黄淮海50份大豆种质资源SSR遗传多样性分析
2021-09-23杨青春舒文涛李金花常世豪张东辉张保亮
李 琼, 耿 臻, 杨青春, 舒文涛, 李金花, 常世豪, 张东辉, 张保亮
(周口市农业科学院, 河南 周口 466001)
大豆(Glycinemax)起源于中国,是我国重要的经济作物,是植物蛋白、脂肪的重要来源[1]。黄淮海地区是我国大豆主产区之一,种植面积占全国的三分之一。大豆种质资源是大豆品种改良的前提和基础。黄淮海区域大豆种质资源的筛选对遗传背景的探索和遗传资源的充分利用具有积极影响[2-3]。本研究通过对50份黄淮海大豆种质资源的遗传多样性分析,为该片区大豆种质资源的利用和开发提供必要信息,为育种材料的选择提供重要的理论依据。
SSR分子标记技术具有多态性丰富、重复性好和突变率高等优点,蕴含着多样的多态性信息,是研究遗传多样性的手段之一[4-5]。目前,SSR广泛应用于大豆遗传多样性的研究过程中。王帅等[6]用SSR标记分析大豆疫霉根腐病抗原的遗传多样性,筛选出5对高多态性SSR引物,构建出60份抗性材料的指纹图谱。李志江等[7]用SSR标记对黑龙江种植大豆品种的遗传多样性进行分析,并筛选出与性状关联的SSR引物。金尚昆等[8]用SSR标记对黄淮海地区284份材料进行基因型分析,检测出北京、河北材料多样性最高。本试验基于42对SSR引物对黄淮海50份大豆种质进行遗传多样性分析,明确南、北居群间的亲缘关系,为黄淮海大豆种质资源的选择和利用奠定理论基础。
1 材料与方法
1.1 材 料
供试材料主要来源于周口市农业科学院大豆原始材料收集库。将表1中黄淮海大豆种质资源根据地理位置分为两大居群,如表2。居群1(POP-Ⅰ)为33份黄淮海南部大豆种质资源,居群2(POP-Ⅱ)为17份黄淮海北部大豆种质资源。
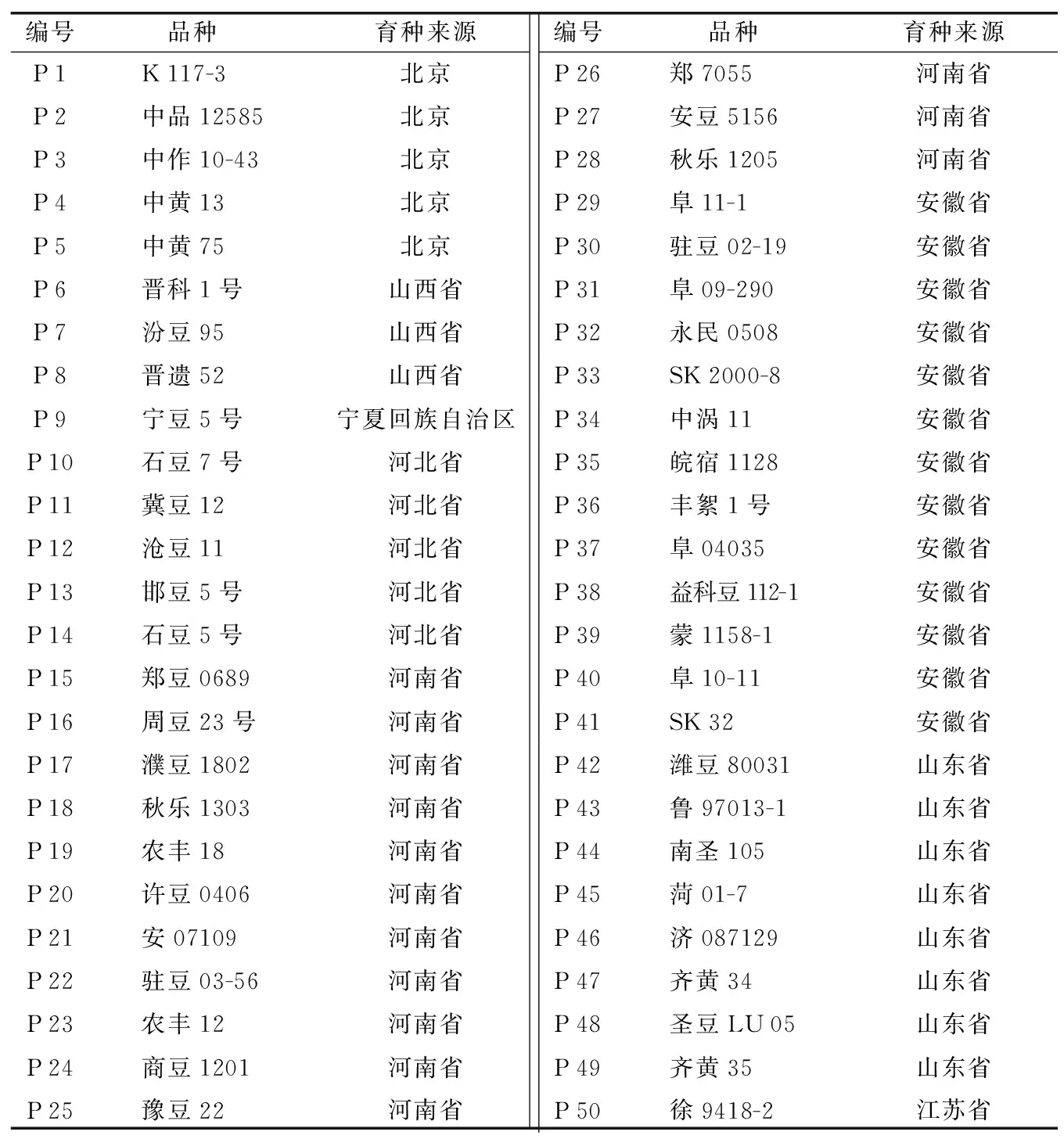
表1 黄淮海50份大豆种质育种来源情况

表2 黄淮海50份大豆种质资源在2个居群中的分布情况
1.2 方 法
1.2.1大豆DNA的提取
选取籽粒饱满的种子种植在育苗盘中,放置在培养箱内(25 ℃,光照8 h·d-1)培育出2片复叶。采集鲜嫩叶片按照CTAB法[9]提取DNA。采用紫外分光光度计检测DNA的浓度,并稀释至60 ng·μL-1,放置于-80 ℃冰箱备用。
1.2.2引 物
采用《2019年大豆国家区试品种报告》中检测参试品种遗传关系所用的42对引物[10],由上海生物工程技术服务有限公司合成。
1.2.3PCR扩增及电泳
