不同种源辽东栎种子和幼树指标变异及相关分析
2021-09-16郝向春韩丽君陈天成
郝向春, 周 帅, 韩丽君, 翟 瑜, 陈天成
(山西省林业和草原科学研究院, 山西 太原 030012)
辽东栎(QuercusliaotungensisKoidz.)隶属于壳斗科(Fagaceae)栎属(QuercusLinn.),为落叶乔木,是华北中低山地带营造针阔混交林的良好树种。辽东栎天然更新以萌蘖为主,但幼苗生长退化严重,加上其在土壤种子库中因动物觅食、虫蛀和生理死亡等原因而丧失[1-2],致使辽东栎种群自然更新困难,现阶段,辽东栎的林分更新和营建主要依靠人工繁育方式。已有研究证实,辽东栎存在丰富的种源间变异[3-5],这为研究者开展辽东栎优良种源筛选提供了必要的条件。
种源试验是林木育种的常用方法之一。通过种源试验,可以研究树种的表型性状和光合生理特性等在种源间的差异及其与生态环境的关系,为提高树种在栽植地的生产能力和适应能力提供理论支撑。自然状态下,辽东栎的成年植株常出现分枝过早、疤节过多的现象。研究表明:木材性状在林分内的个体间差异比种源间差异大[6],且较少的分枝数量和较长的节间长度有利于增加木材无节材的比例[7]。因此,在开展种源试验时,对辽东栎优良个体进行早期筛选,有利于提高辽东栎的木材质量,缩短其林木育种周期。但是,目前能够支撑辽东栎种源选择的基础数据有限,地理和环境条件对辽东栎遗传变异的影响尚不清晰,阻碍了对辽东栎良种的有效选育。
为此,本研究对山西、甘肃、河北、陕西和辽宁5个省份12个辽东栎种源的种子表型性状和发芽率及其幼树的表型性状、叶绿素相对含量(SPAD)和快速叶绿素荧光诱导动力学参数进行变异分析,在此基础上,对这些指标间及其与种源地的地理位置和环境因子进行Pearson相关性分析,并基于不同类型指标对供试辽东栎种源进行Heatmap聚类分析,以期了解辽东栎的表型性状和光合生理指标在种源间和个体间的变异规律,为辽东栎良种选育策略制定提供数据支持。
1 材料和方法
1.1 材料
于2017年秋季,分别在山西、甘肃、河北、陕西和辽宁的自然分布区内选择12个辽东栎野生种源,每个种源在面积1 km2范围内选择至少50株生长旺盛且结实良好的实生植株作为样株,株距在50 m以上。采集样株上、中、下部位的成熟种子,每株1.5 kg以上,将同一种源采集的所有种子混匀。各种源的种源地信息见表1。
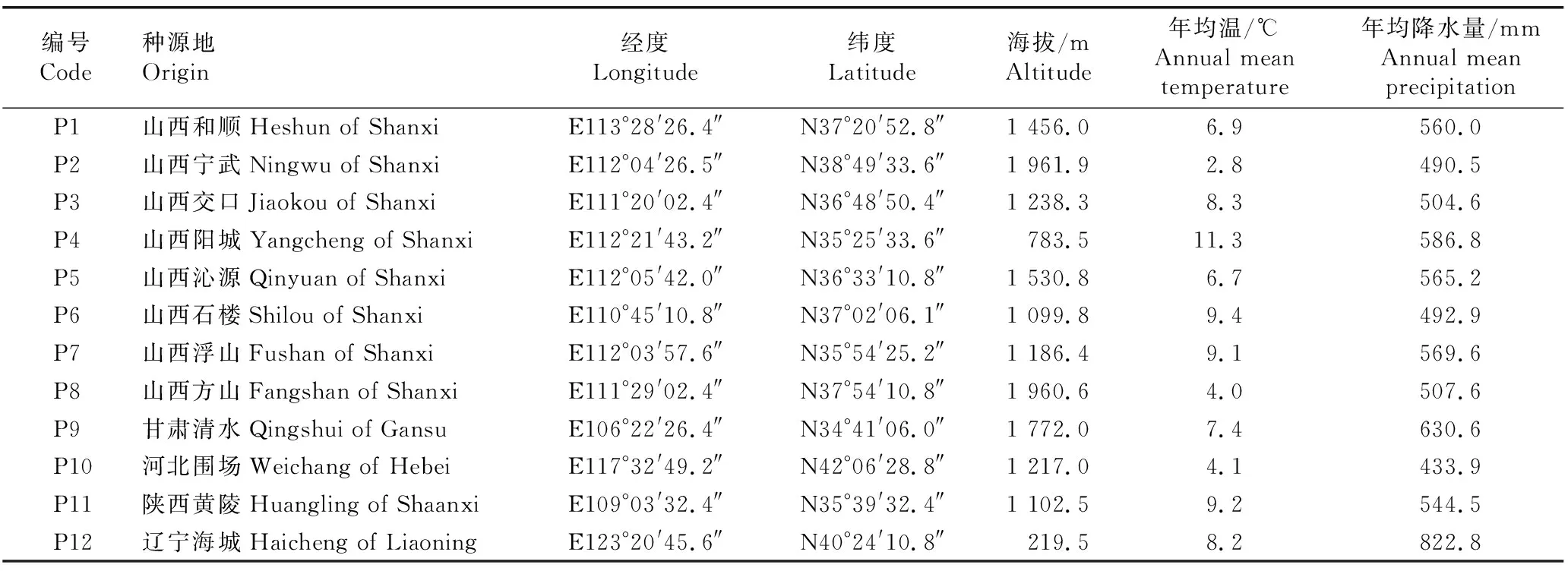
表1 供试各辽东栎种源的种源地信息
1.2 实验基地概况
将辽东栎幼树(株龄1 a)种植在山西省林业和草原科学研究院阳曲实验基地(东经112°44′04.56″、北纬38°05′53.16″,海拔961 m)。该区域属暖温带半干旱大陆性季风气候,年均温8.5 ℃,年均降水量430.4 mm,无霜期164 d。
1.3 方法
