水稻镉胁迫响应机制及防控措施研究进展
2021-09-13许肖博安鹏虎郭天骄韩丹贾玮黄五星
许肖博 安鹏虎 郭天骄 韩丹 贾玮 黄五星
水稻镉胁迫响应机制及防控措施研究进展
许肖博 安鹏虎 郭天骄 韩丹 贾玮 黄五星*
(河南农业大学 烟草学院,郑州 450002;*通信联系人,E-mail: wxhuang@henau.edu.cn)
土壤镉污染是日益严重的环境问题之一。水稻是镉吸收能力最强的大宗谷类作物。水稻对镉的积累及向食物链的转移给人类健康带来威胁。近年来在水稻对镉的吸收和积累过程及其影响因素方面的研究取得了一系列重大进展。本文从镉对水稻种子萌发、生长和发育、产量的影响;土壤中镉的活化、木质部装载运输、韧皮部向籽粒转移三个过程的生理与分子机制以及品种选育、土壤修复、农艺调控等防控措施三个方面进行了综述。以期为水稻镉胁迫相关研究提供理论参考,并为制定降低稻米镉含量的有效策略提供依据。
水稻;镉;生理机制;分子机制;防控措施
镉(Cd)是人体非必需元素,对植物和人类都具有较高的生物毒性[1-2]。镉在人体内易积累不易排出,并且半衰期长,例如在肾脏内的半衰期可达10~30年[3]。低水平的慢性镉摄入会对人体的骨骼和呼吸系统造成危害[4],例如20世纪60年代日本著名的痛痛病事件,就是长期食用镉污染土壤生长的稻米所致[5]。近年来,由于工业“三废”不合理排放、污水灌溉以及含镉农药化肥的不合理使用、土壤酸化等污染问题使得我国土壤中有效镉含量显著增加,导致部分农产品镉含量超标[6]。报告显示我国19.4%耕地土壤污染物含量超标,其中镉以中、低程度污染为主,超标率为7%[7]。
水稻是我国重要的粮食作物,具有富集镉的特性[8],当水稻生长于镉污染土壤中时,镉易在根、茎、叶及籽粒中大量积累,影响水稻的正常生长。稻米是人体镉膳食摄入的主要来源[9],占我国人均每天镉摄入量的55.8%,南方人群占到了65%,水稻品质的优劣直接关系着人类健康[10]。由于土壤污染、高镉积累水稻品种的种植,我国南方一些地区所生产的稻米镉含量超过0.2 mg/kg(GB2762-2017食品中污染物镉限量值)。Zhu等[11]在湖南长沙、株洲、湘潭地区田间调查了近4万份稻米样品,发现稻米镉的平均含量达0.43 mg/kg(0.005~4.80 mg/kg)。Chen等[12]从中国20个省份的当地市场收集到160份精米样本,并对它们的总镉含量进行测定,发现水稻镉含量呈明显的地域分布,南方地区的样本超标率为23%,远高于北部和东部地区(无样品超标)。进一步调查表明采矿是我国水稻土壤镉污染的主要来源,采矿活动中的污染事件会导致严重的镉污染[13]。
因此,控制水稻镉积累,选育低镉积累水稻品种,对人体健康、食品安全生产及环境保护都具有重要的现实意义。本文从镉胁迫对水稻不同生育期的影响。水稻镉吸收转运的生理和分子机质方面的研究进行总结,并概括和展望了调控水稻镉吸收的技术与方法,以期为低镉水稻品种的选育与稻田镉控制提供参考。
1 镉胁迫对水稻的影响
1.1 镉胁迫对水稻种子萌发的影响
种子萌发是植物生命进程的起点,也是植物最早受到重金属胁迫的阶段。镉污染对水稻种子的生长特性指标表现出“低促高抑”的规律。研究证明,高浓度镉胁迫对水稻种子萌发具有明显的抑制作用[14],表现为水稻淀粉酶活性、发芽指数、活力指数的降低,根长下降、根数减少[15]。种子中贮藏的大量淀粉是水稻种子萌发的主要能量来源之一,在淀粉酶的作用下水解成葡萄糖等大分子有机物,输送到正在生长的幼胚中。在高浓度镉胁迫下,淀粉酶的活性受到明显抑制,导致淀粉水解速度下降,胚根和胚轴生长所需要的物质和能量得不到满足,因而胚根和胚轴的伸长受到抑制。与β-淀粉酶相比,α-淀粉酶的活性更易受到镉胁迫的影响[16],这可能与α-淀粉酶中的Ca2+更易被Cd2+所取代[17],从而引起酶空间构型发生改变有关。此外,镉胁迫对种子根长的抑制效应要大于芽[18-19],这与根直接接触镉,以及根部细胞能吸附积累重金属离子有关[20-21]。
与高浓度镉胁迫对种子萌发的影响相比,低浓度镉胁迫对水稻种子萌发影响较小甚至具有促进作用[22]。王泽正等[23]研究发现,单一实验中,低浓度(2、5 mg/L)镉胁迫对水稻种子发芽指数及根长表现为促进作用。此外,不同品种水稻种子对镉的耐受性不同,因而萌发状况也存在较大差异,何俊瑜等[15]对32个水稻品种种子进行了萌发测定,认为可以将参试品种划分为耐受型、中间型和敏感型3种不同类型。
