微生物转化粗甘油制备高附加值产品的研究进展
2021-09-15姜莉莉朱宝伟李昌丽刘凤翊
姜莉莉, 朱宝伟, 李昌丽,3, 杨 薇, 刘凤翊*
(1.营口理工学院 化学与环境工程学院,辽宁 营口 115014; 2.江苏省生物质能源与材料重点实验室,江苏 南京 210042; 3.辽宁省化学助剂合成与分离重点实验室,辽宁 营口 115014)
随着石油资源日益枯竭、环境污染严重,生物柴油作为可再生能源越来越受到人们的重视。利用动物或植物油脂与醇类发生酯交换反应生产生物柴油,其副产物甘油占生物柴油总产量的10%[1],粗甘油过剩问题亟待解决。因此,将粗甘油转化为高附加值产品,不仅可以使生物炼制企业获得利润,也利于生物柴油产业的可持续发展[2]。随着甘油价格不断下降,通过微生物发酵法将粗甘油转化为各种生物基化学品受到了人们的重视[3]。甘油是微生物生长的重要底物,能够产生1,3-丙二醇、乙醇、乳酸、二羟基丙酮、氢气、琥珀酸等生物基化学品[3-7]。尽管通过微生物发酵法生产生物基化学品需要面对化学合成法的激烈竞争,但由于其潜在的巨大经济效益,使以粗甘油为碳源的微生物发酵生产绿色化工产品成为研究热点[7]。生物柴油生产过程中的副产物粗甘油常常含有废催化剂、甲醇、皂化物、游离脂肪酸、无机盐等杂质,限制了其在工业生产中的应用[8]。为了满足现有使用的要求,需要对粗甘油进行纯化去除杂质,然而其提纯过程复杂,成本较高,不具有经济可行性。因此,利用微生物发酵法将粗甘油转化为各种高附加值产品是实现可持续发展的重要途径。本文重点介绍了微生物发酵技术转化粗甘油的代谢途径及应用,以期为后续粗甘油经微生物发酵技术转化获得高质量、高附加值化学品提供借鉴和参考。
1 甘油微生物发酵的代谢途径
以甘油为碳源的常见菌属为梭菌属(Clostridium)、克雷伯氏菌属(Klebsiella) 、乳杆菌属(Lactobacillus)、油脂酵母菌属(Lipomyces)、埃希氏菌属(Escherichia)和念珠菌属(Candida)等,这些微生物可以在好氧或厌氧条件下,将甘油转化生产多种生物基化学品[3-7]。同时,基因工程菌也被广泛研究来提高目标产物的产量[9]。肺炎克雷伯氏杆菌(K.pneumoniae)属于兼性厌氧菌,可以在有氧和厌氧条件下分别通过glp和dha两种操纵子代谢甘油,glp操纵子编码的两种酶分别为甘油激酶和甘油-3-磷酸脱氢酶,dha操纵子编码的4种酶分别为甘油脱氢酶(GDH)、二羟丙酮激酶(DHAK)、甘油脱水酶(GDHt)和1,3-丙二醇氧化还原酶(PDOR)。甘油在肺炎克雷伯氏杆菌中的代谢过程(图1)可以侧面反映微生物转化甘油生产高附加值产品的关键酶和主要代谢途径。

NADH2:还原型烟酰胺腺嘌呤二核苷酸reduced nicotinamide adenine dinucleotide;NAD+:氧化型烟酰胺腺嘌呤二核苷酸oxidized nicotinamide adenine dinucleotide;ADP:腺苷二磷酸adenosine diphosphate;ATP:腺苷三磷酸adenosine triphosphate;GDHt:甘油脱水酶glycerol dehydratase;GDH:甘油脱氢酶glycerol dehydrogenase;PDOR:1,3-丙二醇氧化还原酶1,3-propanediol oxidoreductase;DHAK:二羟丙酮激酶dihydroxyacetone kinase