1.3.1 种子指标测定 每个种源随机选取成熟、饱满且无虫孔的种子25粒,用蒸馏水洗净;在25 ℃恒温培养箱内,用蒸馏水浸种24 h;吸干种子表面水分,使用千分之一电子天平(美国Ohaus公司)称量单粒种子质量,采用量筒水位增加法[8]测量单粒种子体积,使用游标卡尺(精度0.01 mm)测量种子的长度(即种子纵轴的最大长度)、宽度(种子横轴的最大长度)和碗疤直径(碗疤的最大直径)。每个种源设置4个重复。
每个种源随机选取成熟、饱满、无虫孔且未露白的种子300粒,平均分成3组,每组为1个重复。将种子放入铺有3层滤纸的培养皿(直径14 cm)内,用8 mL蒸馏水湿润滤纸,置于昼温25 ℃、夜温20 ℃、空气相对湿度60%、光照强度60 μmol·m-2·s-1和光照时间16 h·d-1的人工气候箱内催芽。催芽期间适时补水,保持滤纸湿润。以种子露白为发芽标准,连续1周无种子萌发时终止实验,统计发芽种子数,计算发芽率,计算公式为发芽率=(发芽种子数/种子总数)×100%。
1.3.2 幼树指标测定 每个种源随机选取成熟、饱满且无虫孔的种子,用粮虫净(河北金德伦生化科技有限公司)进行除虫处理;于2017年9月,采用随机区组设计将种子播种在实验基地的大田中,大田土壤为褐土。每个种源设置3个重复,每个重复至少播种8 000粒种子。种子间距约20 cm,采用相同的除草、施肥和浇水等田间管理措施。
于2019年10月,在植株生长基本停止时统计植株的单株分枝数量,测量株高(植株根茎结合部至顶芽的距离)、地径(植株根茎结合部的直径)、根系总长度和节间长度(植株自上而下第2轮与第3轮叶片间的距离),称量单株地上部和地下部的干质量,并参照相关文献[11-12]计算壮苗指数和根长比,计算公式分别为壮苗指数=〔(地径/株高)+(单株地下部干质量/单株地上部干质量)〕×(单株地下部干质量+单株地上部干质量)和根长比=根系总长度/(单株地下部干质量+单株地上部干质量)。将植株完整挖出,用蒸馏水洗净根系附着的土壤,使用万深LA-S根系和叶面积分析系统(杭州万深检测科技有限公司)测定株高、地径、根系总长度及节间长度。将植株的地上部与地下部分开,置于105 ℃杀青30 min,再置于75 ℃烘干至恒质量,使用千分之一电子天平分别称量单株地上部和地下部的干质量。
每个种源随机选取10株长势良好的植株测定上述指标,每株各指标均测定1次。
1.3.3 环境因子提取 按照供试各种源地的经度和纬度从WorldClim数据库(https:∥www.worldclim.org/data/worldclim21.html)中提取11个温度因子、8个降水因子及全年12个月的太阳辐射强度因子、平均风速因子和蒸腾压力因子,提取分辨率为10′。从国家青藏高原科学数据中心提供的世界土壤数据库(HWSD)土壤数据集(v1.2)中提取32个土壤因子,包括16个上层(0~30 cm土层)土壤因子和16个下层(30~100 cm土层)土壤因子。