滴灌土壤基质势调控对红豆杉生理特性及次生代谢物含量的影响
2021-09-16张玉豪姚素梅
张玉豪,姚素梅,孟 丽,邓 哲,黄 玲
(河南科技学院生命科技学院,河南 新乡 453003)
红豆杉(TaxuschinensisRehd)为我国一级保护植物,因其植株体内含有抗癌活性物质紫杉醇而成为近些年人们研究和开发的热点之一。另外,红豆杉中还含有生物活性较强的黄酮类和多糖类化合物, 对抗肿瘤、提高免疫力、治疗和预防高血糖等疾病也有一定的效果[1]。但在过去的几十年里,野生红豆杉遭到人类的过度开发,自然条件下红豆杉生长缓慢,种群竞争力和再生能力也较弱,人类活动的增加减少了野生红豆杉的种质资源,使野生红豆杉资源呈濒危状态[2-3]。因此,加大红豆杉的人工驯化栽培技术研究对红豆杉种质资源的恢复具有十分重要的意义。
滴灌是现代节水灌溉技术的一种,利用滴灌技术可以制定出合理且便于实施的灌溉制度,使作物根系能够长时间处于最佳水分和养分状态[4],因此在农业生产中得到了广泛应用[5-6]。土壤基质势能够反映出土壤中的水分状况,康跃虎[7]经过多年研究,总结出了利用负压计监测土壤基质势来指导灌溉的滴灌灌溉制度。但前人利用滴灌土壤基质势对植物的调控研究多偏重于温室蔬菜和常见作物[8-9],且前人对红豆杉栽培技术的研究多集中在不同土壤基质[10]、光照[11]和温度[12]对红豆杉生理和生长的影响,而滴灌土壤基质势调控对红豆杉生理特性和次生代谢物含量的影响研究报道甚少。本试验以盆栽太行红豆杉为材料,在自然条件下,通过滴灌技术调控土壤基质势下限,探究滴灌土壤基质势调控对红豆杉叶片生理特性及次生代谢物含量的影响,为红豆杉的人工驯化栽培提供理论依据。
1 材料与方法
1.1 试验区概况
试验于2019年3—8月和2020年6—11月在河南科技学院药用植物栽培场(113°54′ E,35°18′ N)进行,该栽培场位于河南省新乡市,地属大陆性季风气候,多年平均气温14.4℃,多年平均降水量600 mm左右。栽培场采用加密6针型遮阳网遮阴以模拟红豆杉在自然条件下的生长环境,其中遮阳棚采用钢架制造,高约2.5 m。
1.2 试验设计
试验材料为5 a生盆栽太行红豆杉,2019年选取生长一致且良好的太行红豆杉60盆进行试验,其中,红豆杉的平均株高为74.8 cm,盆栽所用盆的直径和高度均为30 cm,所用基质的成分为稻壳∶土∶鸡粪=3∶3∶1,2020年重新选取与2019年所用材料相似且生长一致的5 a生太行红豆杉盆栽进行重复试验。试验时,将红豆杉盆栽均匀分成5组,每组12盆。在每组盆栽中埋入1根DLS系列负压计(滴灌滴头正下方20 cm处),在自然条件下,通过监测负压计读数,将5组盆栽的土壤基质势下限分别控制在-10 kPa(W1)、-20 kPa(W2)、-30 kPa(W3)、-40 kPa(W4)和-50 kPa(W5)。每天8∶00和17∶00记录负压计读数,当负压计读数低于该处理的土壤基质势下限时,利用滴箭滴灌系统对红豆杉盆栽进行灌溉,每次每盆灌溉量为5 mm,约为353 ml,用k24小型流量计检测灌水量。
