干预HSF1影响MDR1的表达逆转结直肠癌HCT-8细胞株耐药的实验分析
2021-09-22郭宁杰李扬扬贾真真崔忠泽路丽祯吴淑华
郭宁杰,李扬扬,贾真真,崔忠泽,路丽祯,吴淑华
目前,化疗仍然是肿瘤患者术后辅助治疗或姑息治疗的主要方式之一,然而多药耐药性的产生严重影响患者生存期。肿瘤产生耐药性的因素众多,其中最经典的分子机制是7号染色体上多药耐药基因MDR1的过表达,其编码的跨膜糖蛋白(P-gp)可将抗肿瘤药物泵出细胞外,使细胞内药物浓度降低,进而介导肿瘤多药耐药,严重影响临床化疗效果。因此,抑制MDR1蛋白的过表达,有可能成为逆转耐药的新途径。新近研究发现,热休克转录因子1(heat shock factor 1, HSF1)在多种恶性肿瘤中高表达,可促进肿瘤细胞的存活和增殖[1-4]。HSF1可通过结合下游靶基因启动子区的热休克元件,进而促进下游基因的转录,并调控蛋白表达。近年,肿瘤中HSF1与非热休克蛋白表达的关系受到关注[5]。本课题组前期实验结果显示,结直肠癌中HSF1和MDR1具有相关性[6]。本实验采用RNA干扰(RNA interference, RNAi)技术,通过抑制或过表达HSF1,观察HSF1表达差异对HCT-8/FU细胞株增殖、克隆、侵袭能力及常用化疗药物5-FU、伊立替康耐药性的影响,为抑制结直肠癌的发生、发展并逆转耐药提供新思路。
1 材料与方法
1.1 材料人回盲部结肠腺癌细胞株HCT-8及其耐药细胞株HCT-8/FU购自中国科学院细胞库;胎牛血清和PRMI 1640培养基购自美国Gibco公司;靶向HSF1基因的特异性慢病毒序列、无义序列组慢病毒及转染增强试剂,均由上海吉凯公司设计提供;HSF1、MDR1、Cyclin D1、MMP-2与BCL-2抗体,均购自美国Abcam公司;辣根过氧化物酶标记的羊抗兔多克隆IgG抗体II抗购自福州迈新公司;CCK-8购自日本同仁研究所;吉姆萨染液购自北京索莱宝公司;Transwell双层细胞培养板购自美国Corning公司;Western blot实验中凝胶试剂盒、RIPA细胞裂解液等试剂盒,均购自江苏碧云天公司;RT-PCR实验中RNA提取试剂盒、逆转录试剂盒和PCR试剂盒等,均购自日本Takara公司;HSF1及MDR1、GAPDH特异性引物序列由Takara公司合成。