1.2 镉胁迫对水稻幼苗生长及生理生化过程的影响
重金属的毒害作用与作物的生育时期相关,水稻幼苗期是对重金属胁迫最为敏感的时期之一,水稻苗期耐镉能力可以很好地评价不同基因型水稻的耐镉性差异[24]。有学者提出镉胁迫下用最长根长、苗高和单株鲜质量作为评价水稻品种耐镉性的指标,发芽势和发芽率不能作为评价指标[25]。杨冬奇等[26]研究表明镉胁迫下水稻的最长根长、单株鲜质量及苗高均显著低于对照,且随着镉浓度的提高,这种差异也更加显著,其中,镉胁迫对最长根长的抑制效应最为明显,其次是单株鲜质量及苗高。随着镉浓度和胁迫时间的增加,水稻幼苗根长生长受到明显抑制,根尖细胞有丝分裂指数明显下降。此外,镉能诱导染色体产生多种类型的畸变, 染色体畸变率随着镉处理浓度的上升和处理时间的延长而升高,呈现明显的剂量效应和时间效应[27]。
镉胁迫也会对水稻的生理生化过程产生影响。受到镉胁迫时水稻体内产生大量活性氧自由基,这些自由基会攻击细胞膜上的不饱和脂肪酸,从而引起细胞膜结构松散及膜功能的下降[28]。丙二醛(MDA)是植株体内脂类过氧化的最终产物,其含量通常可反映植株遭受氧化胁迫的程度[29],镉胁迫下MDA含量升高会导致膜系统受到损害,引起细胞代谢发生紊乱。镉胁迫也会对植物体内许多酶的活性产生影响,其中,超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)等是植物抗氧化系统中的重要保护酶,在受到镉胁迫时其活性增强,SOD分解超氧自由基为H2O2和O2, CAT分解H2O2产生H2O和O2,而POD以过氧化物为底物[30],以植株体内多种还原剂为电子受体清除过氧化物,从而减轻氧化胁迫对植物的损伤。同时,镉胁迫降低了PSⅡ的活性,影响ATP的形成和光合电子传递,使能量耗散途径受阻,导致活性氧产生从而引起叶绿体结构的破坏和叶片失绿,最终影响其光合作用[31-33]。镉胁迫下不同品种水稻可溶性糖含量响应程度不同,存在品种间的差异。郭文燕等[34]研究表明镉毒害能够促进盐粳7号可溶性糖的合成,却抑制楚粳优1号和南恢511可溶性糖的合成。
1.3 镉胁迫对水稻成熟期的影响
水稻按照形态特征可以分为幼苗期、分蘖期、拔节期、抽穗扬花期和灌浆成熟期五个时期[35]。水稻幼苗期对环境中镉的适应能力较弱,其生长容易受到镉的抑制,随着水稻的生长,在生育后期对镉的适应能力逐渐增强,一定程度上减弱了镉的毒害作用[36]。贾沛菡等[37]利用稳定性同位素示踪、Micro-XRF等技术研究了水稻在不同生育期对镉的吸收与转运特征,结果表明,灌浆成熟期和抽穗期是水稻镉吸收与糙米镉积累的关键时期,对糙米中总镉的贡献率分别达到40.16%和28.61%,采用Micro-XRF技术分析不同节点横截面镉的分布特征,推测这两个时期水稻植株内存在镉的木质部运输与韧皮部再转运的协同作用。
水稻籽粒中镉的积累主要通过两种途径:一种是籽粒灌浆前储存在植株中,灌浆时通过韧皮部随同化物一起从叶片输送到籽粒,另一种是籽粒灌浆时通过木质部随蒸腾流输送到籽粒[38]。喻华等[39]研究认为水稻齐穗后土壤中有效镉含量较低时,两种途径同时存在;水稻齐穗后土壤中有效镉含量较高时,后者是主要途径,即通过根系吸收直接运输到籽粒。程旺大等[40]研究表明,水稻籽粒镉含量和最大灌装速率呈显著正相关,说明镉进入籽粒与同化物的积累是同步的。不同水稻品种的灌浆时间和灌浆特性不同[41],因此籽粒镉积累存在差异。吴启堂等[42]研究则认为,镉向籽粒的迁移率不同是造成糙米镉含量存在品种间差异的主要原因。
1.4 镉胁迫对水稻产量的影响
研究表明,镉毒害能够显著降低水稻的产量、穗数及结实率[28],其降幅呈基因型差异。黄冬芬等[43]研究认为镉胁迫下,水稻产量降低的原因主要在于穗数或每穗颖花数的减少。镉对水稻产量的影响主要在生育前期,通过影响水稻光合、呼吸、矿质元素的吸收等使水稻正常发育受阻,从而导致水稻减产,从减产效应上来说,杂交水稻比常规水稻对镉污染有更高的敏感性[44]。镉胁迫对稻米品质也会产生一定影响,丁园等[45]研究发现当土壤镉含量为5 mg/kg时,水稻籽粒蛋白质含量显著减少,糙米中直链淀粉含量显著增加。