红豆杉生长缓慢,试验开始后,每月测量1次红豆杉叶片的光合特性、生理指标和次生代谢物(黄酮和多糖)含量,取具有代表性月份的数据进行分析。由于本试验在自然条件下进行,降雨会打破试验设定的土壤基质势梯度,强降雨更会使各处理土壤基质势增大到一致。为避免降雨对试验生理特性测定的影响,每月的测量日要根据降雨等天气因素进行调整,在各处理土壤基质势基本处于试验控制范围内时对红豆杉叶片生理特性进行测定。次生代谢物含量的测量日同生理特性的测量日保持一致。
1.3 测定项目与方法
1.3.1 光合参数和光响应曲线的测量 采用Li-6400型便携式光合测定仪于晴朗天气的上午9∶00—11∶00测定红豆杉叶片的净光合速率(Pn),气孔导度(Gs),胞间二氧化碳浓度(Ci)和蒸腾速率(Tr)等光合参数。测量时,随机选取各处理具有代表性的植株3棵,并取各处理同一冠层处且同年生的叶片进行测量,每颗测量3次,每次测量记录3次数据。由于红豆杉叶片较小不能铺满叶室,测量时同时夹取4片叶子,使4片叶子不重叠且与叶室垂直,测量后用游标卡尺测量叶片宽度,换算出实际叶面积,用于数据处理。
采用Li-6400型便携式光合测定仪的LED红蓝光源叶室测定不同光照强度下红豆杉叶片净光合速率的大小。将光合有效辐射(PAR)从高到低设定为1 600、1 400、1 200、1 000、800、500、200、100、80、40 μmol·m-2·s-1和0 μmol·m-2·s-1,CO2浓度设定为400 μmol·mol-1。在晴朗天气条件下,使用开放气路于上午9∶00—11∶00测量其光响应曲线,测量时,选取各处理同一冠层处且同年生的叶片进行测量,每个处理测定3次。并用叶子飘模型[13]对光响应曲线进行拟合:
(1)
(2)
(3)
(4)
式中,Pn为净光合速率;Pnmax为潜在最大光合速率;α为拟合曲线的斜率,即初始量子效率;I为光合有效辐射;Rd为暗呼吸速率;β为光抑制系数;γ为光饱和系数;LSP为光饱和点;LCP为光补偿点;用式(1)可拟合出弱光强条件下(PAR≤200 μmol·m-2·s-1)的光响应曲线的斜率α,即表观量子效率(AQE)。