比较上述提取因子的相关系数,排除共线性影响,最终筛选出28个环境因子,包括年均温(T1)、昼夜温差月均值(T2)、等温性(T3)、最热月最高温(T4)、年均温变化范围(T5)、年均降水量(R1)、最湿月降水量(R2)、降水量变化方差(R3)、1月份太阳辐射强度(S1)、2月份太阳辐射强度(S2)、5月份太阳辐射强度(S5)、6月份太阳辐射强度(S6)、10月份太阳辐射强度(S10)、6月份平均风速(W6)、8月份平均风速(W8)、6月份蒸腾压力(TP6)、上层土壤碎石体积百分比(US1)、上层土壤沙含量(US2)、上层土壤黏土含量(US3)、上层土壤有机碳含量(US4)、上层土壤pH值(US5)、上层土壤黏性层土壤的阳离子交换能力(US6)、上层土壤阳离子交换能力(US7)、上层土壤交换性盐基(US8)、上层土壤电导率(US9)、下层土壤黏土含量(SS1)、下层土壤有机碳含量(SS2)和下层土壤基本饱和度(SS3)。
1.4 数据统计及分析
利用SPSS 22.0软件进行单因素方差分析和多重比较分析(LSD法)。根据测定结果,计算各指标的种源间变异系数(CV1)和个体间变异系数(CV2),并计算各指标的种源重复力(T),计算公式分别为CV1=(所有种源某指标的标准差/所有种源该指标的平均值)×100%、CV2=(某种源供试植株某指标的标准差/该种源供试植株该指标的平均值)×100%和T=1-(1/某指标种源间方差分析的F值)。利用R 4.0.2软件对辽东栎各指标与种源地的地理位置和环境因子进行Pearson相关性分析;基于不同类型指标,利用Origin 2019软件对供试种源进行Heatmap聚类分析。
2 结果和分析
2.1 不同种源辽东栎种子和幼树指标的比较及变异分析
2.1.1 种子指标的比较及变异分析 对供试不同种源辽东栎的种子指标进行比较和变异分析,结果见表2。结果表明:辽东栎各种子指标在种源间差异较大,发芽率、种子长度、种子宽度、单粒种子质量、单粒种子体积和碗疤直径的种源间差异均极显著(P<0.01)。供试12个种源的发芽率为8.8%~65.9%,种子长度为17.29~42.88 mm,种子宽度为12.05~30.99 mm,单粒种子质量为1 199.2~2 956.2 mg,单粒种子体积为1.16~3.04 cm3,碗疤直径为5.79~9.10 mm,各指标的最大值均极显著高于最小值,分别是最小值的7.50、2.48、2.57、2.46、2.62和1.57倍。其中,山西浮山(P7)种源的种子长度、种子宽度、单粒种子质量和单粒种子体积最小,该种源的碗疤直径和发芽率较小;山西阳城(P4)种源的种子长度最大,该种源其余5个种子指标较大;山西石楼(P6)种源的发芽率和种子宽度最大,该种源其余4个种子指标也较大;河北围场(P10)种源的单粒种子质量、单粒种子体积和碗疤直径最大,该种源的发芽率较大,种子长度和种子宽度居中。
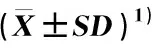
