棉花RCI2基因家族的鉴定及表达分析
2021-09-23刘楚楚吴玉香
刘楚楚,吴玉香
(山西农业大学 农学院,山西 太谷030801)
不同的非生物胁迫,如盐、旱、高温和低温,都会对植物的生长发育产生不利影响,因此了解植物抗逆遗传机制有助于提高植物的抗逆能力,从而提高作物产量。非生物胁迫通过损害细胞膜通透性破坏离子稳态,诱导电解质渗漏[1]。在酵母中,Pmp3基因的缺失会导致酵母质膜电位的增加,提高酵母细胞对阳离子的敏感性。而植物中RCI2基因在酵母中的表达可以补偿Pmp3基因缺失带来的影响,因此,植物中RCI2蛋白在细胞离子稳态和细胞膜电位的维持中具有积极的作用[2]。
稀有冷诱导2(RCI2)基因在大多数植物有不同的名字,且都受到非生物胁迫的显著诱导[3]。在拟南芥中,该基因被称为稀有冷诱导2(RCI2)[4],AtRCI2A和AtRCI2B的表达由低温、脱落酸、干旱和NaCl诱导,而经过不同胁迫处理后,除基因AtRCI2C的表达水平与对照无显著差异外,AtRCI2D-F对这些处理存在不同程度的响应[5];在高耐盐性的长穗偃麦草中,首次发现早期盐诱导3(Esi3)基因,该物种是小麦的野生近缘物种,通过对小麦中Esi3基因同源序列的鉴定以及对这些基因的差异表达模式的分析发现,该基因的表达受到高温以及渗透胁迫的诱导[6];在水稻中该基因被称为低温诱导6(Lti6),OsLti6b的过表达可以增强水稻的耐寒性[7];在玉米中,ZmRCI2在干旱胁迫响应中起重要作用[8];在星星草中,该基因被称为质膜蛋白3(PutPMP3),低温、盐胁迫、盐碱胁迫、外源ABA和干旱均可诱导PutPMP3-1和PutPMP3-2在叶片和根系中表达[9];在油茶中,Cs-RCI2A和CsRCI2E都受到盐和低温的诱导,Cs-RCI2E还会受干旱胁迫的诱导[10]。
RCI2基因,包含2个外显子和一个内含子,编码小分子量高疏水性蛋白,仅有PMP3一个高度保守的结构域(PF01679)[3,5,8,11]。RCI2蛋白具有2个跨膜结构域,根据报道,亚细胞定位在质膜、高尔基体、内质网、胞内小泡等含有膜的结构上[3]。该蛋白根据氨基酸长度可分为2类。I类RCI2s编码较短的蛋白质(约52~64 aa),根据其C端亲水性进一步分为2个亚组(Ia和Ib):Ia组RCI2s具有疏水性C端,而Ib组RCI2s具有亲水性C端[1,3]。而II类RCI2s编码较长的蛋白质(约60~89 aa),具有疏水性C末端,并且富含带电残基[5]。
棉花在我国国民经济中占有重要地位,新疆棉区作为我国的棉花主要种植区,自然降水量不足,蒸腾作用强烈,土地盐渍化严重。因此解析棉花抗逆分子机制、培育高产抗逆新品种具有重要意义。RCI2基因在植物响应逆境的过程中具有重要作用,但至今还没有对棉花中RCI2基因家族进行研究。本文主要对雷蒙德氏棉、亚洲棉、陆地棉和海岛棉中的RCI2基因进行鉴定,分析了它们在染色体上的分布及在不同棉种中的系统发育关系;对陆地棉中RCI2基因启动子区的顺式元件、组织器官表达模式以及非生物胁迫下的表达模式进行研究,有助于了解RCI2基因在棉花抗逆分子机制中的作用,为进一步对棉花的遗传改良提供理论依据。
1 材料和方法
1.1 棉花RCI2基因家族的全基因组鉴定
