基于SRAP标记半枫荷天然种群的遗传多样性分析
2021-09-16叶兴状张明珠刘益鹏文国卫赖文峰范辉华张国防
叶兴状, 张明珠, 刘益鹏, 文国卫, 赖文峰, 范辉华, 张国防, 刘 宝,①
(1. 福建农林大学林学院, 福建 福州 350002; 2. 福建省林业科学研究院特色资源研究所, 福建 福州 350012)
由于人类活动对生态环境的破坏日益严重[1],加之全球气候变暖[2],许多植物的分布区域锐减,由连续分布演变成间断分布,进而成为濒危植物[3]。因濒危植物自身适应水平较低,加之繁殖障碍和地理隔离,濒危植物间的基因交流越来越困难,有的甚至出现瓶颈效应,进而导致其遗传多样性丧失,甚至物种灭绝[4-5]。研究濒危植物的遗传多样性和遗传结构有助于濒危植物的保护与开发利用。
半枫荷(SemiliquidambarcathayensisChang)(2n=32)隶属于蕈树科(Altingiaceae),为稀有种、国家二级重点保护野生植物,零星散状分布于中国长江以南山区,具有重要的科研价值[6-7]。Wu等[7]利用分子标记证明半枫荷为枫香树属(LiquidambarLinn.)和蕈树属(AltingiaNoronha)的属间天然杂交种。半枫荷具有较高的观赏价值,是中国南方为数不多的彩叶树种,春、秋两季叶片部分变为紫红色、红色,且同株有2~5种叶型,具有良好的园林绿化应用前景。半枫荷还是药用植物,为传统苗药,其根具有祛风除湿、镇痛消肿和抑制病毒性肝炎抗原活性的功效[8]。由于半枫荷对生长环境要求苛刻,有效种子少,在群落内常为非优势种,在竞争光热资源时常处于弱势地位,再加上人们过度采挖半枫荷根等原因,导致其遗传资源大量流失,濒临灭绝[9]。目前,仅田晓明等[10]和叶兴状等[11]进行了半枫荷转录组测序等分子标记准备工作,亟需对半枫荷群体的遗传多样性和遗传结构进行研究,这对其遗传资源保护策略的制定具有重要意义。
相关序列扩增多态性(sequence-related amplified polymorphism,SRAP)标记是显性标记,具有简便、高效、重复性好等优点[12]。已有研究结果[12-14]显示:在研究对象基因型未知的情况下,采用SRAP标记技术研究其遗传多样性有效可行。Li等[14]利用SRAP标记对半枫荷近缘的金缕梅科(Hamamelidaceae)植物四药门花〔Loropetalumsubcordatum(Benth.) Oliv.〕的遗传多样性进行了研究,并为其制定了保护策略。
鉴于此,本研究采用SRAP标记分析来自福建、湖南、广东、江西和广西的半枫荷17个天然种群154个样株的遗传多样性和遗传结构,探讨半枫荷的分布特征对其遗传变异的影响,以期为半枫荷遗传资源保护及开发利用奠定理论基础。
1 材料和方法
1.1 材料
于2018年3月至5月对半枫荷的天然分布区进行全面调查,按照居群间地理距离大于10 km、株间距大于10 m的标准,对分布于福建、湖南、广东、江西和广西的17个天然种群进行采样,各种群的具体地理分布信息见表1。其中,分布于福建长汀、永定、清流、延平、邵武、南靖和周宁7个种群的半枫荷株数较少。每个种群各样株采集100 g无病虫害的当年生嫩叶,剪下后立即放入冰盒,带回实验室后将嫩叶平均分成3份,擦拭干净后用锡箔纸包好,然后用液氮冷冻30 min,最后放入-80 ℃冰箱保存,用于提取基因组DNA。