表2 不同种源辽东栎种子指标的比较及变异分析
从各指标的种源间变异系数看,辽东栎各种子指标的种源间变异较大。其中,发芽率的种源间变异系数最大(35.85%),种子宽度的种源间变异系数次之(35.51%),碗疤直径的种源间变异系数最小(14.11%)。从各指标的个体间变异系数看,辽东栎各种子指标的个体间变异也较大。其中,发芽率的个体间变异系数最大(37.04%),种子宽度的个体间变异系数次之(36.45%),碗疤直径的个体间变异系数最小(18.95%)。经计算,供试6个种子指标的种源间变异系数均值为27.81%,明显小于其个体间变异系数均值(32.36%)。
从各指标的种源重复力看,辽东栎各种子指标的种源重复力均大于0.9,其中,种子长度和种子宽度的种源重复力相同且最高(0.99),而种子发芽率的种源重复力最低(0.96)。
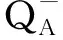
表3 不同种源辽东栎幼树指标的比较及变异分析
从各指标的种源间变异系数看,辽东栎各幼树指标的种源间变异明显不同。其中,壮苗指数的种源间变异系数最大(48.98%),RC/CSo的种源间变异系数次之(33.29%),SPAD的种源间变异系数最小(5.47%)。从各指标的个体间变异系数看,辽东栎各幼树指标的个体间变异也明显不同。其中,壮苗指数的个体间变异系数最大(80.80%),节间长度的个体间变异系数次之(45.20%),TRo/RC的个体间变异系数最小(8.31%)。经计算,供试13个幼树指标的种源间变异系数均值为20.58%,明显小于其个体间变异系数均值(30.01%)。
由表2可见:辽东栎各幼树指标的种源重复力差异较大,其中,壮苗指数和RC/CSo的种源重复力相同且最高(0.95),单叶面积的种源重复力也较高(0.94),单株分枝数的种源重复力最低(0.33),其余指标的种源重复力介于0.3~0.7。
2.2 辽东栎种子和幼树指标间及其与种源地地理位置和环境因子的相关性分析
2.2.1 种子和幼树指标间的相关性分析 对辽东栎的种子和幼树指标进行Pearson相关性分析,相关系数达到显著(P<0.05)和极显著(P<0.01)水平的结果见表4。结果表明:种子长度与发芽率呈显著正相关;种子宽度与发芽率呈显著正相关,与种子长度呈极显著正相关;单粒种子体积与单粒种子质量呈极显著正相关;碗疤直径与单粒种子质量和单粒种子体积呈极显著正相关;壮苗指数与单粒种子体积呈显著正相关,与碗疤直径呈极显著正相关;单叶面积与单粒种子体积和壮苗指数呈显著正相关,与碗疤直径呈极显著正相关。
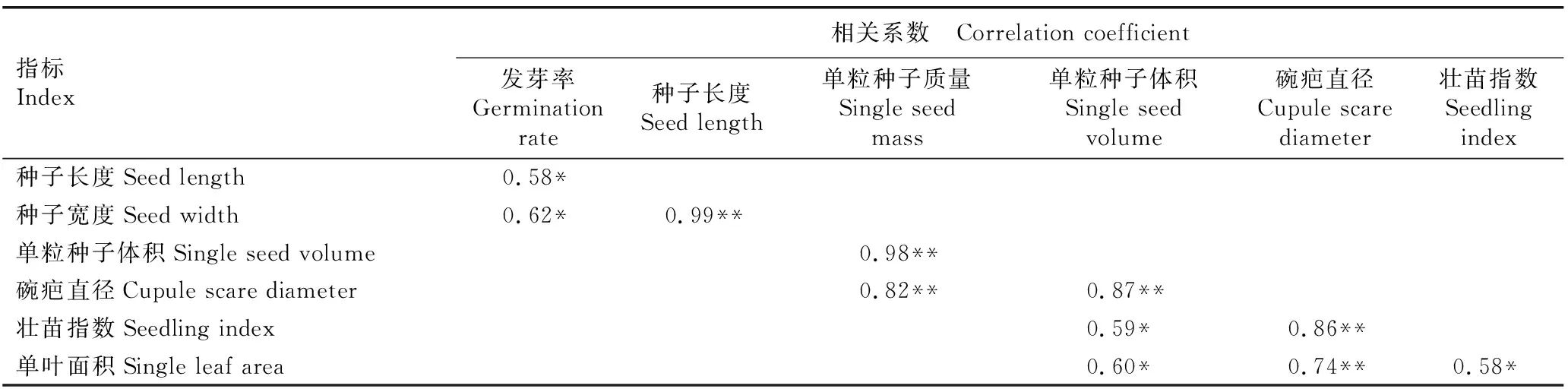
表4 辽东栎种子和幼树指标间的相关系数1)