代谢途径包括有氧代谢途径和厌氧代谢途径。有氧代谢途径是指在有氧条件下,进入微生物细胞内的甘油被甘油激酶催化生成甘油-3-磷酸(G3P),G3P会进一步被甘油-3-磷酸脱氢酶转化为二羟基丙酮磷酸(DHAP),然后由磷酸烯醇式丙酮酸生成丙酮酸,再通过乙酰辅酶A进入三羧酸循环(TCA)转化成各种代谢产物(如柠檬酸、琥珀酸、甲酸、苹果酸等),并为微生物提供能量(鸟嘌呤核苷三磷酸(GTP))和还原当量(还原型烟酰胺腺嘌呤二核苷酸(NADH2)和还原型黄素腺嘌呤二核苷酸(FADH2))[10]。厌氧途径是指在厌氧条件下,甘油进入微生物细胞后发生歧化反应,包括氧化和还原两条途径。在还原途径中,甘油首先在甘油脱水酶(GDHt)的催化下生成3-羟基丙醛(3-HPA),然后由NADH2依赖的1,3-丙二醇氧化还原酶(PDOR)还原为1,3-丙二醇。在氧化途径中,甘油在甘油脱氢酶(GDH)的作用下脱氢生成二羟基丙酮,同时生成还原力NADH2。二羟基丙酮进一步被二羟丙酮激酶(DHAK)磷酸化生成二羟基丙酮磷酸并进入糖酵解途径,通过该途径会产生乙醇、乳酸、乙酸、甲酸、2,3-丁二醇等产物,同时产生还原力NADH2和能量用于微生物的生长和还原产物的合成[11]。已有研究表明微量氧气的存在,不仅不会抑制dha操纵子的表达,而且还会在一定程度上提高还原产物1,3-丙二醇的产量,可见氧气对甘油在微生物中的代谢途径具有重要的调节作用[12]。
2 粗甘油的微生物发酵产物
2.1 1,3-丙二醇
1,3-丙二醇(1,3-PD)可以作为单体合成聚酯、聚醚和聚氨酯等多种聚合物,其在纺织、医药、化妆品、涂料、溶剂等领域具有广泛的应用。生物法生产1,3-PD可以避免使用昂贵催化剂和高温高压的条件,以及有毒中间产物的产生,因而微生物转化法生产1,3-PD受到广泛重视。自然界中,常见的用于微生物转化甘油生产1,3-PD 的菌属包括克雷伯氏菌属[13-15]、肠杆菌属(Enterobacter)[16]、柠檬酸杆菌属(Citrobacter)[17]、梭菌属[18-19]和乳杆菌属[20]。
在微生物细胞内甘油通过歧化作用转化为还原产物1,3-PD,以及氧化产物乙醇等醇类和乳酸等有机酸[11]。传统发酵法主要是利用天然菌株或者基因工程菌等微生物通过纯培养法转化纯甘油生产1,3-PD[21]。随着粗甘油产量的逐年增加,为了降低生物法生产1,3-PD的成本,研究者不断尝试利用粗甘油作为底物发酵生产1,3-PD。但是由于粗甘油中的杂质往往会抑制微生物的生长,传统的纯种发酵技术很难高效利用粗甘油,从而造成了底物转化率和产物浓度均较低等缺点。微生物菌群发酵技术由于具有可利用复杂基质、代谢产物可调控、适应性强等优点,促使研究者不断尝试利用微生物菌群发酵代替传统的纯种发酵[22]。不同微生物转化甘油生产1,3-PD的性能比较见表1。

表1 不同微生物转化甘油生产1,3-丙二醇的性能比较