2019年取6月份和8月份(试验结束前)作为代表月份进行数据分析,2020年取9月份和11月份作为代表月份进行数据分析,由于试验期间不同月份红豆杉光合特性和叶绿素含量的趋势基本一致,因此2019年红豆杉光合特性和叶绿素含量主要以6月份数据分析为主,2020年以9月份数据分析为主。2019年6月份光合特性的测量日为6月1―2日,该测量日W1、W2、W3、W4和W5的土壤基质势分别为-10.25、-16.75、-20.50、-36.25、-40.00 kPa和-8.75、-12.00、-15.50、-29.50、-33.50 kPa。2020年9月份光合特性的测量日为9月2―3日,该测量日W1、W2、W3、W4和W5的土壤基质势分别为-11.8、-20.8、-26、-38.8、-53.05 kPa和-12.6、-25、-30.5、-38.8、-56.8 kPa。
2019年生理指标和次生代谢物含量的测量日为6月1日和8月28日(试验结束前),8月28日W1、W2、W3、W4和W5的土壤基质势分别是-8.75、-21.75、-32.75、-39.50、-44.50 kPa。2020年测量日为9月2日和11月8日,11月8日W1、W2、W3、W4和W5的土壤基质势分别是-5.8、-14.4、-29.4、-39.4和-48.65 kPa。由于文章篇幅有限,且各生理指标在2019和2020年的趋势基本相似,故叶绿素、可溶性蛋白和丙二醛含量以2019年数据分析为主。
1.3.2 生理指标的测定 采取同一冠层处且同年生的叶片测定各处理的生理指标。叶绿素含量采用分光光度法测定[14],可溶性蛋白含量采用考马斯亮蓝法测定[14],丙二醛(MDA)含量采用硫代巴比妥酸(TBA)显色法测定[14]。
1.3.3 次生代谢物含量的测定 采取各处理同一冠层处且同年生的红豆杉叶片进行测定,黄酮含量采用超声波提取分光光度法测定[15],多糖含量采用苯酚硫酸法[16]进行测定。
1.4 数据分析
采用SAS统计软件中的ANOVA过程对数据进行方差分析。利用Microsoft Excel 2013对数据进行处理和绘图。
2 结果与分析
2.1 滴灌土壤基质势调控对红豆杉叶片生理特性的影响
2.1.1 滴灌土壤基质势调控对红豆杉叶片光合参数的影响 光合作用是植物叶片利用CO2和水生成有机物的过程, 是植物生长和作物产量形成的基础[17]。由表1可知,2019年度,随着土壤基质势的降低,红豆杉叶片的气孔导度和胞间CO2浓度基本呈下降趋势,但显著性有所不同,W1和W2的叶片气孔导度无显著差异,W3~W5的叶片胞间CO2浓度之间无显著差异。不同土壤基质势下限对叶片蒸腾速率的影响最为显著,其趋势与气孔导度相似,5个处理间均存在显著性差异,其中,W5叶片的蒸腾速率比W1降低了63.52%。叶片净光合速率随土壤基质势的降低呈先上升后下降趋势,W2叶片净光合速率最大,分别比W1、W3、W4和W5增大了53.95%、14.89%、52.87%和98.29%,且差异显著。2020年各光合参数的变化规律与2019年度基本一致,表明不同土壤基质势下限能够显著影响红豆杉叶片的光合参数,其中W2显著增大了红豆杉叶片的净光合速率,有利于红豆杉叶片光合作用的进行。