2.2.2 种子和幼树指标与种源地气候因子的相关性分析 对辽东栎种子和幼树指标与种源地气候因子进行Pearson相关性分析,相关系数达到显著和极显著水平的结果见表5。结果表明:发芽率与昼夜温差月均值、5月份太阳辐射强度、6月份太阳辐射强度和10月份太阳辐射强度均呈显著正相关;单粒种子质量与年均温变化范围呈显著正相关;单粒种子体积与年均温变化范围呈显著正相关,与1月份太阳辐射强度呈显著负相关;碗疤直径与等温性呈显著负相关,与年均温变化范围呈极显著正相关,与降水量变化方差呈显著正相关,与1月份太阳辐射强度呈极显著负相关;壮苗指数与等温性和1月份太阳辐射强度呈极显著负相关,与2月份太阳辐射强度呈显著负相关,与年均温变化范围呈显著正相关,与最湿月降水量呈极显著正相关;单叶面积与昼夜温差月均值呈显著正相关,与年均温变化范围呈极显著正相关;叶绿素相对含量(SPAD)与年均温呈显著正相关,与最热月最高温呈极显著正相关。
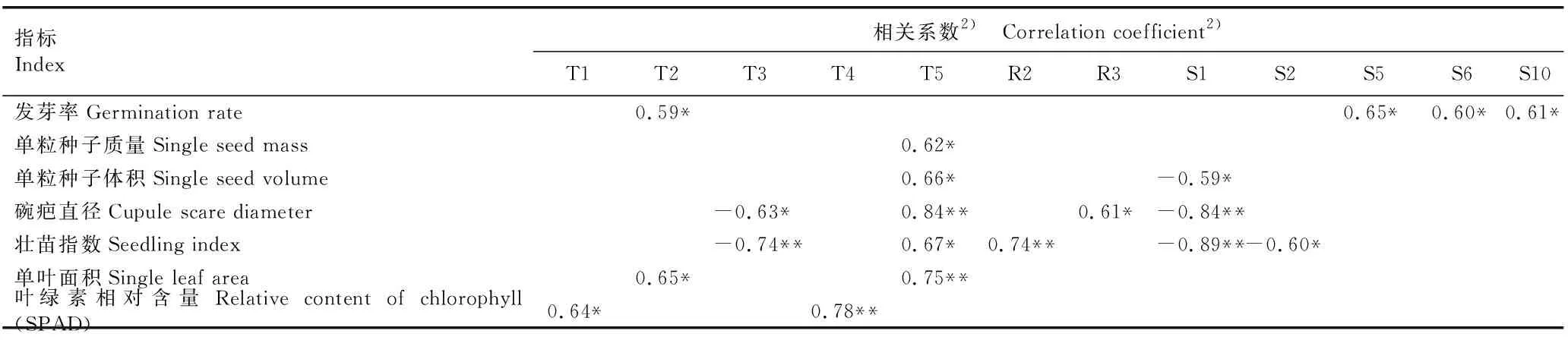
表5 辽东栎种子和幼树指标与种源地气候因子的相关系数1)
2.2.3 种子和幼树指标与种源地地理位置和土壤因子的相关性分析 对辽东栎种子和幼树指标与种源地的地理位置和土壤因子进行Pearson相关性分析,相关系数达到显著和极显著水平的结果见表6。结果表明:种子长度与上层土壤阳离子交换能力和上层土壤交换性盐基呈显著正相关;碗疤直径与经度和纬度呈极显著正相关,与上层土壤黏性层土壤的阳离子交换能力和上层土壤阳离子交换能力呈显著负相关;壮苗指数与经度和纬度呈极显著正相关,与上层土壤沙含量呈显著正相关,与海拔、上层土壤黏土含量、上层土壤黏性层土壤的阳离子交换能力、上层土壤阳离子交换能力和上层土壤交换性盐基呈显著负相关;单叶面积与纬度呈显著正相关,与上层土壤黏性层土壤的阳离子交换能力呈显著负相关,与下层土壤基本饱和度呈极显著负相关;性能指数与上层土壤有机碳含量呈显著负相关,与上层土壤酸碱度呈显著正相关;SPAD仅与海拔呈极显著负相关。

表6 辽东栎种子和幼树指标与种源地地理位置和土壤因子的相关系数1)
2.3 辽东栎不同种源的聚类分析
基于种子指标、幼树指标(其中快速叶绿素荧光诱导动力学参数只选择性能指数1个指标)和种源地环境因子分别对供试辽东栎种源进行Heatmap聚类分析,并综合这些指标对供试种源进行Heatmap聚类分析,结果见图1。
从种子指标聚类结果(图1-A)看,供试的12个种源被分成3组,山西和顺(P1)、山西交口(P3)、山西阳城(P4)、山西石楼(P6)和山西宁武(P2)5个种源聚为一组,河北围场(P10)和辽宁海城(P12)2个种源聚为一组,山西沁源(P5)、山西浮山(P7)、甘肃清水(P9)、山西方山(P8)和陕西黄陵(P11)5个种源聚为一组。从幼树指标聚类结果(图1-B)看,供试的12个种源被分成2组,P10和P12种源聚为一组,其余种源聚为另一组,并且,前组各种源的壮苗指数明显高于后组。从种源地环境因子聚类结果(图1-C)看,供试的12个种源被分成3组,P10和P12种源分别单独为一组,其余种源聚为一组。从综合聚类结果(图1-D)看,P10和P12种源也分别单独为一组,其余种源聚为一组,与种源地环境因子的聚类结果相似。