由表1可知,与纯种发酵相比,利用微生物菌群发酵粗甘油生产1,3-PD不仅转化率相当,而且可以在不灭菌的条件下进行操作,大大降低了生产成本。Jiang等[26]从潮间带污泥中筛选可以转化粗甘油生产1,3-PD的微生物菌群CJD-S,通过对发酵条件的优化,在不灭菌条件下进行批次流加粗甘油实验,最终1,3-PD的质量浓度可以达到41.47 g/L,并生成45.86 g/L乳酸,此研究对利用粗甘油生产高附加值生物基化学品具有一定参考价值。
2.2 生物乙醇
生物乙醇因具有能量密度高、易于运输、可再生性好等特点已成为全世界最主要的生物能源之一。生物乙醇生产的主要原料包括:甘蔗、甜菜、甜高粱等含糖作物,玉米、小麦、高粱、木薯、红薯、马铃薯及菊芋等淀粉质作物,以及秸秆、木屑、农作物壳皮及城乡固体垃圾等木质纤维质原料[27]。目前生物乙醇的生产存在与人争粮、生产成本高等问题,为拓展原料来源、保护环境、进一步提高生物乙醇的市场竞争力,利用甘油作为原料来降低生物乙醇的生产成本逐渐得到研究人员的关注[28]。
常用于转化甘油生产乙醇的微生物包括大肠杆菌(Escherichiacoli)、产气肠杆菌(Enterobacteraerogenes)、肺炎克雷伯氏杆菌等(表2)。在微生物中常利用由二羟基丙酮磷酸(DHAP)介导的代谢途径转化甘油生产乙醇:氧化途径由甘油脱氢酶和二羟基丙酮激酶催化,还原途径由甘油-3-磷酸脱氢酶和激酶催化[35]。已有研究表明以粗甘油为原料发酵生产乙醇在成本上会节约40%左右,将大大提高生物乙醇的市场竞争力[36]。在好氧条件下,通过聚冷凝胶固定嗜鞣管囊酵母菌(PachysolentannophilusY-475)生产乙醇,产量可达90%[5]。在厌氧条件下,大肠杆菌(E.coliATCC 11505)能够抵抗甘油抑制作用,耐受高浓度甘油[37]。乙醇的发酵生产还受到碳源、pH值、代谢物、温度等因素的影响,虽然甘油发酵产率高,但存在甘油消耗速度慢、生产强度低等缺点,为此人们利用微生物菌群和基因工程菌等方法来解决此类问题,以期实现工业化生产[29-30,33]。Maru等[38]通过共培养大肠杆菌CECT 432和肠杆菌spH1构建人工菌群来提高微生物利用粗甘油生产乙醇和氢的转化率。有研究将载有醛醇脱氢酶(AdhE)基因的质粒导入肺炎克雷伯氏杆菌BLh-1中,在以粗甘油为底物的批次流加发酵实验中,乙醇的终质量浓度达到了17.30 g/L,生产强度和转化率分别达到了0.59 g/( L·h)-1和16%,与野生菌相比,乙醇的产量增加了11.5 %[39]。Tao等[40]敲除了产酸克雷伯菌作用下甘油氧化代谢途径中乙酸和还原途径中1,3-PD的合成基因后,在以甘油和葡萄糖酸盐为共底物时,乙醇的产率从62%增加到了97.2%。也有研究通过在发酵培养基中添加酪蛋白水解液或金枪鱼冷凝液来提高微生物转化粗甘油生产乙醇的浓度和转化率,进一步降低生物乙醇的生产成本[41-42]。
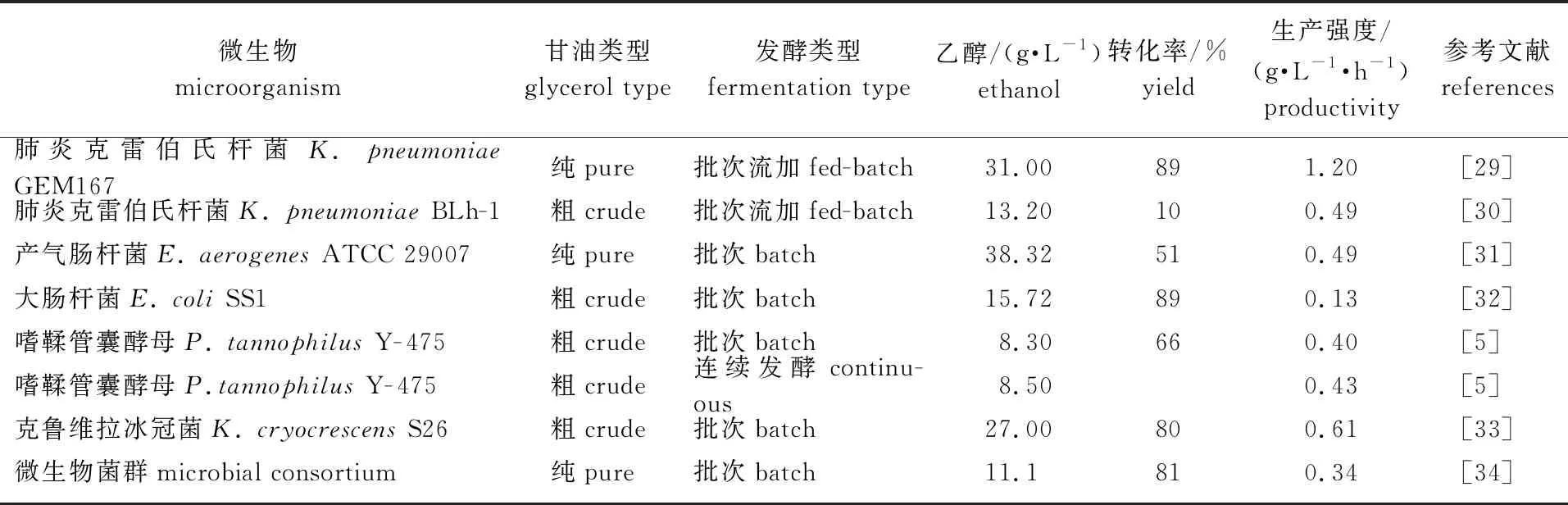
表2 不同微生物转化甘油生产乙醇的性能比较
2.3 乳 酸