1.2 方法
1.2.1细胞培养 人回盲部结肠腺癌细胞株HCT-8及其耐药细胞株HCT-8/FU在RPMI 1640培养基和5%胎牛血清培养基中于37 ℃ 5% CO2的培养箱中培养。将HCT-8/FU细胞接种于含有5 μmol/L的5-FU培养基中培养1周,以维持其耐药性表型,每隔2天换液1次,每3天用0.25%的胰蛋白酶消化传代,取对数生长期的细胞进行后续实验。
1.2.2细胞转染 设计并合成2条靶向HSF1基因特异性序列,其中1条为抑制HSF1的序列,命名为HSF1-RNAi,另一条为过表达HSF1的序列,命名为LV-HSF1,同时设计1条无义干扰组序列,命名为无义序列组,转染实验参照RNAi-mate产品说明书进行操作,优化转染条件。转染前一天将HCT-8/FU细胞接种于6孔板中,共设置4组,分别为空白对照组(未行任何干预的HCT-8/FU细胞)、无义序列组(转染无义序列组慢病毒的HCT-8/FU细胞)、HSF1-RNAi组(转染抑制HSF1基因组慢病毒的HCT-8/FU细胞)、LV-HSF1组(转染过表达HSF1基因组慢病毒的HCT-8/FU细胞)。利用Hitrans G感染增强液辅助转染,将配比好的siRNA和增强液及培养基混匀后加入6孔板中,使试剂均匀覆盖细胞。8 h后更换新鲜培养基,继续培养细胞48 h观察荧光的表达。连续培养72 h后将转染的细胞在8 μg/mL的嘌呤霉素中培养4周,选择并扩增具有嘌呤霉素抗性和荧光标记的细胞,细胞经RT-PCR检测确定转染成功后,用于后续实验。
1.2.3RT-PCR法 使用TB Green嵌合荧光染料法进行相对定量PCR检测。HSF1、MDR1、GAPDH的转录物通过逆转录聚合酶链反应检测4个细胞组分离的总核糖核酸的相对定量。使用RNA提取试剂盒提取待测细胞的总RNA,利用逆转录试剂盒合成cDNA链,以cDNA为模板加入引物(表1),使用CFX96逆转录聚合酶链反应检测系统进行聚合酶链反应扩增。反应条件:95 ℃ 30 s、GOTO 95 ℃ 5 s、60 ℃ 30 s,合计40个循环。RT-PCR结果由系统软件自动分析而成,采用ΔΔCT法,各组样品的相对表达量=2-ΔΔCT,ΔΔCT=实验组ΔCT-空白对照组ΔCT,ΔCT=目的基因CT值-GAPDH CT值。实验重复3次,结果取平均值。