从Phytozome(https://phytozome.jgi.doe.gov/)网站上下载拟南芥RCI2基因家族的氨基酸序列,以fasta格式保存。从Cottongen(https://www.cottongen.org/)网站上下载陆地棉、海岛棉、亚洲棉和雷蒙德氏棉的基因组序列和注释数据。依据拟南芥RCI2家族蛋白序列,利用TBtools软 件[12]中BLAST GUI Wrapper-Two Sequences Files工具(E=1e-5),搜寻上述4个棉种中的RCI2家族蛋白序列。通过在线工具NCBI CD-Search(https://www.ncbi.nlm.nih.gov/cdd/)和Search Pfa(http://pfam.xfam.org/search)对结构域进行验证,最终确定棉花RCI2家族成员并得到其序列。借助在线工具ExPASy(https://www.expasy.org/)计算陆地棉RCI2家族成员的分子量和等电点。利用亚细胞定位工具Cell-Ploc2.0(http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/)预测陆地棉RCI2家族的亚细胞定位。
1.2 雷蒙德氏棉RCI2基因染色体结构分析与命名
利用TBtools[12]软件将雷蒙德氏棉中的RCI2基因进行染色体结构定位,并按照该基因在染色体上的位置进行命名,将基因位置用可视化工具(Gene Location Visualize),绘制雷蒙德氏棉RCI2基因家族成员的染色体分布图。
1.3 棉花RCI2基因家族的进化分析
利用MEGA-7[13]内置的Clustal W对拟南芥和4个棉种的RCI2蛋白序列进行比对,采用UPGMA法[14],Bootstrap重复1000次构建进化树。根据进化树对亚洲棉、海岛棉和陆地棉进行命名。利 用 在 线 工 具NCBI CD-Search(https://www.ncbi.nlm.nih.gov/cdd/)和MEME(http://memesuite.org/)分析RCI2蛋白序列的保守结构域,最大发现基序设置为15,其他参数为默认值。利用TBtools[12]将结果可视化。
1.4 陆地棉RCI2基因启动子顺式元件预测
使用TBtools[12]软件中的Gtf/Gff3序列提取(Gtf/Gff3 sequences Extract)工具,提取棉花RCI2基因家族成员CDS上游2000 bp的启动子序列,将提取到的序列提交至PlantCare(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)进行顺式元件预测。将结果进行筛选统计,并通过TBtools[12]软件中的热图工具(Heatmap)将结果可视化。
1.5 陆地棉RCI2基因表达模式
根据浙大已经发表的陆地棉TM-1组织器官表达转录组数据和不同非生物胁迫下的转录组数据(PRJNA 490626)进行分析,并用TBtools[12]软件中的热图工具(Heatmap)将结果可视化。
2 结果与分析
2.1 棉花RCI2基因家族的全基因组鉴定
根据拟南芥RCI2蛋白序列在棉花的不同数据库查找,最终在雷蒙德氏棉[15]中找到8个RCI2基因;在亚洲棉[16]中找到8个RCI2基因;在浙江大学发表的数据库[17]中,分别找到12个陆地棉和12个海岛棉RCI2基因;在华中农业大学发表的数据库[18]找到11个陆地棉RCI2基因和11个海岛棉RCI2基因;在得克萨斯大学奥斯汀分校发表的数据库[19]中找到14个陆地棉RCI2基因和18个海岛棉RCI2基因(表1)。本文主要对浙江大学测序数据库中的RCI2基因命名并分析。

表1 4个棉种不同数据库下RCI2基因家族成员的鉴定Table 1 Identification of RCI2 gene family members in different databases of four cotton species