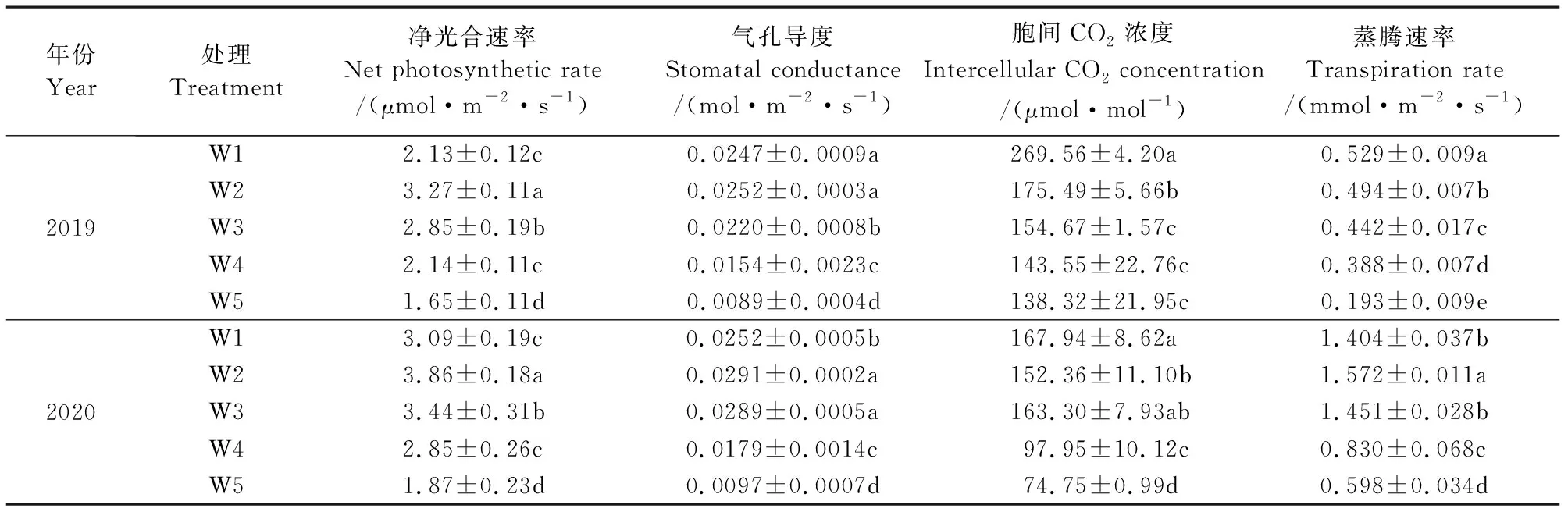
表1 不同土壤基质势对红豆杉叶片光合参数的影响Table 1 Photosynthetic parameters in leaves of Taxus chinensis under different soil matric potentials
2.1.2 滴灌土壤基质势调控对红豆杉叶片光响应曲线的影响 由图1可知,2019和2020两个年度5个处理红豆杉叶片光响应拟合曲线的趋势基本一致,净光合速率随PAR的增大迅速增大,之后增长速度减缓,W2叶片光响应拟合曲线的净光合速率最大,W5最小。表观量子效率AQE是植物对CO2同化的效率,可反映植物光合利用和转化效率的大小,其数值的高低可判断植物吸收和转换光能及利用弱光的能力[18]。暗呼吸速率(Rd)指植物在黑暗条件下的呼吸速率,植物暗呼吸所释放的能量有一部分用于植物的生理活动,因此,Rd数值的大小可反映植物叶片生理活性的高低[19]。由表2可知,2019年度,随着土壤基质势的降低,红豆杉叶片AQE和Rd呈先升高后降低趋势,W2叶片AQE和Rd最大,其AQE比W3、W4和W5分别增长了26.45%、71.32%、80.60%,且差异显著,比W1增长了17.85%,但差异不显著。W2处理的Rd比W3、W4和W5分别增长了20.42%、34.20%和34.36%,且差异显著,比W1增长了13.65%,但无显著差异。植物叶片的光补偿点Lcp和和光饱和点Lsp能够反映植物生长所需的光照条件,可以判断植物的耐阴性和对强光的利用能力,Lcp较低、Lsp较高的植物对光环境的适应性较强[18]。2019年度,随着土壤基质势的降低,红豆杉叶片Lsp趋势与AQE相似,而Lcp趋势则先降后升,其中W2叶片的Lsp最大、Lcp最小。最大净光合速率Pnmax趋势与AQE相似,W2叶片Pnmax显著大于其他处理,比W1、W3、W4和W5分别增长了39.69%、10.91%、69.51%、88.01%。2020年各处理的拟合参数与2019年相比虽然在数值大小上出现轻微变化,但处理间的变化规律与2019年基本一致,表明当土壤基质势较低时,红豆杉叶片对光能的吸收和转换效率降低,对弱光的利用效率也显著减小,W2可显著增强红豆杉叶片的生理活性,升高叶片对光能的吸收和转换效率,且提升叶片对光的生态适应能力,从而使Pnmax达到最大值,有利于红豆杉光合作用的进行。
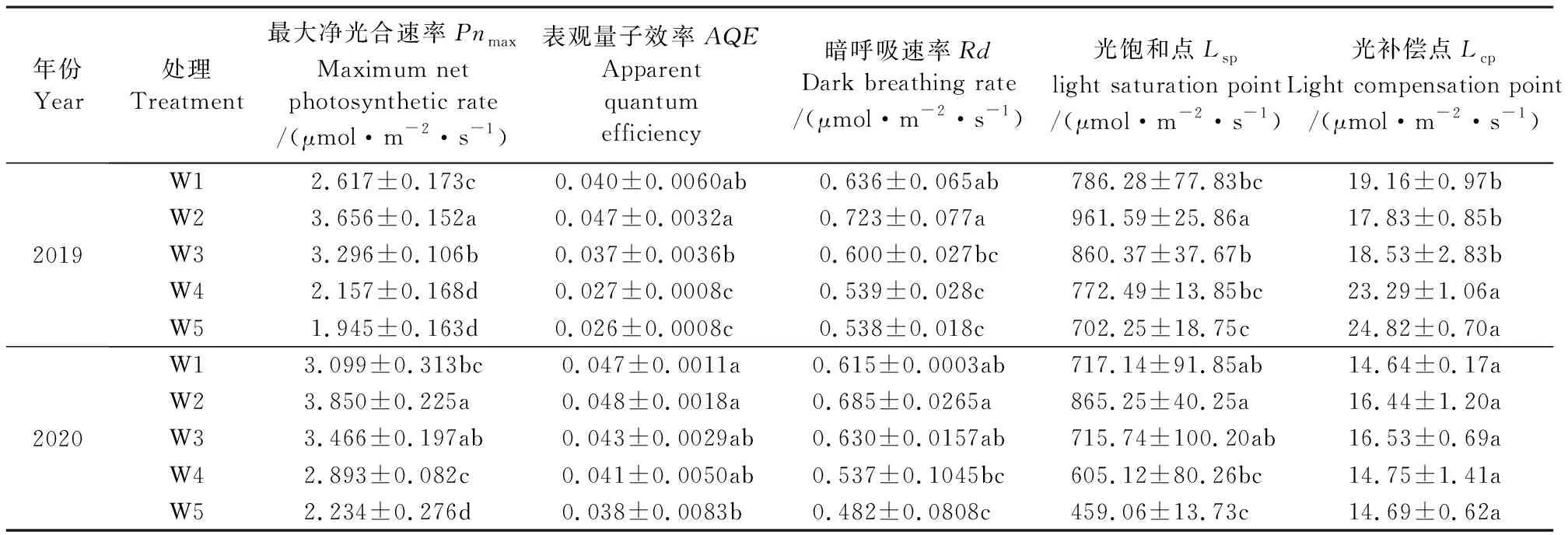