2 水稻对镉吸收、转运与积累的生理机制
水稻根系对镉的吸收是镉进入籽粒的首要环节,然后经木质部装载向地上部转运,并在各组织中转移沉积。水稻籽粒富集镉主要经历四个生理阶段:1)根系对镉的活化与吸收,使其进入根系组织;2)镉在木质部装载,向地上部分转运;3)镉在茎节处向不同组织器官再分配;4)韧皮部再活化转移到水稻籽粒[46]。水稻对镉吸收、转运和积累的特殊生理模式,决定了水稻各部位中镉含量的分布规律是根系>茎叶>籽粒,其中根系中的镉含量是茎叶中的100倍,是糙米中的1000倍[47-48],这是由于水稻根细胞壁与溶解在土壤溶液中的镉直接接触[49],且其根系细胞壁上沉积的镉约占水稻根系中镉总量的45%~90%。
2.1 根系对镉的活化吸收
土壤中的镉进入根系维管柱主要通过两种途径(图1):一种是质外体途径,通过细胞壁、细胞间隙、胞间层和导管空腔组成的结构,利用细胞内外镉离子的浓度差,通过扩散和对流将镉迁移至根系维管柱,属于被动转运,不需要消耗能量。另一种是共质体途径,利用细胞膜上的转运蛋白,先将镉离子转运至根系细胞内,再通过细胞内原生质流动和胞间连丝将其转移至根系维管柱,属于主动吸收,需要消耗能量[50-51]。此外,镉在横向运输到维管柱的过程中,皮层细胞壁上栓质化的凯氏带结构能够阻断质外体通路,导致从皮层来的镉需经过共质体途径,才能到达维管柱的中柱鞘细胞,抑制了镉向地上部的转运[52]。
Zn2+和Fe2+等离子能够影响镉的吸收转运,这些二价阳离子与镉具有相同的核外电子构型和离子半径[53],且其金属转运蛋白底物特异性较低,因此Cd2+能通过占用Mg2+、Zn2+、Mn2+等植物生长必需元素的离子通道进入植物体内。有研究指出Zn2+和Cd2+既存在拮抗作用,也存在协同吸收机制[54-55],这取决于土壤中离子的形态及含量。液泡的区室化解毒作用是植物重金属耐受性机制的一部分[56]。水稻受到镉胁迫时,以谷胱甘肽(GSH)为前体大量合成植物螯合肽(PC),将大部分镉以结合态(PC-Cd)的形式贮存于根部[57],降低了游离态的镉在细胞质中的浓度,是阻止镉长距离运输的有效途径。
2.2 木质部对镉的装载与运输
Uraguchi等[58-59]探究了不同基因型水稻镉积累差异的生理特性与转运机制,认为木质部的装载运输过程,对水稻根系、茎鞘和籽粒镉的积累起决定作用。Cd2+在水稻木质部的装载运输是一个相对独立的过程[52],可分为从木质部薄壁细胞转移到导管和在导管中运输两个阶段,前者借助液泡膜上的转运蛋白将Cd2+装载到木质部导管,后者借助蒸腾作用和根压将Cd2+转移到地上部[60]。高镉积累的水稻品种(如Cho-Ko-Koku)在木质部中镉含量及在导管中的转移速率均比低镉积累水稻品种(如日本晴)要高[61],更能有效向地上部输送镉。
有资料显示,木质部薄壁细胞膜上H-ATPase产生的负性跨膜电势是木质部装载过程的能量来源,木质部细胞壁的阳离子交换量较高,严重阻碍了重金属离子的向上运输,但形成非离子态的金属螯合物后,在蒸腾流中的运输效率得到提升[62]。
2.3 韧皮部对镉向水稻籽粒的转移
茎节是定向调控镉由韧皮部运输到水稻籽粒的重要部位,同位素(107Cd和109Cd)示踪表明[63-64],根系吸收的镉到达木质部后,随着木质部汁液进入茎节,在抽穗前,一部分镉优先积累到稻穗,另一部分则转运到水稻叶片。在叶片解毒机制作用下,水稻叶片储存的镉在重新活化后经穗颈韧皮部累积到水稻籽粒[65]。Kato等[66]研究表明,水稻韧皮部汁液中含有镉螯合剂,镉可能与一种新的13 kDa蛋白和巯基化合物结合。穗颈和韧皮部是镉进入籽粒的路径,穗颈和韧皮部的镉含量与籽粒中镉含量相关,韧皮部的再转移对籽粒中镉含量起重要作用。
3 水稻对镉吸收、转运和积累的分子机制
随着分子生物学的发展,水稻全基因组测序工作已经完成,参与水稻镉吸收转运的多个相关功能基因相继被克隆和研究。根据上述三个生理过程,以下简要分述一些主要基因的功能与特点。
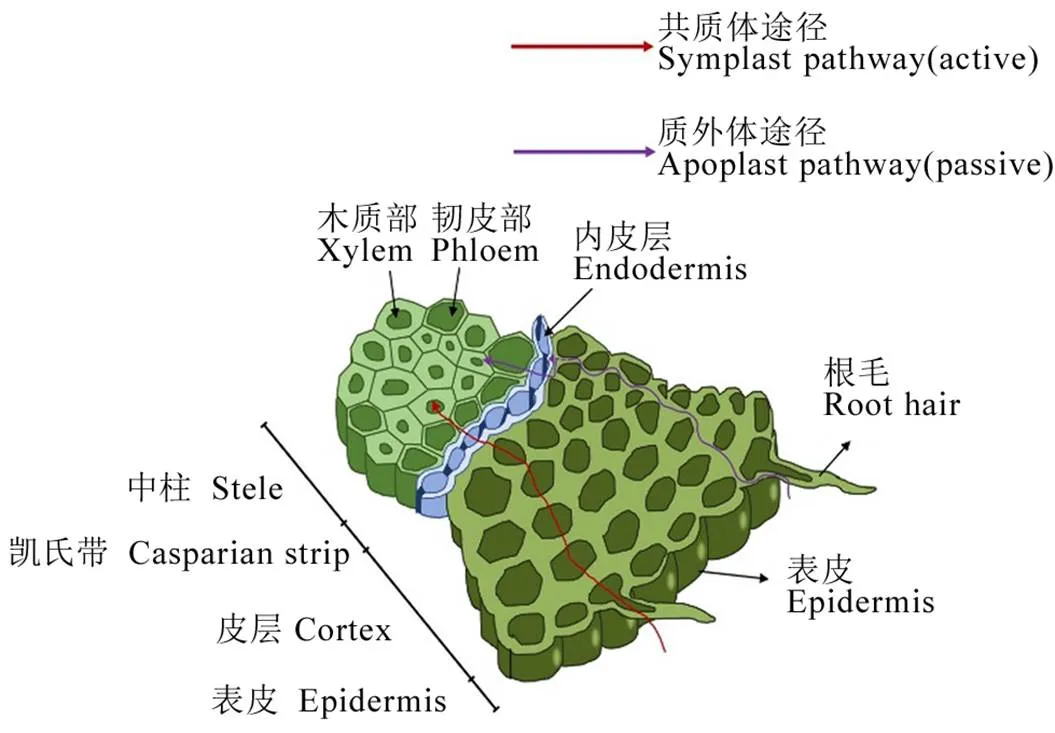
图1 水稻根部镉吸收和转运示意图(参考Li等[51])
Fig.1.Cadmium uptake and transport in rice roots[51].