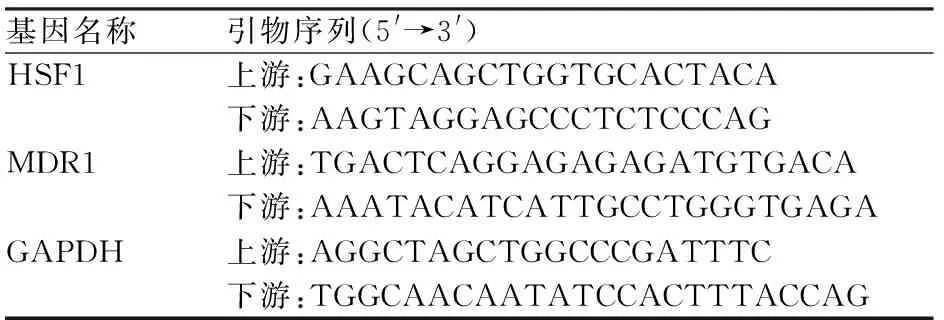
表1 RT-PCR引物序列
1.2.4Western blot法 收集各组细胞,并将细胞在含有新添加的蛋白酶抑制剂PMSF的RIPA裂解缓冲液中裂解。将蛋白质裂解物与5×上样缓冲液按比例混合后,沸水浴煮样5 min,取10 μL蛋白加载到聚丙烯酰胺凝胶上,用SDS-PAGE分离,电泳转移到PVDF膜上。在室温下用5%奶粉在TBST缓冲液中置于摇床上封闭1.5 h,加入一抗HSF1(稀释度1 ∶50 000)、P-gp(稀释度1 ∶2 000)、Cyclin D1(稀释度1 ∶10 000)、BCL-2(稀释度1 ∶2 000)、MMP-2(稀释度1 ∶3 000)及β-actin(稀释度 1 ∶3 000),4 ℃孵育过夜。TBST洗膜,与相应的辣根过氧化物酶标记的二抗在室温下孵育1 h(稀释度1 ∶10 000),TBST洗膜,使用凝胶成像系统ECL化学发光试剂盒显影,曝光成像。Image J软件扫描灰度值,以β-actin内参灰度值为对照,目的基因蛋白相对表达量=目的基因灰度值/β-actin灰度值×100%,用其比值作图,实验重复3次。
1.2.5CCK-8实验 细胞增殖测定使用细胞计数试剂盒CCK-8。取未转染的细胞为空白对照组及实验组(无义序列组、HSF1-RNAi组、LV-HSF1组)接种细胞于96孔板中,每组设5个复孔。在未接种细胞的孔中加入不含血清的培养基作为调零孔,培养24 h后吸去培养基,并将完整培养基(200 μL)添加到每个孔中(伊立替康浓度分别为10、50、100、150、200 μmol/L;5-FU浓度分别为5、10、50、100和200 μmol/L)。48 h后,向每孔中加入10 μL CCK-8溶液,37 ℃孵育2 h。使用酶标仪测定450 nm处每个孔的吸光度值(OD),实验进行3次。根据OD值计算细胞存活率,细胞存活率=OD实验组/OD对照组×100%。根据细胞活力和药物浓度绘制The dose反应曲线。
