有所增长,但增长幅度不同,其中‘卡尼陀’的地上部鲜重、地上部干重和地下部干重增长量最多,因此,‘卡尼陀’的干物质积累量最多,受弱光胁迫的影响最小。
不良环境胁迫下,植物光合色素含量发生明显变化,从而改变其光合能力[22]。叶绿素作为绿色植物不可或缺的色素,在植物光合作用中发挥着重要作用[23]。植物的叶绿素荧光参数将植物在光合作用中的部分变化信息呈现出来,比如光能的吸收、传递等[24]。王成孜等[25]研究表明弱光胁迫下水稻(OryzasativaL.)叶片的PSII实际光化学量子效率(φPSII)及电子传递速率(ETR)显著降低,这与本试验研究结果一致。本研究中,弱光胁迫处理下紫罗兰叶片的总叶绿素含量、φPSII和ETR显著下降,这可能是由于光合色素含量的多少将直接影响植物的光合作用,在弱光胁迫下,紫罗兰叶片获得的光能减少,阻碍了其PSII光化学反应的进程。
细胞膜是维持细胞正常代谢的必要屏障,在控制细胞内外物质运输和交换上发挥了重要作用[26-27]。植物处于逆境胁迫下,其体内会产生较多的活性氧,而SOD等抗氧化酶、渗透调节物质对活性氧的清除和保护膜结构的完整性中发挥重要作用,并减轻活性氧带来的伤害[28]。活性氧积累会造成膜脂过氧化,膜脂过氧化产物丙二醛 (MDA)是反映植物细胞膜受伤害程度的重要指标之一[29-30]。本试验结果表明,弱光胁迫下,3个紫罗兰品种叶片SOD酶活性显著下降、MDA的含量显著增加、可溶性蛋白含量呈下降趋势。可以说明,随弱光胁迫时间增长,细胞中活性氧自由基增加,导致抗氧化酶结构破坏或使其活性降低,这与尹立辉[31]对非洲紫罗兰(Saintpauliaionantha)的耐阴性研究中的结果一致。
综合分析得出3个紫罗兰品种在弱光下的适应程度为‘卡尼陀’>‘贵族’>‘辉煌’。本试验全程在人工控制的环境下进行,虽然有利于试验操作和得出结果,但是若想揭示自然条件下综合环境因子对于紫罗兰的伤害机制仍有一定的局限。弱光胁迫的同时往往伴随着其他的逆境因子,例如水分胁迫、低温胁迫等。因而,今后针对植物各种附加逆境因子综合作用的研究可能对于我们引种、选育更具有实际意义。