表2 不同土壤基质势对红豆杉叶片光响应曲线拟合参数的影响Table 2 Fitting parameters of light response curve in leaves of Taxus chinensis under different soil matric potentials
2.1.3 滴灌土壤基质势调控对红豆杉叶片叶绿素含量的影响 叶绿素作为绿色植物进行光合作用的主要色素,其含量大小直接影响叶片的光合作用,可以在一定程度上反映植物的光合作用水平[20]。由表3可知,红豆杉叶片叶绿素a含量随土壤基质势的降低整体呈降低趋势,W2叶片叶绿素a含量最大,分别比W1和W3增长了6.80%和9.60%,但差异未达到显著水平,比W4和W5增长了44.81%和41.55%, 且差异显著。随土壤基质势的降低,红豆杉叶片叶绿素b含量先升后降,且在W4达到最大值,分别比W1、W2、W3和W5显著增长了354.65%、44.28%、52.33%、83.99%。红豆杉叶片总叶绿素含量随土壤基质势的降低呈先上升后降低趋势,W2显著提高了叶片总叶绿素含量,分别比W1、W4和W5增长了27.45%、14.29%、37.85%,且差异显著,比W3增长了8.25%,但差异不显著。表明土壤基质势较低时,不利于红豆杉叶片叶绿素a的合成,但适当的水分亏缺有利于红豆杉叶片叶绿素b的合成。本试验中,W2显著提升了叶片总叶绿素含量,有利于叶片光合作用的进行。
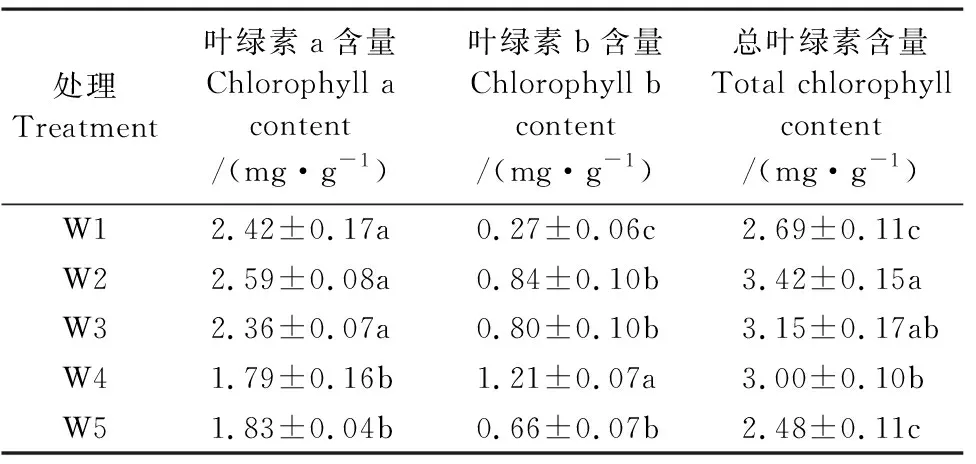
表3 2019年不同土壤基质势对红豆杉叶片叶绿素含量的影响Table 3 Chlorophyll content in leaves of Taxus chinensis under different soil matric potentials in 2019
2.1.4 滴灌土壤基质势调控对红豆杉叶片可溶性蛋白含量的影响 可溶性蛋白作为植物主要的渗透调节物质,在遭受干旱胁迫时,植物会通过渗透调节降低细胞水势和维持细胞膨压,从而使细胞的保水能力增强,防止细胞脱水,进而抵御干旱环境[21]。由图2可知,6月1日时,红豆杉叶片可溶性蛋白含量随土壤基质势的降低呈先升高后降低趋势,W2~W4叶片可溶性蛋白含量均高于W1且与W5差异显著,其中,W3叶片可溶性蛋白含量最高,比W1和W5分别增长了10.36%和14.56%,且差异显著,W5叶片可溶性蛋白含量最低,但此时W5与W1之间无显著差异,表明此时W5叶片仍有较强的渗透调节能力,原因可能是此时试验处理时间较短,低土壤基质势的红豆杉对于水分亏缺做出了应急反应。到8月28日时,红豆杉叶片可溶性蛋白含量随土壤基质势降低所呈的趋势与6月1日相似,但差异性有所不同,W4和W5显著降低了红豆杉叶片可溶性蛋白含量,W2叶片可溶性蛋白含量最高,分别比W1、W3、W4和W5提升了14.35%、11.39%、28.53%、47.31%,且差异显著。表明随着试验时间的延长,低土壤基质势的红豆杉应对水分亏缺的能力降低,W4和W5叶片的渗透调节能力显著下降。