由表2可见,陆地棉中RCI2基因都只包含2个外显子,氨基酸长度在57~88 aa之间,分子量介于6.27~9.56 kD,属于小分子量蛋白;预测等电点为4.23~9.11,除GH_A 07G1428和GH_D07G 1421外,其余基因的等电点都小于7.0,推测其为酸性蛋白。通过亚细胞定位预测发现,陆地棉中RCI2基因都定位在细胞膜上,另外还可能在叶绿体、细胞核、液泡等具有膜结构的细胞器中。根据报道,拟南芥中RCI2基因定位于细胞膜、高尔基体、内质网等结构,玉米、小麦中的该家族基因定位于细胞膜中[3]。

表2 陆地棉RCI2基因家族基本理化性质Table 2 Basic physical and chemical properties of RCI2 gene family in cotton
2.2 雷蒙德氏棉中RCI2基因染色体定位与命名
将雷蒙德氏棉中RCI2基因进行染色体结构定位,发现RCI2家族分布在D4、D5、D6、D7、D8、D12、D13号染色体。根据基因在染色体上的位置(图1),依次命名为Gr RCI2-1、Gr RCI2-2、Gr RCI2-3、Gr RCI2-4、Gr RCI2-5、Gr RCI2-6、Gr RCI2-7、Gr RCI2-8。

图1 雷蒙德氏棉RCI2基因在染色体上的分布Fig.1 Chromosomal location of RCI2 gene family in G.raimondii
2.3 棉花RCI2基因家族的进化分析
将拟南芥、雷蒙德氏棉、亚洲棉、陆地棉、海岛棉中的RCI2基因共同构建系统进化树。根据聚类分析以及雷蒙德氏棉中该基因的命名,将亚洲棉命名为GaRCI2-1~GaRCI2-8;将陆地棉命名为GhRCI2-2A、GhRCI2-2D、GhRCI2-3A、GhRCI2-3D、GhRCI2-4A、GhRCI2-4D、GhRCI2-5A、GhRCI2-5D、GhRCI2-7A、GhRCI2-7D、GhRCI2-8A、GhRCI2-8D;将海岛棉命名为GbRCI2-2A、GbRCI2-2D、GbRCI2-3A、GbRCI2-3D、GbRCI2-4A、GbRCI2-4D、GbRCI2-5A、GbRCI2-5D、GbRCI2-7A、GbRCI2-7D、GbRCI2-8A、GbRCI2-8D(图2)。
所有棉花RCI2基因都只有PMP3一个高度保守的结构域,普遍含有Motif 1和Motif 2(图3)。根据图2和图3,可以将棉花RCI2基因分为3类:第Ⅰ类,氨基酸链较短,只含有2~3个基序,有较短的C末端;第Ⅱ类,氨基酸链较长,含有4个保守基序,C末端含有Motif 3或Motif 5;前2类同拟南芥中RCI2同源性较高,第3类则与拟南芥同源性较低,在Motif 2前有较长的C链,该C链大部分为Motif 4。
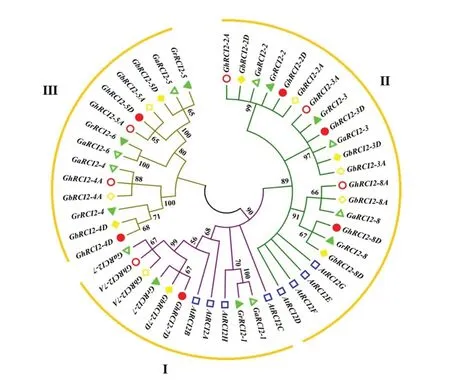
图2 基于拟南芥和四个棉种RCI2基因家族系统进化树Fig.2 Phylogenetic tree of RCI2 gene family based on Arabidopsis thaliana and four cotton species

图3 拟南芥与四个棉种中RCI2基因家族成员结构域分析Fig.3 Domain analysis of RCI2 gene family members in A.thaliana and four cotton species
2.4 陆地棉RCI2基因启动子顺式元件预测