3.1 根系对镉活化吸收相关的分子机制
镉是植物生长的非必需元素,水稻中不存在镉特异性转运蛋白,镉在水稻根部内外皮层间的运输主要借助其他离子通道蛋白的介导。和是水稻中控制吸收铁元素的主要转运蛋白基因,分别编码亚铁离子转运蛋白和铁调控转运蛋白,和的酵母突变体具有镉离子转运活性,但实验表明对水稻镉的摄取能力低于OsNRAMP5,可能并非是影响水稻镉吸收的主要转运蛋白基因[67]。目前普遍认为,镉进入水稻根系主要借助锰铁离子通道,在根中,主要在根表皮、外皮层、皮层外层以及木质部周围组织中表达,其编码的蛋白定位于细胞质膜上,突变体表现为丧失大部分镉的吸收能力,籽粒中镉含量比野生型显著降低[68-69],然而锰是植物必需矿质营养元素,的敲除或丧失增加了水稻在低锰环境中生长不良的风险,这可能会限制其在水稻镉控制方面的实际应用[70]。
3.2 木质部对镉装载与运输相关的分子机制
木质部介导的镉从根向地上部的转运能力是决定水稻茎秆和籽粒镉积累量的关键[59]。镉离子进入植物根系后,通常被区隔化到液泡中,这一过程由定位于水稻根部液泡膜上的镉转运蛋白OsHMA3主导,能够编码P-1B型重金属ATP合成酶[92],该基因功能丧失或在的T-DNA插入突变体,将导致镉不能被转运到液泡,镉由根系向地上部运输效率大幅提高,从而引起秸秆和籽粒中镉含量升高[93]。相反,过表达能够将镉截留在根部,从而显著降低水稻籽粒的镉积累[92,94],为低镉籽粒水稻的培育提供了新的有效途径。
3.3 韧皮部对镉向水稻籽粒转移相关的分子机制
有研究表明,水稻籽粒中镉含量与水稻基因型有关[95],地上部的镉由韧皮部转移至各个组织器官中。OsLCT1为小麦低亲和性阳离子转运蛋白(Low affinity cation transporter, LCT)的同系物,主要在大的维管束细胞周围以及分散的维管束中表达,定位于细胞质膜上的OsLCT1转运蛋白能够介导镉等阳离子的转运[96]。RNAi株系中,敲除不影响木质部介导的镉转运,但韧皮部介导的镉转运显著减少,敲除株系水稻籽粒中镉含量仅是对照组的1/2[85]。
水稻的遗传基础解析在近两年取得了重大进展。Luo等[84]对叶片镉含量相关QTL位点进行精细定位,克隆到1个能够特异调控镉在水稻叶片中积累的基因(Cd Accumulation in Leaf 1)。主要在根、叶鞘和节中表达,在叶片中基本不表达,编码的类植物防御素蛋白(defensin-like protein)定位于细胞壁上[97],具有胞外信号肽的特点,可通过3个巯基特异地与镉螯合,随后这些螯合态的镉从木质部薄壁细胞的细胞质中分泌出来,经木质部长距离运输最终在叶片中大量积累。可能镉与CAL1的紧密结合使其难以再次跨膜,因此CAL1在促进镉向叶片积累的同时,并未参与韧皮部向籽粒的转运。
4 减少水稻籽粒镉积累的技术措施
4.1 镉污染土壤修复技术
土壤镉污染修复方法众多,最常用的治理方法包括物理修复、化学修复和生物修复。污灌水田试验表明,土壤中的镉主要分布在0-20 cm耕作层中,向下则显著减少,因此可以采用物理修复方法将表层含镉高的土壤移走或采用客土覆盖的形式[98],降低植物对土壤中镉的吸收,但这种方法工作量大,投入成本较高。
化学修复方法主要是指向镉污染土壤施用改良剂、螯合剂等。土壤中的pH和Eh是影响水稻镉吸收的重要环境因子,在碱性和还原性(低Eh值)条件下,土壤表面的负电荷增加,促进了土壤对镉的吸附[52],pH值的升高促使了土壤中的Cd2+与SO42−、S2−及碳酸盐等离子发生共沉淀,降低了土壤镉的迁移性和生物有效性,从而抑制了水稻对镉的吸收。在酸性土壤环境施用磷灰石、农用石灰、生物炭等,对水稻镉的吸收有一定抑制作用。Rehman等[99]在镉污染农田利用石膏改良剂,显著降低了水稻秸秆中镉的浓度和吸收量,提高了水稻的产量和生物量。