1.2.6细胞侵袭试验 将Matrigel基质胶按1 ∶6与RPMI 1640培养基混合,取60 μL铺入Transwell室的上部,37 ℃孵育3 h。在24孔板内每孔加入600 μL含有20%的FBS RPMI 1640培养基,将小室置于孔内,调整细胞数至每毫升1×105个,取100 μL细胞溶液加入Transwell小室,封闭24 h。用棉签将膜顶部的非迁移性细胞清除,甲醇固定,吉姆萨染色,倒置显微镜下观察。随机分析每个腔室的3个视野,计算每个视野的平均穿膜细胞数,实验进行3次。
1.2.7平板克隆实验 分别取对数生长期每孔1×103个细胞接种于6孔板中,并加入2 mL 10% FBS RPMI 1640。细胞在37 ℃ 5%CO2中孵育1~2周。当培养皿中出现肉眼可见的细胞克隆球时,培养终止。采用吉姆萨染色,在100×视野下计数并拍照。分析每个培养皿中的3个随机区域,实验重复3次。

2 结果
2.1 结直肠癌耐药细胞株中HSF1、MDR1的表达采用Western blot法检测HCT-8/FU细胞与其亲本细胞系(HCT-8)中HSF1、MDR1的表达。结果显示,HCT-8/FU细胞中HSF1蛋白表达明显高于HCT-8细胞株;与HCT-8细胞株相比,HCT-8/FU细胞中MDR1的蛋白表达水平明显升高,差异有统计学意义(P<0.01,图1)。
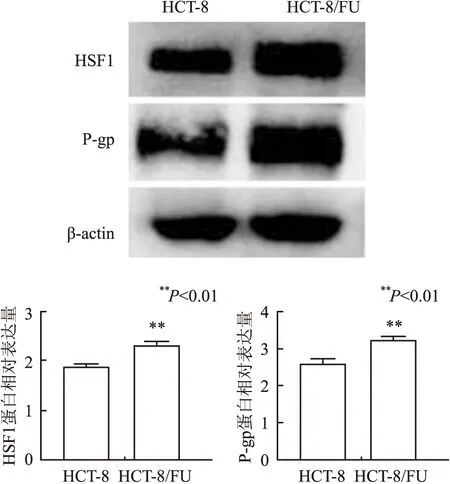
图1 Western blot法检测HCT-8及HCT-8/FU细胞株中HSF1、MDR1(P-gp)的表达
2.2 各组中HSF1 mRNA、蛋白的表达通过携带特定上调或下调HSF1序列的慢病毒分别转染HCT-8/FU,嘌呤霉素筛选以获得稳定敲除HSF1及过表达HSF1细胞株,以转染无义序列组的稳定细胞株作为无义序列组。通过倒置荧光显微镜观察各组细胞株中慢病毒的荧光蛋白表达,各组间的细胞形态无明显变化(图2)。
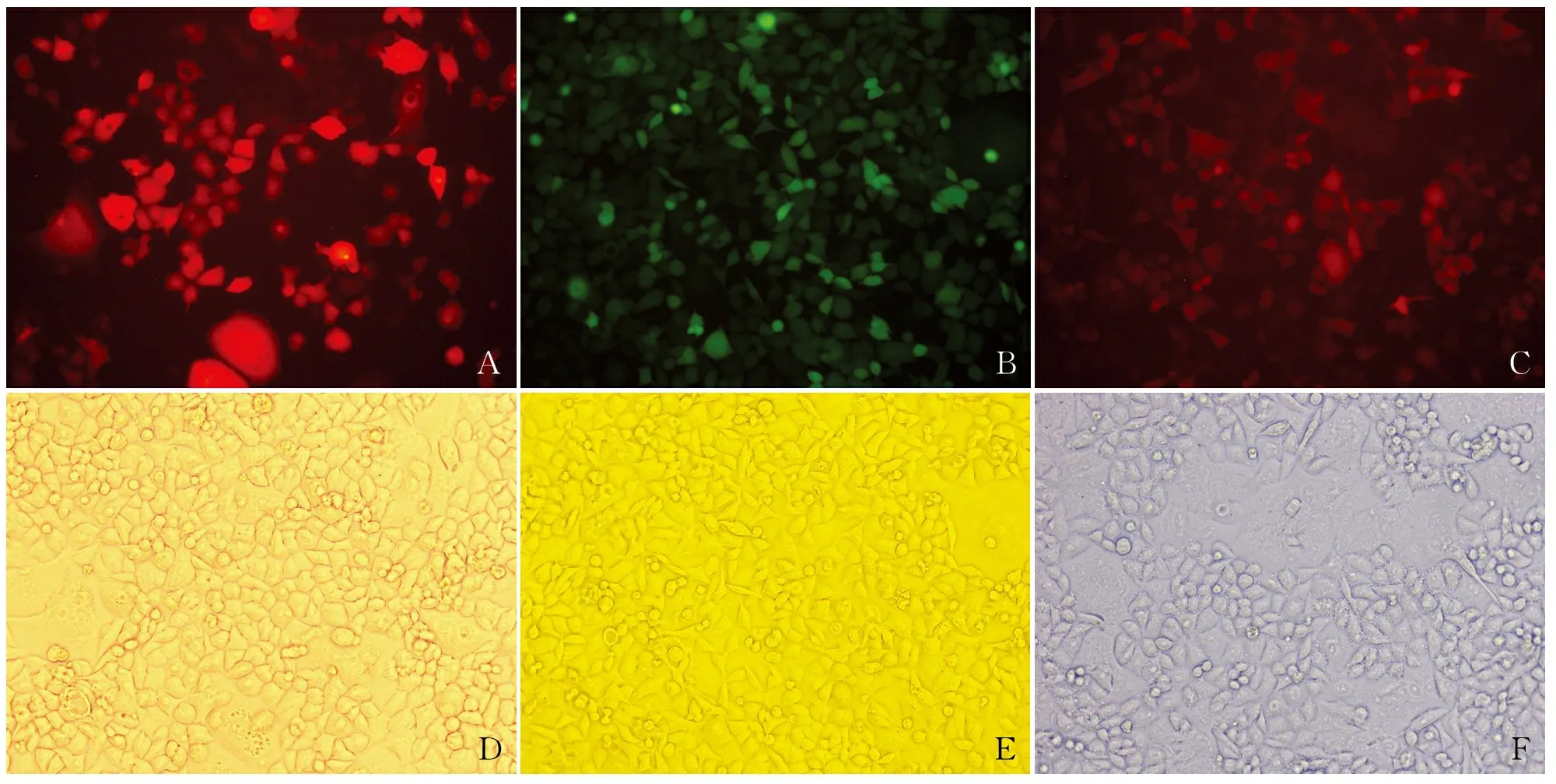
图2 各组转染慢病毒的荧光表达:A.无义序列组荧光图;B.HSF1-RNAi组荧光图;C.LV-HSF1组荧光图;D.无义序列组白光图;E.HSF1-RNAi组白光图;F.LV-HSF1组白光图