2.1.5 滴灌土壤基质势调控对红豆杉叶片丙二醛含量的影响 MDA是细胞过氧化产物的一种,其含量可以衡量植物细胞活性氧伤害的程度[22]。由图3可知,6月1日时,各处理之间叶片MDA含量无显著差异。至8月28日时,W4和W5与其他处理相比显著提升了红豆杉叶片MDA含量,W5比W1、W2、W3和W4分别增长了30.34%、40.88%、34.32%、5.37%,W4比W1、W2和W3分别增长了23.69%、33.70%、27.47%。表明在6月1日时,各处理红豆杉叶片细胞受活性氧伤害较低,随着试验时间的延长,至8月28日时土壤基质势较低的W4和W5叶片细胞膜脂过氧化加剧,叶片细胞膜受到了一定程度的损伤。
2.2 滴灌土壤基质势调控对红豆杉叶片次生代谢物含量的影响
2.2.1 滴灌土壤基质势调控对红豆杉叶片黄酮含量的影响 由图4可知,2019年8月份各处理叶片黄酮含量与6月份相比呈增大趋势。6月1日时,随着土壤基质势的降低,叶片黄酮含量呈先升高后降低趋势,W3黄酮含量最高,分别比W1、W4和W5增加了23.72%、19.29%、13.61%,且差异显著,比W2增长了2.93%,但差异不显著。到8月28日时,各处理间叶片黄酮含量的变化趋势与6月1日时相似,W3叶片黄酮含量依然最大,分别比W1、W2、W4和W5增长了15.90%、8.99%、4.12%、12.64%,但此时W3只与W1和W5差异显著。2020年11月各处理黄酮含量与9月相比呈降低趋势,且降低幅度较大,但叶片黄酮含量在各处理间的变化规律与2019年基本一致。
2.2.2 滴灌土壤基质势调控对红豆杉叶片多糖含量的影响 由图5可知,2019年6月1日各处理红豆杉叶片多糖含量还未表现出显著差异,8月28日5个处理红豆杉叶片多糖含量随土壤基质势的降低呈先升高后降低趋势,W1和W2叶片多糖含量显著高于W3~W5,W2多糖含量最高,分别比W3、W4和W5显著增加17.40%、25.12%、24.48%,比W1增加7.84%,差异不显著。2020年各处理间叶片多糖含量的变化规律与2019年基本一致,红豆杉叶片多糖含量于试验后期在W2处理下达到最大值。
3 讨 论