采用10 μL PCR反应体系,2×TransTaq®-T PCR SuperMix 5 μL,上、下游引物各0.5 μL,DNA模板1 μL,双蒸水3 μL。反应程序:94 ℃预变性5 min,94 ℃变性40 s,55 ℃退火50 s,72 ℃延伸50 s,共循环35次,72 ℃延伸10 min,4 ℃冰箱保存待用。PCR产物用6%变性聚丙烯酰胺凝胶电泳检测,电压220 V,时长2.5 h,印染法染色,拍照。
1.2.4数据分析
采用人工读带方法,将电泳图片上可重复、易分辨的条带标记为“1”,同一位置无带标记为“0”,建立原始数据矩阵。利用POPGEN 32软件对所选群体的观测基因数(Na)、有效等位基因数(Ne)、Shannon’s多态性信息指数(I)[11]、遗传距离(GD)、遗传一致度(GI)。计算多态性信息含量(PIC)[12]、NTSYS-PC软件计算遗传相似系数(Gs),按照非加权配对法UPGMA和SHAN程序进行聚类分析,Eigen程序进行主成分分析(PCA)。
2 结果与分析
2.1 引物标记多态性分析
所用的42对引物(见表3)共扩增出203条带,扩增片段长度在102~340 bp之间,每对引物扩增条带数为2~10个,平均为4.83个,多态性条带为189个,多态位点百分率(PPB)为97.54%。其中,satt 184、satt 194、satt 373和satt 590的多态位点百分率较低,分别为85.71%、75.0%、83.33%和60%。多态性信息含量(PIC)变幅为0.115 9~0.989 4,平均为0.721 6。PIC是用来衡量基因变异程度的指标。 Botstein等[13]认为,PIC<0.25时,为低度动态性信息引物;0.25
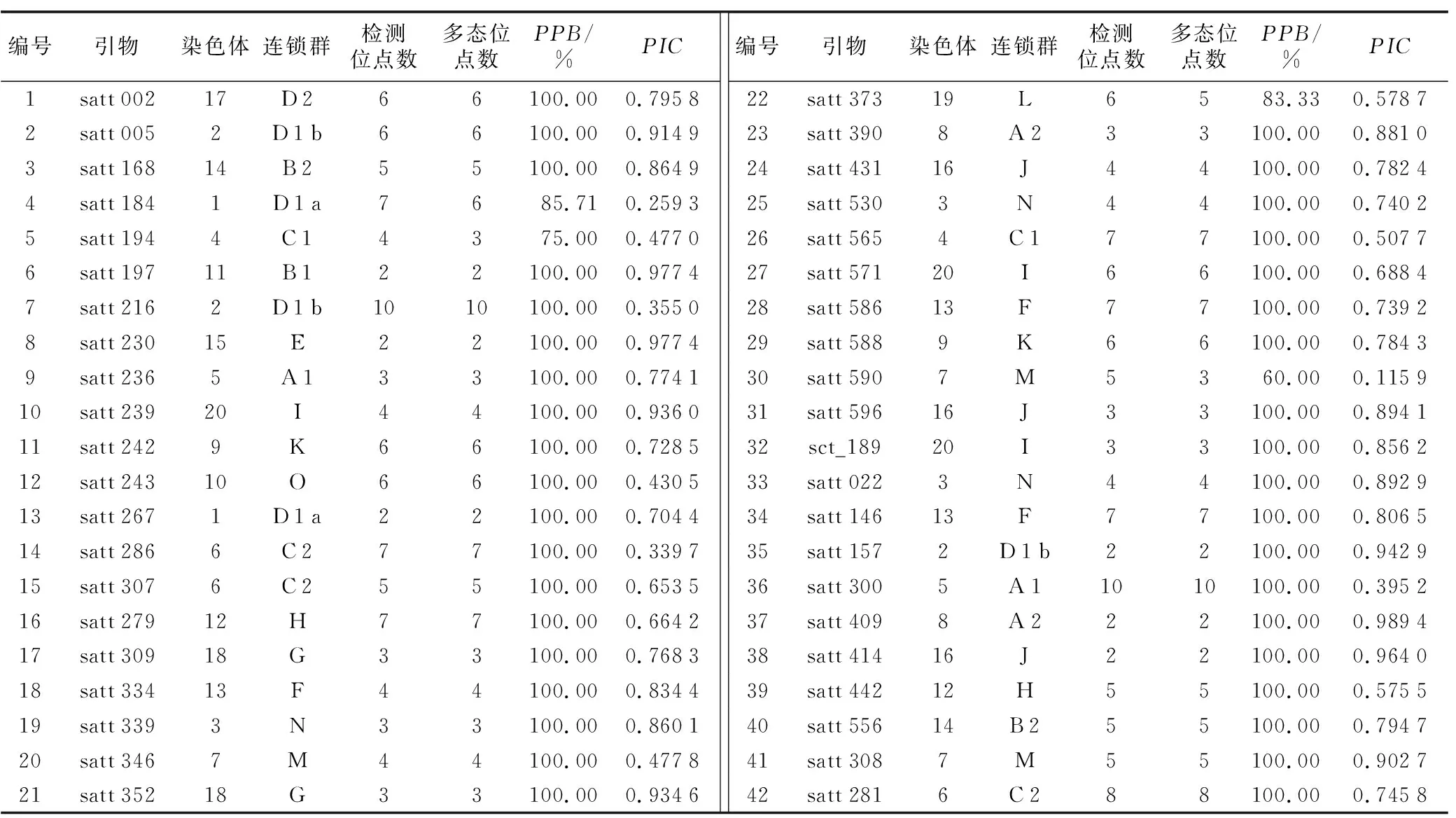
表3 SSR引物多态性信息
2.2 黄淮海2个大豆种质居群的遗传多样性和遗传变异分析
由表4可知,供试的50份大豆种质平均等位基因数(Na)为1.975 4,平均有效等位基因数(Ne)为1.417 2,平均Nei’s基因多样性指数(H)为0.267 2,平均Shannon’s信息指数(I)为0.421 2,多态位点百分率为97.54%。黄淮海北部大豆种质的有效等位基因数(Na)、Nei’s基因多样性指数(H)、Shannon’s信息指数(I)均大于黄淮海南部。黄淮海北部大豆种质资源的遗传多样性较黄淮海南部高,表明黄淮海北部大豆种质资源较为丰富。两个大豆居群的总遗传多样度(Ht)为0.270 4,居群内遗传多样度(Hs)为0.259 1。居群间遗传分化系数(Gst)为0.141 7,基因流(Nm*)为3.03。由此可见,14.17%的遗传变异在两个居群之间,85.83%的遗传变异在居群内部。黄淮海两个大豆居群间差异较小,但居群内部个体差异较大,居群间基因从一个居群到另一个居群的基因运动十分丰富。