本实验结果显示,HSF1-RNAi组中HSF1 mRNA及蛋白表达水平明显低于空白对照组及无义序列组,而LV-HSF1组中HSF1的mRNA、蛋白表达水平明显高于空白对照组及无义序列组,差异有统计学意义(P<0.01,图3)。无义序列组中HSF1的mRNA、蛋白水平与空白对照组相比,差异无统计学意义(P>0.05,图3);提示已获得稳定敲除HSF1及过表达HSF1细胞株。
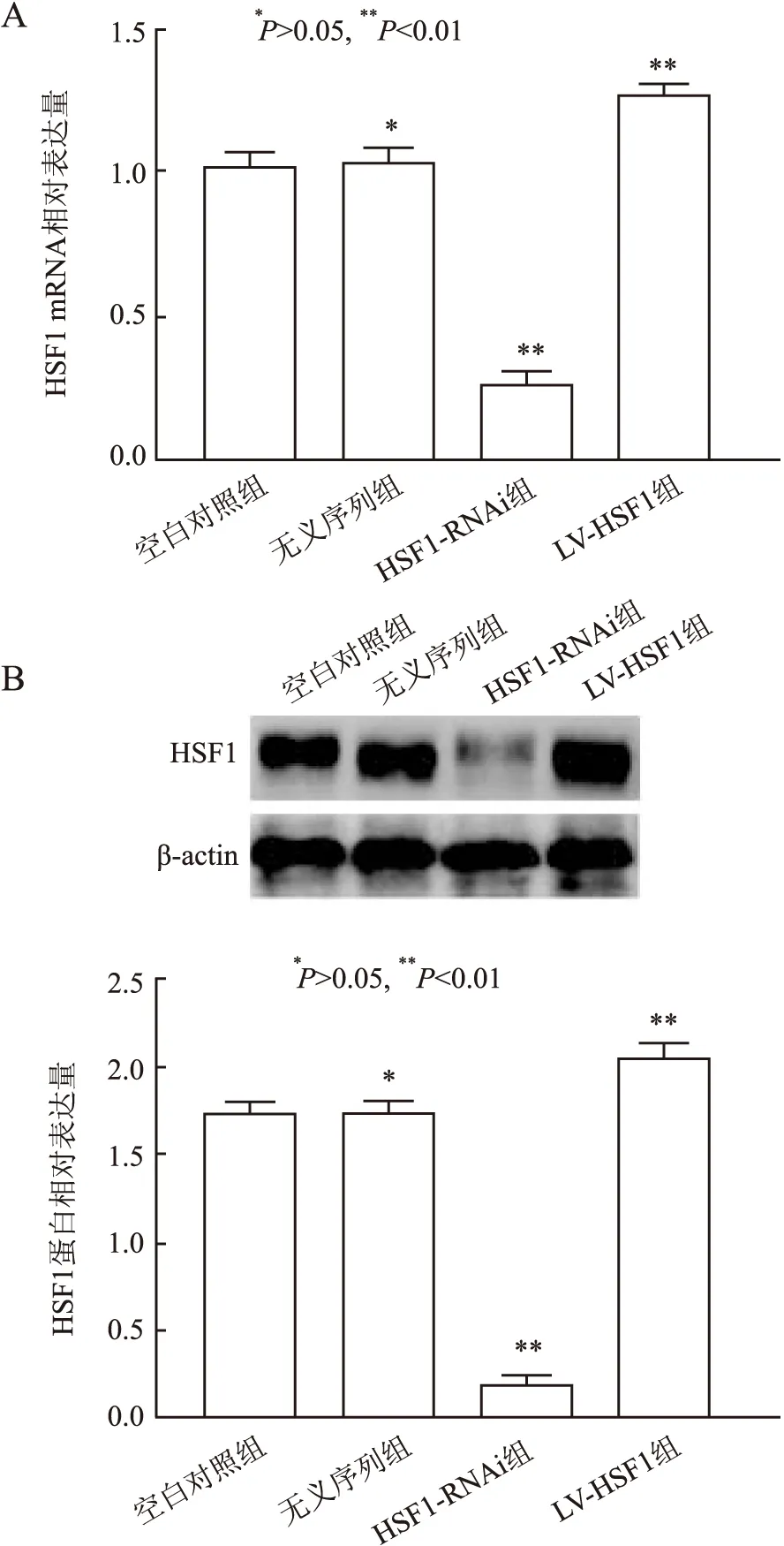
图3 各组中HSF1 mRNA及蛋白的表达:A.HSF1 mRNA的表达量;B.HSF1蛋白的表达量及柱状统计图
2.3 不同HSF1表达对MDR1 mRNA、蛋白的影响实验结果显示,HSF1-RNAi组中MDR1 mRNA及蛋白表达水平明显低于空白对照组及无义序列组,而LV-HSF1组中MDR1的mRNA及蛋白表达水平明显高于空白对照组及无义序列组,差异有统计学意义(P<0.01,图4)。无义序列组中MDR1的mRNA及蛋白水平与空白对照组相比,差异无统计学意义(P>0.05,图4)。
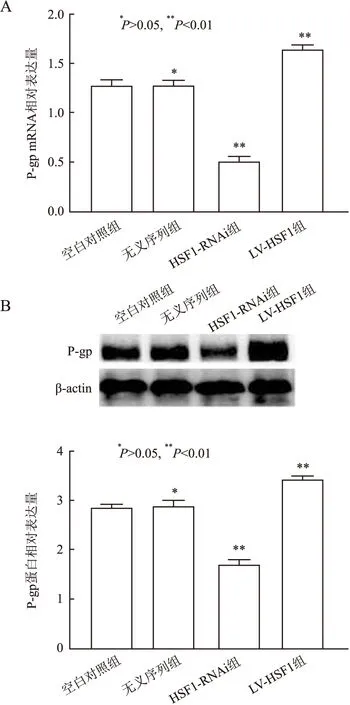
图4 各组中MDR1的mRNA及蛋白表达: A.P-gp mRNA的表达量;B.P-gp蛋白的表达量及柱状统计图
2.4 不同HSF1表达对Cyclin D1、MMP-2与BCL-2的影响实验结果显示,HSF1-RNAi组中Cyclin D1、MMP-2与BCL-2的蛋白表达水平明显低于空白对照组及无义序列组,而LV-HSF1组中Cyclin D1、MMP-2与BCL-2的蛋白表达水平明显高于空白对照组及无义序列组,差异有统计学意义(P<0.01,图5),而Cyclin D1、MMP-2与BCL-2蛋白在空白对照组与无义序列组中的表达差异无显著性(P>0.05,图5)。
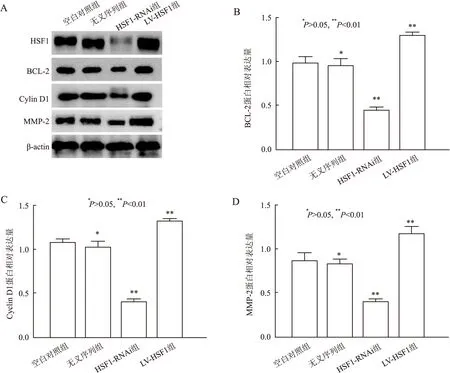
图5 Western blot法检测不同因子在各组间的表达量:A.Western blot法检测各因子蛋白表达量;B.BCL-2蛋白表达柱状统计图;C.Cyclin D1蛋白表达柱状统计图;D.MMP-2蛋白表达柱状统计图
2.5 HSF1表达差异对细胞株的克隆能力及侵袭能力的影响平板克隆实验结果显示,HSF1-RNAi组与空白对照组比较,其HCT-8/FU细胞的群体依赖性及增殖能力明显下降,而LV-HSF1组中细胞表现出群体依赖性和增殖能力的增强,差异有统计学意义(P<0.01)。无义序列组与空白对照组增殖能力差异无显著性(P>0.05,图6A)。