土壤水分是影响植物生长发育的重要环境因子,在干旱条件下,植物的呼吸作用、光合作用都会受到一定程度的限制,从而影响植物的生理代谢和生长发育[23]。光响应曲线的拟合能够阐明植物光合作用在各种环境因子下的生态适应性[24],Pnmax、AQE、Lsp、Lcp和Rd等拟合参数, 可反映植物光合能力、生理活性及对弱光利用能力的大小[25],是研究植物生态生理的重要指标。李佳等[26]对米槁幼苗进行不同土壤含水率处理,发现土壤含水率为田间持水量的55%~65%时,米槁幼苗叶片Pn、Pnmax和Lsp达到最大,重度干旱胁迫时,叶片蒸腾速率和气孔导度大幅下降。本试验中,土壤基质势显著影响了红豆杉叶片的光合参数,且低土壤基质势显著降低了红豆杉叶片生理活性和对光强的生态适应性,因此不利于光合作用的进行。而W2显著提升了红豆杉叶片的生理活性,增大了叶片对弱光的利用能力和利用范围,且叶片气孔导度也达到最大值,从而提升叶片Pn和Pnmax,有利于叶片光合作用的进行。
植物叶片中的叶绿素参与光能的吸收、传递和转换等过程,直接影响着植物的光合作用和生长发育[27],叶绿素a和叶绿素b是叶绿素的主要组成成分,叶绿素a以吸收长波光为主,叶绿素b以吸收短波光为主[28],目前关于不同土壤水分条件下植物叶绿素含量的变化报道不一。李英浩等[29]研究发现,随着土壤含水率的降低,燕麦叶片叶绿素含量呈降低趋势。董明等[30]研究发现,阿诺红鞑靼忍冬叶片叶绿素含量在干旱胁迫下呈不断上升趋势。本研究中,随着土壤基质势的降低,红豆杉叶片叶绿素a含量整体呈下降趋势,而叶绿素b含量呈先升高后下降趋势,并在W4达到最大值,表明水分亏缺不利于叶绿素a的合成,而适度水分亏缺则有利于叶绿素b的合成。本试验中,总叶绿素含量在W2时达到最大值,有利于红豆杉光合作用的进行。
植物在顺境环境下,体内的抗氧化酶能够及时地清除不断产生的活性氧,从而使植物能够保持正常的生理活性[31]。但当植物遭受水分胁迫时,植物体内活性氧大量积累,从而引发细胞膜脂过氧化作用,MDA作为膜脂过氧化产物可能和细胞内组分发生相互作用,从而破坏酶和细胞膜的结构,使膜稳定性降低,透性增大,细胞内含物外渗[22],因此,可以通过测定MDA含量间接测定膜系统受损程度,进而判断植物的抗逆性。渗透调节是植物抵抗水分胁迫的一项重要应急反应,可溶性蛋白是重要的渗透调节物质,可以为细胞内的束缚水提供结合衬质,从而减小脱水对细胞结构的破坏[32],本试验中,2019年6月时,W2~W4红豆杉叶片可溶性蛋白含量均高于W1和W5,虽然W5叶片可溶性蛋白含量低于W1,但两者差异不显著,且此时各处理之间MDA含量无显著性差异。8月28日时,W4和W5叶片可溶性蛋白含量显著低于W1~W3处理,且W4和W5叶片MDA含量显著高于其他处理。原因可能是,试验前期低土壤基质势的红豆杉对水分亏缺做出了应急反应,渗透调节能力增强,膜系统损伤降低,细胞的保水性增强,从而使MDA含量降低,各处理叶片MDA含量无显著差异。到试验后期时,较低土壤基质势的W4和W5应对水分亏缺的能力降低,渗透调节能力减弱,细胞膜脂过氧化加剧,MDA含量增加。