表1 半枫荷17个天然种群的地理分布信息
1.2 方法
参考Murray等[15]的方法从半枫荷叶中提取基因组DNA,使用Nano-200微量分光光度计(杭州奥盛仪器有限公司)测定DNA的浓度和质量,使用质量体积分数1.0%琼脂糖凝胶电泳检测DNA条带的完整性。使用T100梯度PCR仪(美国Bio-Rad公司)进行扩增。参考董蒙蒙[16]31的方法,从225对SRAP引物组合中筛选出10对多态性丰富、条带清晰的引物组合用于半枫荷154个样株的遗传多样性分析。
PCR反应体系包括:100 ng·μL-1DNA 1.0 μL,2×TaqPCR StarMix with Loading Dye混合液12.5 μL,5 mol·L-1上游和下游引物各1.0 μL,加ddH2O至25.0 μL。PCR反应程序为:94 ℃预变性7 min;94 ℃变性45 s、35 ℃退火1 min、72 ℃延伸1 min,7个循环;94 ℃变性45 s、35 ℃退火1 min、52 ℃延伸1 min,28个循环;最后72 ℃延伸10 min。
PCR反应结束后,取10 μL扩增产物,加入1×TBE电泳缓冲液,在质量体积分数30%的聚丙烯酰胺凝胶上进行电泳分离,然后参考董蒙蒙[16]32的方法银染。使用YGD-Ⅱ医用观片灯(冀州市宏光康复器械厂)观察条带,并用DSC-RX10M4数码相机(日本SONY公司)拍照、保存。
1.3 数据处理
统计电泳图中100~2 000 bp间的条带,某一位点上有条带记为“1”,无条带记为“0”,其中,模糊条带与同区间清晰条带比对,将数据输入EXCEL 2010软件构建原始的“1”、“0”矩阵。
利用POPGEN1.32软件[17]计算观察等位基因数、有效等位基因数、多态性条带百分比、Nei’s遗传多样性指数和Shannon’s多态性信息指数。使用MEGA5.0软件[18]绘制半枫荷17个天然种群和154个样株的聚类图。采用GenAlEx 6.5软件[19]进行Mantel检验,检验半枫荷种群间遗传距离与地理距离的相关性,并用该软件进行分子方差分析(analysis of molecular variance,AMOVA),计算种群间的遗传分化系数和基因流以及种群内和种群间遗传变异的贡献率。采用STRUCTURE2.3.4软件[20]分析遗传结构,分组数(K)设置为1~17,每个K值重复运算10次,500 000次马尔科夫链蒙特卡罗(MC)重复之后进行100 000次burn-in。使用STRUCTURE Harvester确定最佳K值。使用CLUMPP2.0软件[21]将10次运算结果合并,并采用Distruct1.1软件[22]将半枫荷群体遗传结构以图形方式可视化输出。
2 结果和分析
2.1 扩增结果及遗传多样性分析
从225对SRAP引物中筛选出10对用于半枫荷154个样株基因组DNA的扩增,10对SRAP引物的序列及扩增结果见表2。结果显示:共得到235个条带,每对引物扩增的条带数为18~28,平均每对引物扩增出23.5个条带,其中多态性条带179个,平均多态性条带百分比为77.2%。

表2 用于半枫荷154个样株基因组DNA扩增的引物序列及扩增结果