Transwell实验结果显示,HSF1-RNAi组与空白对照组相比,其穿过具有基质凝胶透孔的细胞数量显著减少,而LV-HSF1组穿孔细胞明显增加,差异有统计学意义(P<0.01),无义序列组与空白对照组差异无统计学意义(P>0.05,图6B)。
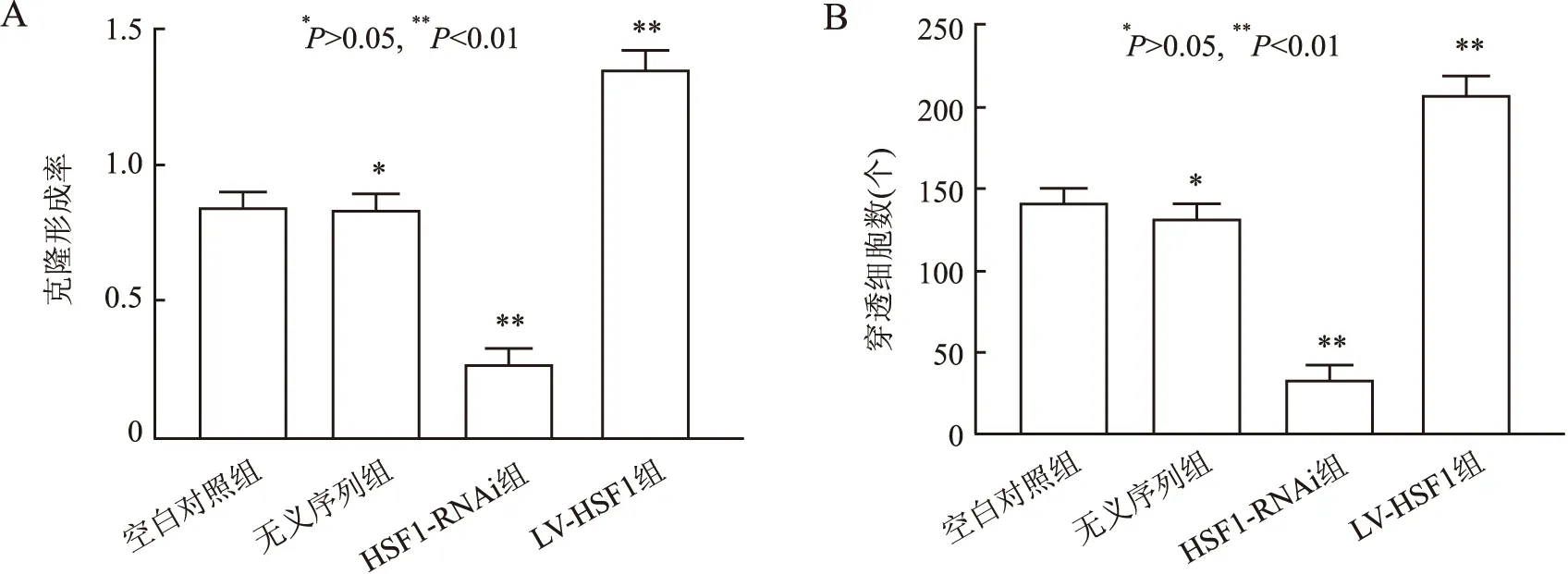
图6 平板克隆实验(A)及Transwell实验(B)检测HSF1的表达差异对细胞株的克隆及侵袭能力的影响
2.6 HSF1表达差异细胞株对伊立替康和5-FU化学的敏感性采用CCK-8实验检测各组细胞株的增殖活性,观察不同HSF1表达细胞株对不同浓度的伊立替康和5-FU的化学敏感性。结果显示,与空白对照组相比,HSF1-RNAi组分别对伊立替康和5-FU的化学敏感性呈浓度依赖性增强,细胞活性明显降低;而LV-HSF1组细胞则表现对伊立替康和5-FU的耐药性增强,细胞活性明显升高,差异有统计学意义(P<0.05,图7),空白对照组与无义序列组细胞对药物敏感性差异无显著性(P>0.05,图7)。
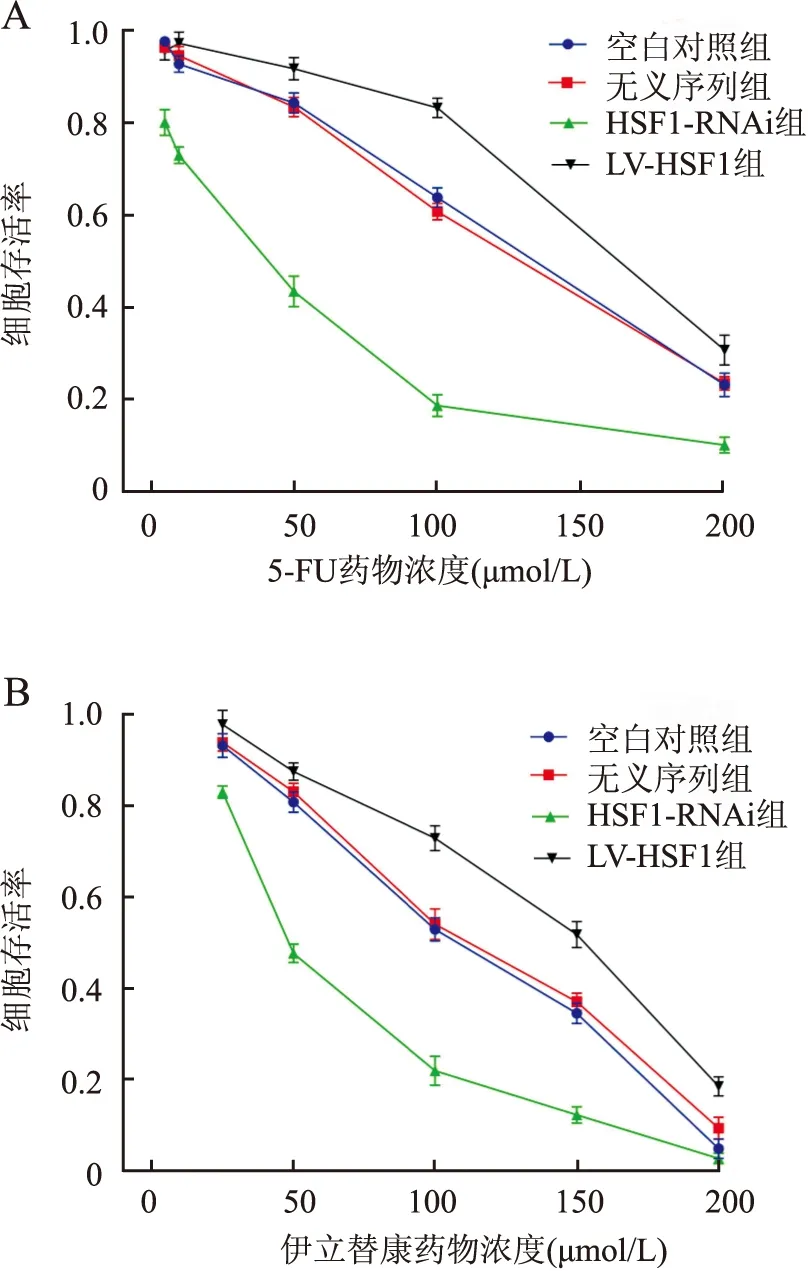
图7 各组细胞株对5-FU及伊立替康的敏感性:A.5-FU剂量反应曲线;B.伊立替康剂量反应曲线
3 讨论
HSF1是真核生物热休克反应的主要调节因子。HSF1以未活化的单体形式存在于细胞质中,当受到外界刺激时,如生物、化学(化疗药物)、物理(热应激)等有害条件的刺激时,活化为三聚体进入细胞核,进入胞核的HSF1可结合下游目的基因,从而调控相关基因的表达[7]。HSF1活化是瞬时的,并随着应激缓和而减弱。但肿瘤细胞中HSF1通常表现出持续活化状态,即HSF1在长期蛋白毒性压力下,其介导的蛋白毒性应激反应可维持肿瘤细胞稳态,促进肿瘤细胞存活[8]。新近研究显示,多种类型的人类恶性肿瘤中HSF1均呈高表达,其可形成一种肿瘤细胞保护性机制以促进细胞存活增殖和侵袭转移,从而影响肿瘤的发生、发展[9-15]。HSF1促癌作用是复杂的,其可以参与多种信号通路诱导癌细胞的增殖。