通过对启动子顺式元件的分析,可以为基因的组织特异性表达或应激反应性表达模式提供思路和帮助。本研究利用在线PlantCARE软件对陆地棉中RCI2基因起始密码子上游2000 bp的基因组序列进行分析,并对主要的顺式元件分类统计。除常规顺式元件外,将剩余的顺式元件分为3类:生长发育相关的顺式元件、胁迫相关的顺式元件以及激素相关的顺式元件[11,20,21]。生长发育相关的顺式元件主要包括茎和根分生组织表达(CATbox)、胚乳表达(GCN 4-motif和AAGAA-motif、AACA-motif)、开花(MRE、CCAAT-box和ATrich-element)、玉米醇溶蛋白代谢(O2-site)、分生组织特异性激活(CCGTCC-box)共9个顺式元件,其中胚乳相关的顺式元件数目最多,其次是开花相关的顺式元件;胁迫相关的顺式元件主要包括无氧诱导(ARE)、干旱诱导(MBS)、低温(LTR)、应激(TC rich repeats,STRE)和创伤应激(WRE3,WUN-motif,W box)共8个,其中无氧诱导、创伤应激相关的顺式元件和STRE元件数目最多,其次为低温诱导相关的元件;激素相关的顺式元件包括脱落酸相关(ABRE)、茉莉酸甲酯相关(MYC和TGACG-motif)、乙烯相关(ERE)、水杨酸相关(TCA-element)、生长素相关(Aux RR-core,TGA-element)、赤霉素相关(GARE-motif,P-box)共9个,其中脱落酸、茉莉酸甲酯、乙烯相关的顺式元件数目最多(图4)。在这3类顺式元件中,除GhRCI2-8D无生长发育类相关元件外,每个基因至少含有1个生长发育相关的元件和多个胁迫和激素相关的元件,且激素相关元件数目最多,这表明RCI2基因的转录涉及复杂的网络调控机制。

图4 陆地棉中RCI2基因启动子顺式元件分析Fig.4 Cis-element analysis of RIC2 gene promoter in G.hirsutum
2.5 陆地棉RCI2基因表达模式分析
根据NCBI上浙江大学已经发表的组织器官表达转录组数据,将陆地棉TM-1中的12个RCI2基因按照log2(TPM+1)的值作图,发现该家族基因在不同器官的表达模式不同:GhRCI2-7基因在棉花中组成性表达;GhRCI2-8在胚3 DPA时特异性表达;GhRCI2-4D在棉花纤维中表达量较高,具有一定的组织特异性;GhRCI2-2和GhRCI2-3在根、茎、花中表达量较高;GhRCI2-5在整个生长发育期的表达量都比较低。在棉花的各个组织器官中,均有RCI2基因的表达,说明该家族基因参与多个组织器官发育的调控,影响整个植株的生长(图5)。

图5 陆地棉中RCI2基因组织器官表达分析Fig.5 Expression analysis of RCI2 gene in tissues in G.hirsutum
根据NCBI上浙江大学已经发表的非生物胁迫下TM-1转录组数据,将陆地棉中的12个RCI2基因按照log2[(stress-TPM+1)/(CK-TPM+1)]的值作图,发现该家族基因的表达都会受到盐、旱、高温、低温的诱导,但在不同胁迫下的表达模式各不相同。GhRCI2-7在PEG、NaCl、低温、高温下都受到明显的诱导,与对照比较,表达量基本呈现先降低后升;GhRCI2-3受PEG和NaCl诱导较为明显,与对照相比,表现为先升高后降低的表达模式;GhRCI2-4受NaCl和高温诱导较为明显;GhRCI2-5在不同胁迫下表达模式整体呈现升高的趋势;GhRCI2-2在不同胁迫下呈现先升高后降低的表达趋势(图6)。