基于SRAP引物扩增结果,半枫荷17个天然种群的遗传多样性分析结果见表3。结果显示:基于SRAP引物检测出的半枫荷17个天然种群的多态性条带数为32~167,均值为113.6,多态性条带百分比(PPB)为17.9%~93.3%,均值为63.5%;观察等位基因数(Na)和有效等位基因数(Ne)分别为1.2~1.9和1.2~1.6,均值分别为1.6和1.5;Nei’s遗传多样性指数(H)和Shannon’s多态性信息指数(I)分别为0.089 4~0.354 4和0.123 9~0.521 0,均值分别为0.250 4和0.366 2,其中,福建连城种群的Nei ’ s遗传多样性指数和Shannon ’ s多态性信息指数均最高,分别为0.354 4和0.521 0。
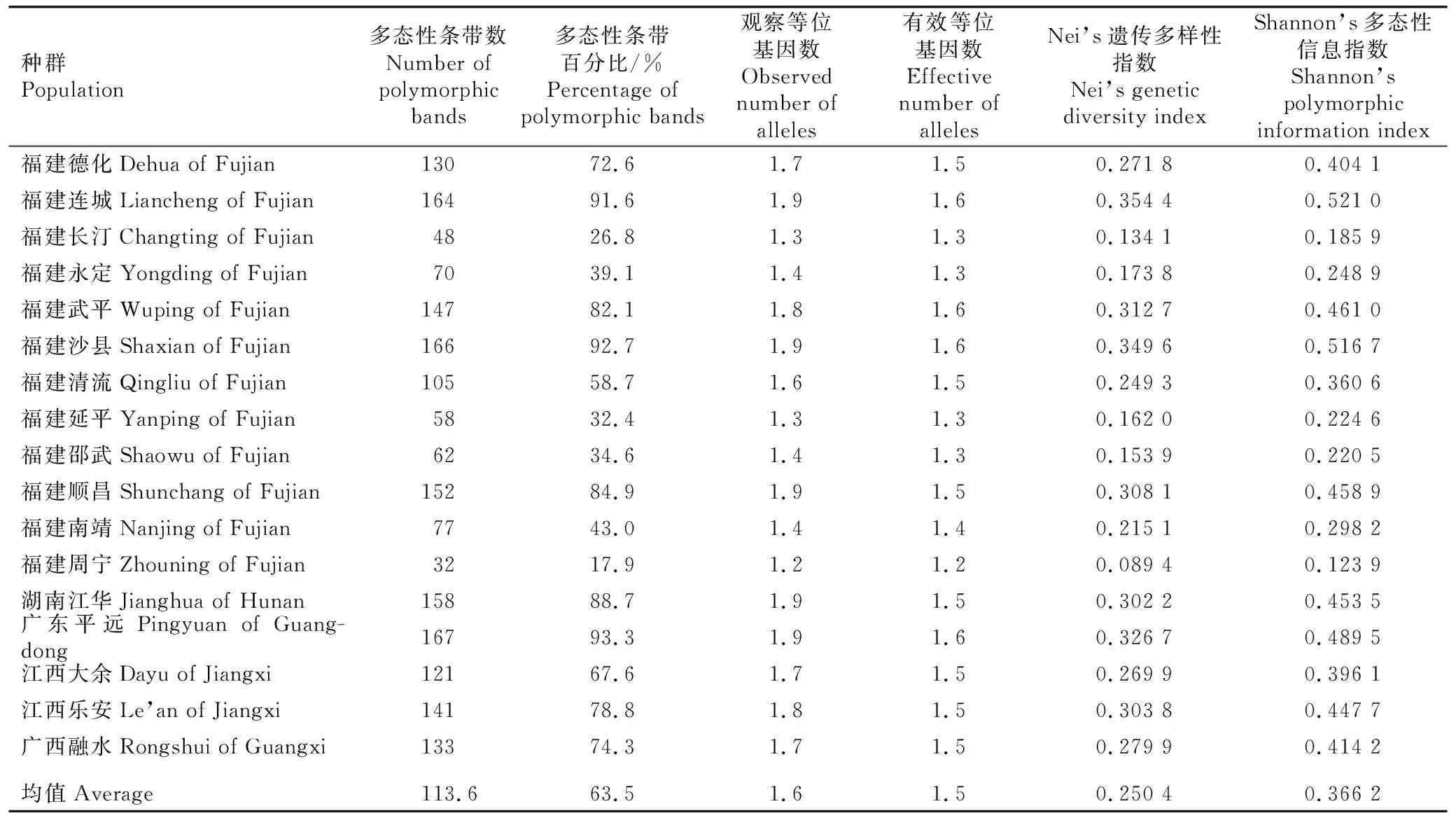
表3 基于SRAP引物扩增结果半枫荷17个天然种群的遗传多样性分析
基于Nei’s遗传多样性指数对半枫荷17个天然种群的遗传多样性进行排序,从大到小依次为福建连城种群、福建沙县种群、广东平远种群、福建武平种群、福建顺昌种群、江西乐安种群、湖南江华种群、广西融水种群、福建德化种群、江西大余种群、福建清流种群、福建南靖种群、福建清流种群、福建永定种群、福建延平种群、福建邵武种群、福建长汀种群和福建周宁种群。
2.2 遗传结构分析
分子方差分析(AMOVA)结果表明:半枫荷17个天然种群间的遗传分化系数为0.241,种群间基因流为1.579,种群内遗传变异的贡献率高达75.95%,而种群间遗传变异的贡献率仅占24.05%,且种群间和种群内的差异达到极显著(P<0.001)水平。说明半枫荷遗传多样性主要分布在种群内,种群间仅出现一定程度的遗传分化。
Mantel检验结果表明:半枫荷17个天然种群的遗传距离和地理距离间无显著相关性(r=0.068,P=0.350),表明地理隔离不是导致半枫荷遗传变异高的主要因子。
Neighbor-joining聚类图(图1)显示:在遗传距离0.18处,半枫荷17个天然种群划分为Ⅰ和Ⅱ 2个集群。集群Ⅰ包括福建的德化、连城、延平、沙县、清流、南靖、周宁、邵武和顺昌种群,江西的乐安和大余种群,广东平远种群以及广西融水种群共13个种群,集群Ⅱ包括湖南江华种群以及福建的长汀、永定和武平种群共4个种群。在遗传距离0.11处,集群Ⅰ可进一步划分为Ⅰa、Ⅰb、Ⅰc和Ⅰd 4个亚群,其中,亚群Ⅰa包括福建的德化和连城种群,江西的乐安和大余种群以及广东平远种群,亚群Ⅰb包括福建的延平、沙县和清流种群,亚群Ⅰc包括福建的南靖和周宁种群,亚群Ⅰd包括福建的邵武和顺昌种群以及广西融水种群;集群Ⅱ可进一步划分为Ⅱa和Ⅱb 2个亚群,其中,湖南江华种群单独构成亚群Ⅱa,亚群Ⅱb包括福建的长汀、永定和武平种群。
UPGMA聚类图(图2)显示:在遗传距离0.65处,半枫荷17个天然种群的154个样株分为2个集群,其中,湖南江华种群中16个样株单独聚为集群Ⅱ,其他138个样株共同聚为集群Ⅰ。在遗传距离0.30处,集群Ⅰ划分为6个亚群,其中,广东平远种群中大部分样株聚为亚群Ⅰc;Neighbor-joining聚类图中福建的长汀、永定和武平种群属于集群Ⅱ,而UPGMA聚类图中这3个种群的样株在集群Ⅰ中,基本聚在亚群Ⅰe。