研究发现,胆囊癌中活化的HSF1信号增加TGF-β1的表达和旁分泌信号,诱导癌旁成纤维细胞向肿瘤相关成纤维细胞的转分化,导致癌旁成纤维细胞向胆囊癌募集,并增加肿瘤相关成纤维细胞中血小板反应蛋白的表达,从而促进胆囊癌的发生、发展[16]。研究表明,HSF1通过调节干细胞标记SOX2的表达,激活MMP-2,增强癌干细胞特性[17]。同时HSF1可抑制E-cadherin的表达,促进β-catenin及MMP-2的表达,从而促进肿瘤的侵袭和转移。本课题组前期实验结果显示,HSF1在结直肠癌中表达增高,并与结直肠癌的发生、发展及预后密切相关[18],但其分子机制尚不明确。本实验通过体外细胞培养,采用RNAi技术观察HSF1对结直肠癌细胞的影响。实验结果显示,LV-HSF1细胞株中细胞周期素Cyclin D1与抗细胞凋亡因子BCL-2的表达均升高,并且与肿瘤细胞侵袭相关的因子MMP-2表达亦同时升高;细胞生物学行为显示肿瘤细胞的侵袭、转移能力及细胞群体依赖性、增殖能力均明显增强;而HSF1-RNAi细胞株中则表现为Cyclin D1、MMP-2与BCL-2的表达降低,肿瘤细胞的侵袭转移及增殖能力下降。即高表达HSF1的结直肠癌细胞具有更强的增殖、侵袭及转移能力。实验推测,在肿瘤的发生、发展中HSF1作为转录因子,可不依赖热休克反应,而启动调控下游多种靶基因,促进肿瘤增殖、抗凋亡,进而发生侵袭及转移。HSF1在多种信号通路中的交互作用使其成为癌症发展过程中至关重要的分子,有可能成为潜在的靶点。
在热休克反应中,活化后的HSF1以三聚体形式进入胞核,可特异性结合下游靶基因启动子区5′端的热休克元件(heat shock element, HSE),启动靶基因的转录翻译,从而调控目的基因的表达[19]。有研究发现,HSF1不仅可与热休克蛋白基因启动子区的HSE结合,启动热休克蛋白的转录,同时也可启动非热休克蛋白家族基因的转录[20-21]。HSF1可与Snail的热休克元件区结合以促进其表达,进而通过调控上皮-间质细胞转化促进迁移[22]。新近研究显示[23],多药耐药基因MDR1基因启动子区域中存在热休克元件。在小鼠的心肌细胞和肝细胞中,沉默HSF1可增强MDR1蛋白(P-gp)的表达[24]。