表4 不同地理来源的大豆种质的遗传多样性分析
2.3 黄淮海2个大豆种质居群间的遗传距离
利用POPGEN软件处理数据结果可知,黄淮海大豆种质资源南、北两个居群的遗传相似度(GI)为0.869 6,遗传距离(GD)为0.210 9。由此可见,两个居群间遗传相似性高,遗传距离较近。总体来说,黄淮海南、北两个大豆居群遗传背景较为相似,亲缘关系较近。
2.4 黄淮海大豆种质资源聚类分析和主成分分析
利用NTSYS软件对42对SSR引物所检测出的188个多态性位点数据进行分析,计算并建立遗传相似度矩阵,进行UPGMA聚类(图1)。由图1可知,50份黄淮海大豆种质的遗传相似系数(GS)的变化范围为0.62~1。遗传相似系数(GS)在0.620处,50份材料被分为两个类群。类群Ⅰ包括36份材料,类群Ⅱ包括14份材料。其中,POP-Ⅰ中33份材料中有2份材料:石豆7号(P 10)、南圣105(P 44)被聚到类群Ⅱ中。POP-Ⅱ中17份材料中有5份材料P9(宁豆5号)、冀豆12(P 11)、邯豆5号(P 13)、安07109(P 21)、安豆5156(P 27)被聚到类群I中。其余的43份(占总体种质资源的86%)大豆种质材料所处类群与POP-Ⅰ、POP-Ⅱ的居群分类情况一致。结果表明,两个居群(POP-Ⅰ、POP-Ⅱ)中的大部分材料与UPGMA聚类结果一致,仅存在7份材料在黄淮海南、北部之间互相交错。因此,可推测黄淮海大豆种质资源与地理分布具有显著的相关性的同时,一定程度上与育种来源遗传背景有所关联。
利用NTSYS软件对42对SSR引物所检测出的188个多态性位点数据分析所得遗传相似系数矩阵进行主成分分析(Principal Component Analysis,PCA),得出二维点散图(见图2)和三维立体点散图(见图3)。由图2、图3可以看出,两者中聚类结果与UPGMA聚类(图1)结果完全一致。进一步证明了上述分成两个类群结构的准确性。
3 讨 论
3.1 黄淮海大豆种质资源遗传多样性和亲缘关系