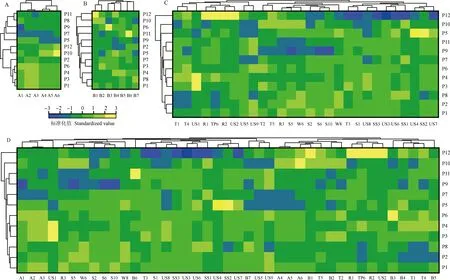
P1: 山西和顺Heshun of Shanxi; P2: 山西宁武Ningwu of Shanxi; P3: 山西交口Jiaokou of Shanxi; P4: 山西阳城Yangcheng of Shanxi; P5: 山西沁源Qinyuan of Shanxi; P6: 山西石楼Shilou of Shanxi; P7: 山西浮山Fushan of Shanxi; P8: 山西方山Fangshan of Shanxi; P9: 甘肃清水Qingshui of Gansu; P10: 河北围场Weichang of Hebei; P11: 陕西黄陵Huangling of Shaanxi; P12: 辽宁海城Haicheng of Liaoning.A1: 发芽率Germination rate; A2: 种子长度Seed length; A3: 种子宽度Seed width; A4: 单粒种子质量Single seed mass; A5: 单粒种子体积Single seed volume; A6: 碗疤直径Cupule scare diameter. B1: 壮苗指数Seedling index; B2: 单叶面积Single leaf area; B3: 根长比Root length ratio; B4: 叶绿素相对含量Relative chlorophyll content (SPAD); B5: 节间长度Internode length; B6: 单株分枝数Branch number per plant; B7: 性能指数Performance index on absorption basis. T1: 年均温Annual mean temperature; T2: 昼夜温差月均值Monthly mean diurnal range of temperature; T3: 等温性Isothermality; T4: 最热月最高温The maximum temperature of the hottest month; T5: 年均温变化范围Annual mean temperature range; R1: 年均降水量Annual mean precipitation; R2: 最湿月降水量Precipitation of the wettest month; R3: 降水量变化方差Variance of precipitation variation; S1: 1月份太阳辐射强度Solar radiation intensity in Jan.; S2: 2月份太阳辐射强度Solar radiation intensity in Feb.; S5: 5月份太阳辐射强度Solar