图6 陆地棉中RCI2基因在不同胁迫下表达模式分析Fig.6 Expression pattern of RCI2 gene in G.hirsutum under different stress conditions
3 讨论
RCI2的全基因组鉴定及分析已经在拟南芥、水稻[5]、玉米[8]、黄瓜[11]等植物中完成。本研究利用生物信息学技术分别从雷蒙德氏棉、亚洲棉、陆地棉与海岛棉中分别鉴定出了8、8、12、12个RCI2基因。目前主要的栽培棉种陆地棉[G.hirsutum,(AD)1]和海岛棉[G.barbadense,(AD)2]是异源四倍体棉种,它们的纤维产量占全世界总产量的98%以上[22]。异源多倍体棉种由A基因组和D基因组的二倍体种杂交加倍形成,是棉属植物跨洋传播的一个偶然结果。雷蒙德氏棉(G.raimondii)被认为是与异源多倍体棉花实际D基因组亲本最接近的现存种;而草棉(G.herbaceum)与亚洲棉(G.arboreum),都与异源多倍体的实际A基因组亲本具有同等程度的差异[23]。多倍体化是植物进化及物种形成的重要推动力,但是基因组的加倍可能导致基因功能的冗余,从而使基因出现功能分化、功能丧失和亚功能化[22]。但是,RCI2基因在棉花的进化过程中高度保守,四倍体棉种中A、D两个亚组的基因可以分别和二倍体棉种亚洲棉、雷蒙德氏棉中的基因相互对应。虽然在浙江大学测序数据库中没有找到Gr RCI2-1与Gr RCI2-6相对应的陆地棉与海岛棉RCI2基因,但在华中农业大学、得克萨斯大学奥斯汀分校发表的数据库种可以找到(表1),可能是由于RCI2基因片段较小(表2),在测序过程中存在误差。
除了细胞膜外,RCI2基因可以在不同质膜上发挥作用,如AtRCI2D定位于内质网和高尔基体上[5]。据研究报道,干旱胁迫会引起活性氧的产生,损伤类囊体膜[24];盐胁迫会导致叶绿素降解[25];热胁迫会改变类囊体结构,降低光系统的活性[26],这些都会抑制植物的光合作用[27]。当植物的根所在的外部环境中盐浓度过高时,植物会吸收盐和营养物质,通过共质体将其转移到根的中心,再穿过木质部的薄壁细胞到达维管细胞中,因此共质体是植物控制盐的一个重要途径,在这个过程中液泡发挥了重要作用[28]。在本研究中,预测GhRCI2-2定位于叶绿体上,GhRCI2-5和GhRCI2-8定位于细胞核上,GhRCI2-7定位于液泡上,这表明棉花中不同的RCI2基因通过不同的机制应对非生物胁迫。在胁迫诱导表达分析中,GhRCI2-7基因受到低温、高温、干旱和盐的诱导(图6),且GhRCI2-7基因与拟南芥AtRCI2A、AtRCI2B同源性为89%,根据报道AtRCI2A、AtRCI2B受到低温、干旱、盐及外源ABA的显著诱导[5],说明它们功能的相似性。ABRE是胁迫诱导的ABA基因启动子上的顺式元件,ABA通过产生ROS诱导第二信使激活防御反应。抗氧化剂酶基因和非酶防御系统基因的表达也被ABA信号诱导机制激活[29]。这表明,棉花可能通过产生ABA诱导RCI2基因的表达来应对非生物胁迫。
陆地棉GhRCI2-7基因在棉花中组成型表达,同时也明显受到多种非生物胁迫的诱导。因此,该基因可以作为目标基因来对RCI2基因家族进行下一步的功能研究,从新的角度解析棉花抗逆通路。
4 结论
本文主要鉴定了棉花中RCI2基因家族成员,根据染色体定位和亲缘关系分析对其进行了分类和命名,通过分析家族基因的启动子顺式元件和诱导表达模式发现,陆地棉中所有RCI2基因都有胁迫响应相关元件,并对低温、高温、干旱和盐等非生物胁迫有显著反应,为进一步解析该基因家族的功能奠定了理论基础。