遗传多样性评价参数能够反映种内不同居群之间及居群内部不同个体之间的遗传变异水平[14-15]。Shannon’s信息指数(I)是衡量种群或地区种质资源遗传多样性指数的标准。Nei’s基因多样性指数(H)是通过计算遗传距离来分析种质遗传多样性。本研究从处在黄淮海南、北部不同地理位置的两个居群中选择50份种质材料进行SSR遗传多样性分析。大豆总体种质平均Nei’s基因多样性指数(H)为0.267 2,平均Shannon’s信息指数(I)为0.421 2。表明黄淮海大豆遗传多样性较为丰富,具有多样的遗传关系。50份种质的平均等位基因数(Na)为1.975 4,平均有效等位基因数(Ne)为1.417 2。其中,POP-Ⅰ中33份种质的平均等位基因数(Na)为1.926 1,平均有效等位基因数(Ne)为1.401 6;POP-Ⅱ中17份种质的平均等位基因数(Na)为1.926 1,平均有效等位基因数(Ne)为1.414 0。等位基因数和有效等位基因数可以反映等位基因分布的均匀程度,2个指标越接近,说明等位基因分布越均匀[16-18]。因此,供试的50份大豆种质间和两个居群内部各个种质间的等位基因分布都较为均匀,有利于基因多样性的稳定遗传。
3.2 种质资源的聚类与地理来源的关系
刘新星等[19]、魏苗等[20]、董俊丽等[21]的研究认为,SSR聚类与地理来源之间具有较大的相关性,分类结果基本与地理来源相一致,只有部分不同来源的材料聚到其他分类中。
本研究结果表明,50份黄淮海大豆种质资源中,两个居群(POP-Ⅰ、POP-Ⅱ)中86%的材料与UPGMA聚类和主成分分析(PCA)结果一致。其中仅石豆7号、南圣105这2份材料被聚类到黄淮海北部类群中,宁豆5号、冀豆12、邯豆5号、安07109、安豆5156这5份材料被聚类到黄淮海南部类群中,共有7份材料在黄淮海南、北部2个片区间互相交错。推测可能由于这些种质一定程度上与育种单位的遗传背景有所关联。比如:石豆7号的父母本(晋遗16/冀豆4号)均来自黄淮海北部,因此石豆7号被聚类到黄淮海北部类群中;冀豆12的母本为油83-14、邯豆5号的母本为徐8313均来自黄淮海南部,因此,冀豆12、邯豆5号被聚类到黄淮海南部类群中;宁豆5号更适于黄淮海灌溉区域种植,因此,其具有特殊的气候环境要求。
种质材料来源地或地理位置是影响种质材料遗传关系的重要因素,一般来自同一地区或地理生态区的种质材料具有相近的遗传信息,在聚类分析中就具有更为相近的遗传距离。本研究聚类结果也已经证实大豆居群的形成与其起源地的地理位置和气候环境有着密切的关系。
3.3 黄淮海不同居群种质资源遗传多样性比较