在小鼠和人类肺癌A549细胞以及人类乳腺癌T47D细胞、MCF7细胞中,MDR1的表达与HSF1的表达呈正相关[25]。由此可见,HSF1与MDR1的表达有关,但两者的调控关系尚存在争议。本课题组前期实验结果显示,结直肠癌中HSF1高表达组中MDR1的表达明显高于HSF1阴性组,两者呈正相关,但未对其调控机制进行深入分析。本实验结果显示,通过沉默结直肠癌耐药细胞株中的HSF1,发现MDR1的mRNA及蛋白水平均明显下降,而上调HSF1后,MDR1的表达明显增高;提示HSF1可调控MDR1的转录及翻译。作者认为HSF1有可能是通过与MDR1启动子区的HSE结合,启动MDR1的转录,进而调控MDR1蛋白的表达,诱导肿瘤耐药。
研究发现,HSF1与乳腺癌细胞的雌激素抵抗有关,HSF1的过表达导致了雌激素α降解,雌激素受体α激活基因表达减少,降低HSF1水平则恢复雌激素受体α的表达并恢复对抗雌激素的反应[26]。在结直肠癌中,有关HSF1诱导5-FU和伊立替康耐药的发生机制鲜见报道。本实验结果显示,人结肠腺癌5-FU耐药细胞株中HSF1 mRNA及蛋白表达水平均明显高于亲本细胞株。应用RNAi技术后,同一药物浓度梯度下,沉默HSF1的肿瘤细胞对伊立替康和5-FU的化学敏感性增强,而过表达HSF1的细胞对伊立替康和5-FU耐药性明显增强。我们认为过表达HSF1后可增强MDR1转录及翻译,以促进MDR1蛋白(P-gp)的表达,进而使肿瘤细胞将药物逆浓度梯度转运出细胞外,使肿瘤细胞内活性药物减少,减少了对肿瘤的杀伤作用,并介导肿瘤的多药耐药性。同时,HSF1过表达导致的肿瘤细胞增殖活性、抗凋亡能力及侵袭能力的增强,也在一定程度上增加了肿瘤细胞对化疗药物的抵抗力。
综上所述,HSF1不但可增强肿瘤细胞克隆及侵袭能力,促进肿瘤细胞增殖,同时可启动MDR1转录,导致MDR1蛋白(P-gp)高表达,从而增加化疗药物的外排,削弱化疗药物的杀伤作用,导致肿瘤细胞耐药。本实验仅通过RNAi技术对HSF1与MDR1的相互作用及其分子机制进行初步分析,两者是否可通过HSE结合而直接作用,并发挥生物学功能尚未验证,以及体外实验中HSF1表达的改变能否影响肿瘤的致瘤性,仍有待进一步分析。因此,深入探讨HSF1及MDR1相互作用的分子机制,对寻找新的个性化分子靶向治疗方案、抑制肿瘤生长及逆转肿瘤耐药具有重要意义。