生物修复被认为是最有应用前景的土壤修复技术,目前主要是指植物修复和微生物修复。修复镉污染土壤的植物主要包括豆科植物、禾本科植物、藜科植物、十字花科等植物类型[100]。利用根系过滤和生物吸附等,将重金属转移至植物体或降低重金属生物有效性,以此达到修复的目的,但对富集植物的后期处理是限制其发展的技术难题。修复镉污染土壤的微生物主要包括细菌、真菌、藻类、放线菌等,通过其代谢活动影响植物有机酸等物质的分泌,改变植物根际的pH,使重金属物理或化学特性发生改变,以此降低镉在土壤中的迁移和转化[52, 101]。Lin等[102]分离鉴定了3株耐镉细菌,实验表明无论是单一菌株还是组合菌株,都能显著减少镉污染土壤中水稻籽粒中镉的积累,显示出对镉污染土壤进行生物修复的潜力。
4.2 农艺调控措施
农艺措施也是调控水稻镉吸收的重要手段,适量有机肥与化肥配施不仅能维持水稻体内微量元素平衡,而且能够有效降低对镉的吸收。Lin等[103]研究表明硅肥能减轻镉对水稻的毒害,这可能与硅抑制镉的吸收和转运、增强水稻抗氧化酶活性等有关。磷可以在生理上调节水稻对镉的积累和敏感性,Yang等[104]利用水培实验表明,缺磷处理显著降低了水稻幼苗中镉的积累,但加重了镉的毒害症状,表现为分蘖数、根长、苗高和干质量减少。缺磷通过抑制生物量积累,减少碳同化,提高了水稻对镉的敏感性。铁是镉的竞争性金属元素,缺磷通过竞争性地增加铁的吸收和积累来减少水稻对镉的吸收。此外,多种有机酸、多羧基氨基酸螯合剂、Ca2+、 Mg2+、Cu2+、Zn2+、Mn2+等均能抑制植物对Cd2+的吸收和积累[105-106],但随着土壤镉浓度的增加,水稻根系对K+、Zn2+、Mn2+等元素吸收量也随之减少,这些金属元素与镉吸收、转运存在消长关系。硒对镉既存在拮抗作用,也存在协同吸收作用,这取决于硒和镉的相对剂量。Feng等[107]研究表明低镉(≤35.6 μmol/L)处理水平下,硒(≤12.7 μmol/L)能抑制水稻对Cd的吸收,提高水稻生物量,高镉(89~178 μmol/L)水平下,加入硒使得水稻对镉的吸收增加。此外,采用间作或轮作、适当田间淹水管理等方式,也能够减少稻米镉的积累。
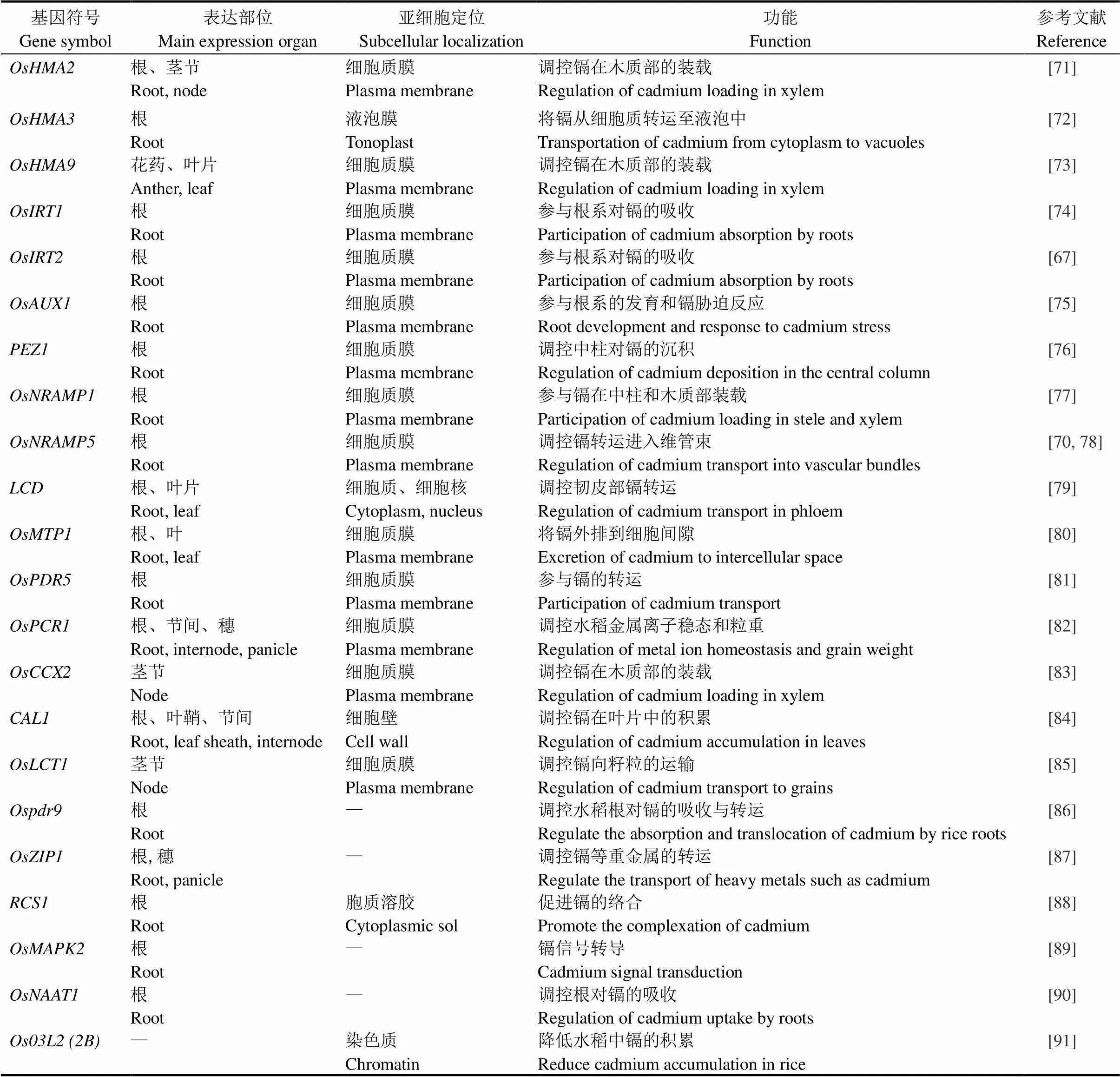
表1 水稻中镉吸收转运相关的基因
4.3 育种调控措施
土壤修复和农艺措施在生产实践中具有一定的可操作性,但往往存在投入成本高、收效低、劳动量大及后续污染等问题。利用分子生物学和遗传方法实现育种调控,培育出低镉水稻品种,从源头上控制稻米的镉含量,实现“边生产边修复”,逐渐成为污染农田修复研究的热点。
利用基因技术,实现降低水稻体内镉积累的研究已有大量报道。水稻中的镉主要通过定位于细胞质膜上的OsNRAMP5进行吸收,利用碳离子束诱变方法获得的OsNRAMP5突变体[69],丧失了大部分镉的吸收能力,其秸秆和籽粒中镉的含量均显著低于野生型。Tang等[108]通过基因编辑技术靶向突变华占和隆科638S,同时敲除杂交稻双亲中的基因,获得的杂交种籽粒镉含量较亲本降低了98%,但OsNRAMP5也是植物必需矿质营养元素锰的转运蛋白[68, 78, 109-110],参与锰从根向地上部的吸收转运[70],的敲除会影响对锰的吸收转运,导致水稻产量下降。过表达可以将镉截留在根部液泡中,从而降低水稻籽粒中镉积累[93]。Luo等[84]通过多代回交和转基因验证,最终克隆了一个正向调控水稻叶片镉含量的基因,编码的一个植物防御素类似蛋白,能够与镉特异螯合,这些螯合态的镉经木质部长距离运输,最终在叶片中大量积累,而不向水稻籽粒积累,因此CAL1影响了韧皮部向水稻籽粒的再分配过程。合理利用和改造这类基因,为培育修复型水稻品种、污染农田的修复提供了新的思路和方向。
5 展望
镉暴露会对生物体和人类造成一系列的不良影响,了解镉在水稻中的运转机制和影响镉积累的因素,有利于制定有效的降镉策略,减轻对生物体和人类的危害。近年来,人们对镉吸收和转运机制的认识取得了很大进展,在水稻中发现了一些镉转运蛋白家族,如水稻自然抗性相关巨噬细胞蛋白(natural resistance-associated macrophage protein,NRAMP)家族。一些调节水稻镉吸收积累的转运蛋白具有广泛的底物范围,除镉元素外,也能够转运与水稻生长发育所必需的微量元素(如锰、铁等),敲除相关基因是否会对水稻必需金属元素的获取造成负面影响,是水稻改良过程中的一个重要问题[109]。已知的重金属离子转运蛋白并不能完全解析水稻在镉吸收转运的动态变化过程,因此基于分子生物学技术对未知转运蛋白或其他分子进行识别是非常必要的。土壤理化性质与生态生理对镉的生物活性有重要影响,因地制宜选择实用的生物修复方法,筛选适宜的微生物,作为可行的方法已被广泛研究。此外,利用基因工程技术进行稻米镉积累的QTL定位和转录组分析,可以提高对镉胁迫下水稻分子反应机制的认识,有利于高产低镉水稻品种的筛选。因此,未来的研究可以集中在以下3个方面:
1)识别与验证水稻中与镉转运或耐性有关的蛋白。水稻对不利环境的高度适应性意味着存在高效的抗性机制,包括产生抗菌肽(AMPs)等。AMPs可参与植物对非生物胁迫的反应,如高盐度、干旱、高温或低温以及重金属[110]。在植物AMPs中,防御素是研究最多的家族之一[111],如基因编码的植物防御素类似蛋白,其蛋白上的3个巯基可以特异地与镉进行螯合而不与钙、锰和锌等二价离子结合,促进镉在水稻叶片中的积累而不参与向籽粒的运输。禾本科植物防御素的研究取得了相当大的进展,但只有极少数的DEFL基因库中的肽进行了功能鉴定,此外,不同防御素的分子机制仍有待研究,防御素和禾本科植物对环境胁迫的高适应性,将为农业的实际应用提供新的思路与方向。有研究表明,ATP结合盒(ABC)转运蛋白可能在植物对镉的耐受性中起重要作用[112],该蛋白定位于根细胞的质膜上。Fu等[113]对一个ABCG型转运蛋白OsABCG36进行了功能验证,结果表明,OsABCG36通过将镉运出根细胞而参与水稻对镉的耐受。水稻中有120多种ABC转运蛋白,然而,大多数ABC转运蛋白在植物中的确切功能尚不清楚,有待进一步验证。
2)生态修复措施。镉污染的修复方法众多,但效果大多难以达到预期,微生物修复技术因其环境友好和成本效益高而越来越受到人们的关注。铜绿假单胞菌是一种多金属耐受菌,对重金属有很高的耐受性[114],特别是镉(2200 mg/L)、锌(1800 mg/L)和铅(1200 mg/L)。盆栽实验表明,该菌使糙米和精米中镉的累积量分别减少了31.2%和25.5%,有效减少了水稻籽粒中镉的积累[102]。从镉污染土壤中分离和鉴定耐镉细菌,并研究耐镉细菌(单一菌株和混合菌株)减少水稻籽粒中镉积累的能力,综合评估菌株在稻田镉污染土壤生物修复中的潜力,是未来水稻镉污染防治的重要方向。此外,不同区域稻田生态特性存在差异,应对稻田土壤理化性质进行系统研究,因地制宜采取磷、钾等肥料配比,施用改良剂等,系统调控水稻镉积累。
3)通过基因工程选育高产低镉积累品种。探索镉胁迫下的遗传和分子机制有助于低镉积累水稻品种的开发,RNA-Seq是最近开发的转录组分析方法,RNA-Seq平台的转录组分析阐明了水稻的镉胁迫反应。Sun等[115]应用RNA-Seq的方法检测出镉胁迫下水稻幼苗中2197个差异表达基因(987个上调,1210个下调),其中编码生长素反应蛋白IAA的基因被上调,编码参与信号转导的蛋白质的基因如TIFY家族被下调,编码过氧化物酶的基因在次生代谢产物的生物合成过程中,有些表达上调,有些表达下调,表明过氧化物酶在水稻对镉胁迫的反应中起着关键作用。这一结果为水稻幼苗镉胁迫分子机制的进一步研究提供了依据,也能为高产低镉水稻品种的选育奠定基础。
[1] 郑陶, 李廷轩, 张锡洲, 余海英, 王勇.水稻镉高积累品种对镉的富集特性[J].中国农业科学, 2013, 46(7): 1492-1500.
Zheng T, Li T X, Zhang X Z, Yu H Y, Wang Y.Accumulation characteristics of cadmium-accumulated rice cultivars with high cadmium accumulation[J]., 2013, 46(7): 1492-1500.(in Chinese with English abstract)
[2] He S, He Z, Yang X, Stoffella P J, Baligar V C.Soil biogeochemistry, plant physiology, and phytoremediation of cadmium-contaminated soils[J]., 2015, 134: 135-225.