DH: 福建德化Dehua of Fujian; LC: 福建连城Liancheng of Fujian; LA: 江西乐安Le’an of Jiangxi; DY: 江西大余Dayu of Jiangxi; PY: 广东平远Pingyuan of Guangdong; YP: 福建延平Yanping of Fujian; SX: 福建沙县Shaxian of Fujian; QL: 福建清流Qingliu of Fujian; NJ: 福建南靖Nanjing of Fujian; ZN: 福建周宁Zhouning of Fujian; SW: 福建邵武Shaowu of Fujian; RS: 广西融水Rongshui of Guangxi; SC: 福建顺昌Shunchang of Fujian; JH: 湖南江华Jianghua of Hunan; CT: 福建长汀Changting of Fujian; YD: 福建永定Yongding of Fujian; WP: 福建武平Wuping of Fujian.图1 半枫荷17个天然种群的Neighbor-joining聚类图Fig. 1 Neighbor-joining dendrogram of 17 natural populations of Semiliquidambar cathayensis Chang
使用STRUCTURE软件检测供试半枫荷17个天然种群的遗传结构,结果(图3)显示:K=2时,ΔK值最大;K=7时,ΔK值次之(图3-A)。K=7时,半枫荷的遗传结构聚类较清晰(图3-B),说明半枫荷17个天然种群可先划分为2大集群,再进一步细分为7个亚群。
综合Neighbor-joining聚类图、UPGMA聚类图和STRUCTURE分析结果,可将半枫荷17个天然种群先划分为2个集群,然后在Neighbor-joining聚类图的划分基础上,将亚群Ⅱa中的广东平远种群单独划分为1个亚群,共7个亚群。


DH: 福建德化Dehua of Fujian; LA: 江西乐安Le’an of Jiangxi; DY: 江西大余Dayu of Jiangxi; LC: 福建连城Liancheng of Fujian; CT: 福建长汀Changting of Fujian; YD: 福建永定Yongding of Fujian; WP: 福建武平Wuping of Fujian; NJ: 福建南靖Nanjing of Fujian; ZN: 福建周宁Zhouning of Fujian; SX: 福建沙县Shaxian of Fujian; QL: 福建清流Qingliu of Fujian; YP: 福建延平Yanping of Fujian; SC: 福建顺昌Shunchang of Fujian; SW: 福建邵武Shaowu of Fujian; RS: 广西融水Rongshui of Guangxi; JH: 湖南江华Jianghua of Hunan; PY: 广东平远Pingyuan of Guangdong.图3 半枫荷17个天然种群的分组(A)和遗传结构(B)分析Fig. 3 Analyses on cluster (A) and genetic structure (B) of 17 natural populations of Semiliquidambar cathayensis Chang