盖钧镒等[22]分析中国栽培大豆不同生态类型群体间遗传演化关系,表明中国栽培大豆不同生态类型群体间地理生态分化明显时,遗传距离较大。遗传分化系数(Gst)代表不同群体间的遗传分化水平,群体间遗传分化系数越大说明亲缘关系越远[23]。基因流(Nm*)是指生物个体从其发生地分散出去导致不同群体之间基因交流的过程,基因流的基本作用是削弱群体间的遗传差异[24]。
本研究可知,2个居群的总遗传多样度(Ht)为0.270 4,居群内遗传多样度(Hs)为0.259 1。居群间遗传分化系数(Gst)为0.141 7,基因流(Nm*)为3.03。通过对黄淮海大豆南部、北部两个群体的分化关系和相似系数研究发现,黄淮海两个大豆居群间差异较小,但居群内部个体差异较大。黄淮海两个大豆居群间存在中低度分化,居群间基因交流十分丰富。两个居群间遗传距离较小,相似系数较大。推测可能由于黄淮海南、北地区地理位置较近,气候环境相似,造成两者遗传基础极为相似。黄淮海北部大豆种质的有效等位基因数(Na)、Nei’s基因多样性指数(H)、Shannon’s信息指数(I)均大于黄淮海南部。黄淮海北部大豆种质资源的遗传多样性较黄淮海南部高,黄淮海北部大豆种质资源较为丰富。这与金尚昆等[8]认为黄淮海地区不同省区中,北京和河北材料多样性最高的结论一致。
推测可能由于山东、安徽、河南和江苏的部分地区大豆推广面积可观,黄淮海南部是大豆在黄淮海地区的主要种植区域[25]。因此,黄淮海北部的育种单位在新品种选育过程中会选择黄淮海南部种质资源作为亲本资源,进而培育出适于黄淮海南部推广的新品种;或者把在黄淮海北部表现晚熟的品种在黄淮海南部进行参试、审定并推广,无形中促进了黄淮海南、北两个居群之间和黄淮海南部居群内部种质资源的频繁交流。然而,如果对黄淮海地区南、北部种质资源中可作为优良亲本的种质材料进行重复利用,就会造成黄淮海地区大豆种质遗传背景趋于单一、遗传多样性指数有所降低和大豆生产推广风险逐渐升高。
4 结 论
本研究所用42对SSR引物具有丰富的多态信息。根据50份黄淮海大豆种质材料的遗传相似度聚类分析后,分为2个类群,UPGMA聚类结果与2个居群结构基本一致。居群间遗传距离较小,相似系数较大,存在中低度遗传分化。居群内部个体差异较大,居群之间存在丰富的基因交流。黄淮海北部大豆种质资源的遗传多样性较黄淮海南部高,黄淮海北部大豆种质资源较为丰富。
研究黄淮海大豆种质遗传多样性能够充分了解该地区种质资源的遗传背景,在育种过程中应尽量避免优良亲本的重复利用,减少相似遗传背景的干扰,拓宽遗传基础。应注重跨地区引种,合理利用不同时期育成的品种,有利于保持大豆的遗传多样性。