radiation intensity in May; S6: 6月份太阳辐射强度Solar radiation intensity in Jun.; S10: 10月份太阳辐射强度Solar radiation intensity in Oct.; W6: 6月份平均风速Mean wind speed in Jun.; W8: 8月平均风速Mean wind speed in Aug.; TP6: 6月份蒸腾压力Transpiration pressure in Jun.; US1: 上层土壤碎石体积百分比Volume percentage of gravel in upper soil; US2: 上层土壤沙含量Sand content in upper soil; US3: 上层土壤黏土含量Clay content in upper soil; US4: 上层土壤有机碳含量Organic carbon content in upper soil; US5: 上层土壤pH值pH value of upper soil; US6: 上层土壤黏性层土壤的阳离子交换能力Cation exchange capacity of soil in clay layer of upper soil; US7: 上层土壤阳离子交换能力Cation exchange capacity of upper soil; US8: 上层土壤交换性盐基Total exchangeable bases of upper soil; US9: 上层土壤电导率Electrical conductivity of upper soil; SS1: 下层土壤黏土含量Clay content in subsoil; SS2: 下层土壤有机碳含量Organic carbon content in subsoil; SS3: 下层土壤基本饱和度Basic saturation of subsoil.A: 基于种子指标的聚类结果Cluster result based on seed indexes; B: 基于幼树指标的聚类结果Cluster result based on sapling indexes; C: 基于种源地环境因子的聚类结果Cluster result based on environmental factors of origins; D: 基于种子指标、幼树指标和种源地环境因子的聚类结果 Cluster result based on seed indexes, sapling indexes, and environmental factors of origins.图1 辽东栎不同种源的Heatmap聚类分析Fig. 1 Heatmap cluster analysis on different provenances of Quercus liaotungensis Koidz.