3 讨论和结论
3.1 半枫荷天然种群的遗传多样性
通常情况下,濒危植物由于瓶颈效应、近亲繁殖、奠基者效应和遗传漂变等原因,其遗传多样性较低,如细果秤锤树(SinojackiamicrocarpaC. T. Chen et G. Y. Li)〔Nei’s遗传多样性指数(H)为0.100 7,Shannon’s多态性信息指数(I)为0.165 8〕[23]和水青树(TetracentronsinenseOliv.)(H值为0.076,I值为0.112)[24]。本研究中,半枫荷天然种群具有较高水平的遗传多样性(H值为0.250 4,I值为0.366 2)。且半枫荷其他近缘种的遗传多样性也普遍较高,如银缕梅〔Parrotiasubaequalis(H. T. Chang) R. M. Hao et H. T. Wei〕(H值为0.203 1,I值为0.313 2)[25]和檵木〔Loropetalumchinense(R. Br.) Oliv.〕(H值为0.65)[26]。蕈树科是超蔷薇类(Superosids)植物的基部类群,半枫荷遗传多样性较高可能是蕈树科植物作为一个古老树种系统发育的结果,也可能是半枫荷为属间天然杂交种,还可能与第四纪冰期半枫荷种群结构相对稳定、现存种群为其避难所残迹有关[6-7]。遗传多样性可能与自然环境中种群大小或密度等变量有关,但难以定量测定这些变量对遗传多样性的影响。福建的连城、武平、沙县和顺昌种群以及广东平远种群的遗传多样性较高且种群中半枫荷株数多,其中,福建的连城、武平、沙县和顺昌种群主要分布在高海拔、陡坡密林处,生境破坏程度较小,推测人为干扰较少是这4个种群遗传多样性较高的重要原因。
半枫荷生境总体受人为干扰较大,其天然种群呈现破碎化间断分布,种群规模通常较小,甚至部分为孤立木,迫使半枫荷自交繁殖。近亲繁殖会导致濒危植物种群数量减少,降低其遗传多样性[27]。半枫荷在群落内常为非优势种,在竞争光热资源时常处于弱势,加之对光热的需求苛刻,不利于其生殖生长和种群规模的扩大,进而影响其遗传多样性。
3.2 半枫荷种群遗传结构和遗传分化
半枫荷遗传变异主要存在于种群内(贡献率75.95%),种群间的遗传变异相对较小(贡献率24.05%)。但其近缘植物四药门花[14]仅15%的变异来自种群内,说明半枫荷种群内的遗传变异较为丰富,具有较丰富的可开发利用的遗传资源。半枫荷属于混合交配的繁育系统,但其种群间遗传分化系数为0.241,说明其种群内遗传分化水平较高,可能与该种对生境要求特殊以及分布区的地形地貌有关。半枫荷零星分布于武夷山脉、戴云山脉、南岭山脉、雩山山脉、罗霄山脉、十万大山、雪峰山脉、苗岭、大瑶山脉及黎母山脉,多生长在空气湿度大的沟谷、密林(阳坡)及竹林中,各种群被走向多样的高山阻隔。K=2~7时,半枫荷17个种群遗传结构的划分均存在较大的杂合性,K=2时,虽然ΔK值最大,却不能显示清晰的遗传结构,K=7时,各种群间的遗传分化更明显,因此,这17个种群可划分为7个基因库。半枫荷全基因组杂合率为2.93%,重复序列约占68.87%(未发表数据),表明半枫荷遗传背景复杂、杂合性高,与半枫荷为枫香树属与蕈树属天然杂交种的结论相吻合[7]。植物繁育类型是植物种群遗传变异模式的决定性因子[28]。