[3] Phuc H D, Kido T, Oanh N T P, Manh H D, Anh L T, Oyama Y, Okamoto R, Ichimori A, Nogawa K, Suwazono Y, Nakagawa H.Effects of aging on cadmium concentrations and renal dysfunction in inhabitants in cadmium-polluted regions in Japan[J]., 2017, 37(9): 1046-1052.
[4] Nawrot T S, Staessen J A, Roels H A, Munters E, Cuypers A, Richart T, Ruttens A, Smeets K, Clijsters H, Vangronsveld J.Cadmium exposure in the population: From health risks to strategies of prevention[J]., 2010, 23(5): 769-782.
[5] Grant C A, Clarke J M, Duguid S, Chaney R L.Selection and breeding of plant cultivars to minimize cadmium accumulation.[J]., 2008, 390(2-3): 301-310.
[6] Clemens S, Aarts M G M, Thomine S, Verbruggen N.Plant science: The key to preventing slow cadmium poisoning[J]., 2013, 18(2): 92-99.
[7] 环境保护部, 国土资源部.全国土壤污染状况调查公报[EB/OL].(2014-04-17)[2021-03-22].http://www.mee.gov.cn/gkml/sthjbgw/qt/201404/W020140417558995804588.pdf
Ministry of Environmental Protection, Ministry of Land and Resources, the People’s Republic of China.Bulletin of the National Soil Pollution Survey [EB/OL].(2014-04- 17)[2021-03-22].http://www.mee.gov.cn/gkml/sthjbgw/ qt/201404/W020140417558995804588.pdf (in Chinese)
[8] Chaney R L, Reeves P G, Ryan J A, Simmons R W, Welch R M, Scott Angle J.An improved understanding of soil Cd risk to humans and low cost methods to phytoextract Cd from contaminated soils to prevent soil Cd risks[J]., 2004, 17(5): 549-553.
[9] Sheng Z M, Lin J, Dan H, Xiang X T, Jun Y J, Yang J X, Chao P.Cadmium exposure via diet and its implication on the derivation of health-based soil screening values in China[J]., 2015, 25(4): 433-442.
[10] Song Y, Wang Y, Mao W F, Sui H X, Yong L, Yang D J, Jiang D G, Zhang L, Gong Y Y.Dietary cadmium exposure assessment among the Chinese population[J]., 2017, 12(5): e0177978.
[11] Zhu H H, Chen C, Xu C, Zhu Q H, Huang D Y.Effects of soil acidification and liming on the phytoavailability of cadmium in paddy soils of central subtropical China[J]., 2016, 219: 99-106.
[12] Chen H P, Tang Z, Wang P, Zhao F J.Geographical variations of cadmium and arsenic concentrations and arsenic speciation in Chinese rice[J]., 2018, 238: 482-490.
[13] Liu X J, Tian G J, Jiang D, Zhang C, Kong L Q.Cadmium (Cd) distribution and contamination in Chinese paddy soils on national scale[J]., 2016, 23(18): 17941-17952.
[14] 孙亚莉.水稻种子萌发与幼苗期对镉胁迫的响应差异研究[D].长沙: 湖南农业大学, 2018.
Sun Y L.Studies on the difference of response to cadmium stress in rice seed germination and seedling stage[D].Changsha: Hunan Agricultural University, 2018.(in Chinese with English abstract)
[15] 何俊瑜, 任艳芳, 周国强, 杨良静, 王阳阳.不同水稻品种种子萌发期耐镉性的研究[J].华北农学报, 2010, 25(3): 108-112.
He J Y, Ren Y F, Zhou G Q, Yang L J, Wang Y Y.Study on cadmium tolerance of different varieties ofL.during seed germination stage [J]., 2010, 25(3): 108-112.(in Chinese with English abstract)
[16] 何俊瑜, 任艳芳, 朱诚, 蒋德安.镉胁迫对不同水稻品种种子萌发、幼苗生长和淀粉酶活性的影响[J].中国水稻科学, 2008(4): 399-404.
He J Y, Ren Y F, Zhu C, Jiang D A.Effects of cadmium stress on seed germination, seedling growth, and amylase activities in rice [J]., 2008 (4): 399-404.(in Chinese with English abstract)
[17] Mikami B, Adachi M, Kage T, Sarikaya E, Nanmori T, Shinke R, Utsumi S.Structure of raw starch-digestingbeta-amylase complexed with maltose[J]., 1999, 38(22): 7050-7061.
[18] 宋玉芳, 许华夏, 任丽萍, 龚平, 周启星.土壤重金属对白菜种子发芽与根伸长抑制的生态毒性效应[J].环境科学, 2002(1): 103-107.
Song Y F, Xu H X, Ren L P, Gong P, Zhou Q X.Eco-toxicological effects of heavy metals on the inhibition of seed germination and root elongation of Chinese cabbages in soils[J]., 2002(1): 103-107.(in Chinese with English abstract)
[19] 李伟强, 毛任钊, 刘小京.胁迫时间与非毒性离子对重金属抑制拟南芥种子发芽及幼苗生长的影响[J].应用生态学报, 2005(10): 1943-1947.
Li W Q, Mao R Z, Liu X J.Effects of stress duration and non-toxic ions on heavy metals toxicity toseed germination and seeding groth[J]., 2005(10): 1943-1947.(in Chinese with English abstract)
[20] 檀建新, 尹君, 王文忠, 贾荣革, 史吉平.镉对小麦、玉米幼苗生长和生理生化反应的影响[J].河北农业大学学报, 1994(S1): 83-87.
Tan J X, Yin J, Wang W Z, Jia R G, Shi J P.Effects of cadmium on growth and physiological and biochemical responses of wheat and maize seedlings[J]., 1994 (S1): 83-87.(in Chinese with English abstract)
[21] 王春春, 沈振国.镉在植物体内的积累及其对绿豆幼苗生长的影响[J].南京农业大学学报, 2001(4): 9-13.