乳酸(2-羟基丙酸)是一种含有羧基和羟基的双功能分子,作为一种平台化合物,常用于生产聚乳酸、乙醛、丙烯酸、2,3-乙酰基丙酮等聚合物和化学品,广泛应用于食品、化妆品、制药等行业。乳酸具有手性中心,存在L-乳酸和D-乳酸两种异构体。由于化学法合成的乳酸常常需要高温和高压等条件,并产生外消旋混合物,因此可以生产光学纯乳酸的微生物发酵法一直受到人们的重视[43]。自然界中,克雷伯氏菌属、肠杆菌属、乳杆菌属、肠球菌属(Enterococcus)和根霉菌属(Rhizopus)等微生物能利用甘油产生高浓度的乳酸[8,44-45]。电化学已被应用于微生物转化甘油生产乳酸的发酵过程中,产生的电场通过氧化或还原的方式影响发酵环境和微生物代谢活动,进而提高乳酸的产量和甘油的转化率[46]。Feng等[9]利用基因工程菌肺炎克雷伯氏杆菌(K.pneumoniae)过表达D-乳酸脱氢酶(LdhA),并敲除与1,3-PD 合成相关的两个基因dhaT和yqhD,从而使光学纯D-乳酸达到142.1 g/L,转化率为90%。为了克服粗甘油杂质对纯培养微生物的抑制作用,Wang等[6]从活性污泥中筛选微生物菌群转化粗甘油生产1,3-PD和乳酸。通过间歇发酵实验发现:与单菌相比,微生物菌群发酵表现出优越的底物耐受性和发酵性能,最终在微氧和不灭菌条件下获得27.77 g/L 1,3-PD和14.68 g/L乳酸。Murakami等[47]通过在底物中加入少量的乙酸来提高乳酸的产量,在批次流加发酵实验中于30 g/L甘油和10 g/L乙酸的条件下,L-乳酸的浓度和甘油的转化率分别达到55.3 g/L和0.991 mol/mol。
2.4 1,3-二羟基丙酮
1,3-二羟基丙酮是合成多种精细化学品和生物可降解聚合物的重要前体,具有促进大脑发育、保护视力和预防心脏病等功效,在化妆品、食品、医药等行业有广泛的应用[48]。由于甘油价格低廉,与化学合成法相比微生物发酵法生产1,3-二羟基丙酮更具经济性和安全性[49]。多种天然微生物和基因工程菌可以利用粗甘油生产1,3-二羟基丙酮,包括葡糖杆菌属(Gluconobacter)、葡糖醋杆菌属(Gluconacetobacter)等细菌;裂壶藻属(Schizochytrium)等藻类;毕赤酵母菌属(Pichia)等真菌[50]。葡糖杆菌属是目前生产1,3-二羟基丙酮应用最广泛的微生物,其可以通过膜结合的甘油脱氢酶催化甘油不完全氧化生产1,3-二羟基丙酮。该类微生物在需氧条件下生长,对氧气的需求相对较高。为了提高氧传递效率进而提高1,3-二羟基丙酮的产量,人们深入探究了生物反应器的构型、培养基的理化性质以及菌株等参数。Zhou等[51]利用压缩供氧密封搅拌反应器培养氧化葡萄糖酸杆菌(G.oxydansNL71),在发酵32 h后转化粗甘油生产1,3-二羟基丙酮的质量浓度可达到(301.2±8.2) g/L,生产强度为(9.41±0.23) g/(L·h)。
3 结 语
随着生物柴油产业的迅速发展,其副产物粗甘油已出现供过于求的局面,由于粗甘油中存在多种杂质导致其利用价值较低。基于微生物法具有作用条件温和、绿色环保、可持续发展等特点,因此利用微生物转化粗甘油生产高附加值产品越来越受到人们的重视。由于粗甘油成分复杂,所含杂质常常抑制微生物生长,因此,如何提高微生物对粗甘油的耐受性和转化能力是实现以粗甘油为底物的关键问题。与传统微生物纯种发酵相比,微生物菌群发酵具有可利用复杂原料、代谢产物可调节、不需要严格的无菌环境、适应性强等优点,因此基于微生物菌群的发酵技术有望解决廉价原料难以利用、底物转化率低、副产物多、发酵和分离成本高等产业化难题,为生物基化学品的工业化生产提供了新的技术路线,是今后廉价基质高效利用的研究方向和重点。