异交物种的遗传分化系数平均值为0.19[29],而自交物种的遗传分化系数平均值为0.414 0[30],表明半枫荷很可能属于兼性自交的繁育系统,这与调查中发现半枫荷多数植株结实率低相吻合,且半枫荷极可能是自交亲和。Schmitt等[31]研究半枫荷同科植物北美枫香(LiquidambarstyracifluaLinn.)的自花不育特征,认为自交会导致果实中种子数极大减少。因此,半枫荷结实率低可能由于自交率较高和属间天然杂交亲和性低所致。本研究中,半枫荷种群间基因流为1.579,大于1,说明种群间基因交流强度适中,也说明种群间没有显著的遗传分化,因此,半枫荷天然种群的遗传多样性水平对遗传漂变不敏感。
综合Neighbor-joining聚类图、UPGMA聚类图和STRUCTURE分析结果,半枫荷17个天然种群先划分为Ⅰ和Ⅱ 2个集群,集群Ⅰ进一步划分为5个亚群,其中,亚群Ⅰa包括福建的德化和连城种群以及江西的乐安和大余种群,亚群Ⅰb仅包括广东平远种群,亚群Ⅰc包括福建的延平、沙县和清流种群,亚群Ⅰd包括福建的南靖和周宁种群,亚群Ⅰe包括福建的邵武和顺昌种群以及广西融水种群;集群Ⅱ进一步划分为2个亚群,其中,湖南江华种群单独构成亚群Ⅱa,亚群Ⅱb包括福建的长汀、永定和武平种群。虽然采样不均匀时,STUCTURE软件计算结果的可靠性可能会降低[32],但采用3种方法对半枫荷17个天然种群遗传结构的划分结果基本一致,只是在部分样株划分上有差异,因此,本研究对半枫荷17个天然种群的遗传结构划分结果可信度较高。值得注意的是,尽管广西融水种群与福建的邵武和顺昌种群在地理位置上距离很远,但聚为同一亚群,并且广西融水种群和福建顺昌种群的遗传多样性较高,这可能是由于这3个种群来自同一祖先。STRUCTURE分析结果显示:广西融水种群以及福建的邵武和顺昌种群来自同一个基因池,也证明这3个种群的祖先可能相同。可见聚类结果与地理位置有一定相关性,但并不显著,同时Mantel检验结果也表明半枫荷种群间遗传距离和地理距离无显著相关性。
本研究中,10对SRAP引物对半枫荷154个样株共扩增出235个条带,179个条带具有多态性,平均多态性条带百分比为77.2%,说明本研究可靠性较高。但由于海南、广西、贵州和湖南等省、自治区半枫荷的具体分布信息未能完全掌握,加之本研究中7个半枫荷种群的半枫荷株数较少,导致本研究取样存在不足,对研究结论的准确性造成一定的影响。作者所在团队将在半枫荷全基因组解析完成后开展半枫荷基因组重测序工作,以期全面解析其遗传多样性、遗传结构和谱系地理,弥补本研究的不足。
3.3 半枫荷遗传资源保护
外业调查发现,半枫荷生境渐趋恶化、破碎化日益严重、结实周期长短不一、种子空粒较多、大多数半枫荷周围未发现小苗、且多为孤立木,结合本研究结果,虽然半枫荷遗传多样性较高,但自交概率较高,种群衰退趋势明显,半枫荷天然种群亟需保护与复壮。具体建议如下:首先,应加快完成迁地保护,尽可能收集所有种群的种子,保存其遗传资源完整性。本研究团队已在福建省顺昌县建立半枫荷种质资源库,接下来还将进一步扩大种质资源收集范围。其次,应立即实施就地保护,保护原生境。遗传多样性较高的福建的连城、沙县、武平和顺昌种群,湖南江华种群,广西融水种群以及广东平远种群应给予优先保护。此外,福建德化、长汀和清流等小种群应严格保护(如采取建立保护小区、封山育林及回归定植等措施),禁止一切破坏行为。