黄酮类化合物是植物酚类次生代谢物中的一大类,在植物应对非生物胁迫方面起到了重要作用[33],李丹丹等[34]研究表明,适度干旱胁迫可以提升紫花苜蓿植株中黄酮类化合物含量。张倩倩等[35]研究发现,蒙古黄芪和膜荚黄芪在适度干旱胁迫下,黄芪器官中黄酮化合物含量上升。本试验中,2019―2020两年度内,W3红豆杉叶片黄酮含量均最高,可能是土壤基质势为W3时,红豆杉处于轻微水分亏缺状态,为应对水分亏缺环境,叶片次生代谢增强,黄酮类化合物得到积累,从而增强红豆杉对水分亏缺的防御能力。前人对不同土壤水分条件对植物多糖含量的研究结果不尽相同,梁健萍等[36]研究发现,轻度干旱胁迫(土壤含水量60%~65%)能促进黄芪根中多糖的积累。冯娜等[37]研究发现,在牛膝茎叶生长期和根伸长期将土壤含水量控制在75%和85%时有利于牛膝多糖的积累。贾向荣等[38]研究发现,在田间持水量为75%的适宜水分条件下黄精多糖含量达到最大值。在本试验中,2019―2020两年度红豆杉叶片多糖含量均在试验后期于W2达到最大值,表明W2更有利于红豆杉叶片多糖含量的积累。
本研究筛选出适合红豆杉光合作用和药用成分黄酮和多糖积累的土壤基质势下限,为红豆杉的人工驯化栽培提供了理论依据。但水分对植物生理及品质调控的机理极其复杂,不同土壤基质势下限对红豆杉生理及品质调控的研究尚有大量工作要做。另外,黄酮类和多糖类化合物种类繁多,本试验仅测定了红豆杉叶片总黄酮和总多糖含量,对具体种类的黄酮和多糖化合物的分析尚需深入,且次生代谢过程复杂,受多种酶活性的影响。因此,展开滴灌土壤基质势对红豆杉叶片黄酮和多糖种类、次生代谢关键酶活性的研究是进一步研究的重点。
4 结 论
1)不同土壤基质势对红豆杉叶片的光合作用影响显著,叶片净光合速率和气孔导度均在W2时达到最大值,随着土壤基质势的降低,红豆杉叶片蒸腾速率下降。W2显著提升了红豆杉叶片AQE和Rd,同时显著提高了叶片Lsp和Pnmax,增大了叶片对光的利用范围,且W2叶片叶绿素a和总叶绿素含量最高,因此,将滴灌滴头正下方20 cm处土壤基质势下限设置在-20 kPa有利于红豆杉光合作用的进行。
2)在试验前期时,红豆杉对于适度水分亏缺有一定的抗性,通过应急反应减弱了水分亏缺对细胞造成的伤害。但在试验后期,较低土壤基质势的W4和W5应对水分亏缺的能力下降,可溶性蛋白含量降低,MDA含量增加。
3)红豆杉叶片黄酮含量在W3时达到最大值,多糖含量于试验后期时在W2达到最大值,因此将滴灌滴头正下方20 cm处土壤基质势下限设置在-30 kPa有利于红豆杉叶片黄酮的累积,设置为-20 kPa有利于红豆杉叶片多糖的累积。