3 讨 论
研究发现,中国的西部地区、南方湿润区和高海拔地区均不适宜辽东栎生长[13]。本研究中,辽东栎幼树的壮苗指数与种源地经度和纬度呈极显著正相关,与海拔呈显著负相关。另外,随着海拔升高,辽东栎叶片的叶绿素相对含量(SPAD)降低;随着经度升高,辽东栎种子的碗疤直径增大;随着纬度升高,辽东栎种子的碗疤直径和幼树的单叶面积均增大。值得注意的是,壮苗指数与表征种子营养丰富程度的单粒种子体积和碗疤直径分别呈显著或极显著正相关。据此推测,在辽东栎分布区内,随着经度和纬度升高,辽东栎种子碗疤直径和幼树叶片面积明显增大,其光合产物的积累量及其在种子中的储存量也随之增高。另外,高海拔区域的辽东栎幼树叶片的叶绿素极可能因受到过量紫外线辐射而加速分解,导致光合产物积累量减少,影响种子营养储备。这些因子对辽东栎种子营养物质储备的影响,导致其幼树性状出现差异,进而影响辽东栎的天然分布格局。
种子大小对种子萌发具有重要影响[14]。辽东栎的种子质量与种子的长度、宽度和发芽率的相关性较高[15-17]。本研究中,辽东栎的种子长度和种子宽度与发芽率呈显著正相关,表明从种子长度和种子宽度看,辽东栎大粒种子的发芽率优于小粒种子,但单粒种子体积与发芽率的相关性却不显著,具体原因有待深入研究。碗疤对栎属植物种子与母株间的物质交流至关重要[8]。供试12个种源辽东栎种子的碗疤直径与单粒种子质量、单粒种子体积、壮苗指数、单叶面积以及种源地的等温性、年均温变化范围、降水量变化方差、1月份太阳辐射强度、经度、纬度、上层土壤黏性层土壤的阳离子交换能力和上层土壤阳离子交换能力显著或极显著相关,可见,辽东栎种子的碗疤直径对生境变化敏感,可通过碗疤直径大小推测其种子的养分储备状态。
李梅[3]的研究结果表明:辽东栎的群体间表型变异仅达到群体内表型变异的59.41%,而本研究中辽东栎幼树指标的种源间变异系数均值(20.58%)占个体间变异系数均值(30.01%)的68.58%,说明辽东栎育种应侧重于单株选择。此外,单株分枝数和节间长度的个体间变异系数分别较种源间变异系数增加58.63%和46.90%,说明辽东栎无节材育种也应侧重于单株选择。
温度[18-19]、水分[20-21]、盐分[22-23]、光照[24-25]、昆虫取食[26-27]及酸雨[28]等环境胁迫均能诱导植物快速叶绿素荧光诱导动力学参数显著变化。Kurjak等[29]的研究结果表明:欧洲山毛榉(FagussylvaticaLinn.)的PSⅡ最大光化学效率(Fv/Fm)和PIABS的种源间差异显著。本研究中,辽东栎幼树的多个快速叶绿素荧光诱导动力学参数的种源间差异显著,例如:山西方山(P8)种源幼树的PIABS极显著高于山西沁源(P5)种源,而其PSⅡ单位反应中心耗散的能量(DIo/RC)却显著低于P5种源,这可能是因为P5种源幼树在实验地受到环境胁迫,影响了光合电子传递效率,为避免过量光能诱导的活性氧破坏光合机构,P5种源幼树提高了DIo/RC。本研究还发现,辽东栎幼树的RC/CSo的种源重复力达0.95,且该指标的种源间差异极显著,预示着快速叶绿素荧光测量技术可用于辽东栎种源的快速鉴定。对辽东栎种源进行早期选择能大幅缩短其育种周期,但关于辽东栎各种源幼树指标在后期生长中的延续程度尚不清楚,需要进一步深入研究。另外,尽管本研究采用大量混合采种方法来减少种源内的个体间差异,但仍然无法消除同一地区不同半同胞家系间的基因差异可能对研究结果产生的影响,后续研究应采取有效措施避免这方面的影响。