Wang C C, Shen Z G.Uptake of Cd by three species of plants and responses of mung bean to Cd toxicity[J].,2001(4): 9-13.(in Chinese with English abstract)
[22] 何俊瑜, 任艳芳.镉胁迫对水稻种子萌发、幼苗生长和淀粉酶活性的影响[J].华北农学报, 2008(S1): 131-134.
He J Y, Ren Y F.Effect of Cadmium on seed germination, seedling growth and amylolytic activity of rice[J]., 2008(S1): 131-134.(in Chinese with English abstract)
[23] 王泽正, 杨亮, 李婕, 付东东, 胡维薇, 范正权, 彭丽成.微塑料和镉及其复合对水稻种子萌发的影响[J].农业环境科学学报,2021, 40(1): 44-53.
Wang Z Z, Yang L, Li J, Fu D D, Hu W W, Fan Z Q, Peng L C.Single and combined effects of microplastics and cadmium on the germination characteristics of rice seeds[J]., 2021, 40(1): 44-53.(in Chinese with English abstract)
[24] 程旺大, 姚海根, 张国平, 汤美玲, Dominy P.镉胁迫对水稻生长和营养代谢的影响[J].中国农业科学, 2005(3): 528-537.
Cheng W D, Yao H G, Zhang G P, Dominy P.Effects of cadmium stress on growth and nutrient metabolism of rice[J]., 2005 (3): 528-537.(in Chinese with English abstract)
[25] 陈志德, 仲维功, 王军, 杨杰, 张红生.镉胁迫对水稻种子萌发和幼苗生长的影响[J].江苏农业学报, 2009, 25(1): 19-23.
Chen Z D, Zhong W G, Wang J, Yang J, Zhang H S.Effects of cadmium stress on seed germination and seedling growth of various rice varieties[J]., 2009, 25(1): 19-23.(in Chinese with English abstract)
[26] Yang D Q, Liu S X, Xia S P, Peng G X, Huang F L.Effects of cadmium stress on the growth of rice seedlings [J]., 2019, 20(3): 11-16.
[27] 何俊瑜, 任艳芳, 严玉萍, 朱诚, 蒋德安.镉胁迫对水稻幼苗生长和根尖细胞分裂的影响[J].土壤学报, 2010, 47(1): 138-144.
He J Y, Ren Y F, Yan Y P, Zhu C, Jiang D A.Impacts of cadmium stress on the growth of rice seedlings and division of their root tip cells[J]., 2010, 47 (1): 138-144.(in Chinese with English abstract)
[28] 陈京都.水稻镉胁迫响应差异机理和调控效应的研究[D].扬州: 扬州大学, 2013.
Chen J D.Research on difference and mechanism of rice response under cadmium stress and regulation[D].Yangzhou: Yangzhou University, 2013.(in Chinese with English abstract)
[29] 王昌全, 郭燕梅, 李冰, 袁大刚, 张济龙, 林正雨, 唐敦义.Cd胁迫对杂交水稻及其亲本叶片丙二醛含量的影响[J].生态学报, 2008(11): 5377-5384.
Wang C Q, Guo Y M, Li B, Yuan D G, Zhang J L, Lin Z Y, Tang D Y.Effects of Cd stress on the content of MDA in leaves of the hybrid rice and their parents[J]., 2008(11): 5377-5384.(in Chinese with English abstract)
[30] 邵国胜, Hassan M J, 章秀福, 张国平.镉胁迫对不同水稻基因型植株生长和抗氧化酶系统的影响[J].中国水稻科学,2004(3): 57-62.
Shao G S, Hassan M J, Zhang X F, Zhang G P.Effects of cadmium stress on plant growth and antioxidant enzyme system in different rice genotypes[J]., 2004(3): 57-62.(in Chinese with English abstract)
[31] 徐红霞, 翁晓燕, 毛伟华, 杨勇.镉胁迫对水稻光合、叶绿素荧光特性和能量分配的影响[J].中国水稻科学, 2005(4): 338-342.
Xu H X, Weng X Y, Mao W H, Yang Y.Effects of cadmium stress on photosynthesis, chlorophyll fluo- rescence characteristics and excitation energy distribution in leaves of rice[J]., 2005(4): 338-342.(in Chinese with English abstract)
[32] 葛才林, 骆剑峰, 刘冲, 殷朝珍, 王泽港, 马飞, 罗时石.重金属对水稻光合作用和同化物输配的影响[J].核农学报, 2005(3): 214-218.
Ge C L, Luo J F, Liu C, Yin C Z, Wang Z G, Ma F, Luo S S.Effect of heavy metals on the photosynthesis and photosynthates transformation in rice[J]., 2005(3): 214-218.(in Chinese with English abstract)
[33] 王锦文, 边才苗, 陈珍.铅、镉胁迫对水稻种子萌发、幼苗生长及生理指标的影响[J].江苏农业科学, 2009(4): 77-79.
Wang J W, Bian C M, Chen Z.Effects of lead and cadmium stress on seed germination, seedling growth and physiological indexes of rice[J]., 2009(4): 77-79.(in Chinese with English abstract)
[34] 郭文燕, 田雄, 李尚锟, 李伟, 黄永相, 胡燕, 赵夏夏, 郭建夫.镉胁迫对抽穗期水稻生理生化特性的影响[J].安徽农业科学, 2018, 46(14): 37-43.
Guo W Y, Tian X, Li S K, Li W, Huang Y X, Hu Y, Zhao X X, Guo G F.Effects of cadmium stress on the physiological and biochemical characteristics of rice at heading stage[J]., 2018, 46 (14): 37-43.(in Chinese with English abstract)
[35] 胡雨丹, 周航, 辜娇峰, 霍洋, 邓鹏辉, 魏宾纭, 刘俊, 廖柏寒.水培试验下水稻Pb吸收累积关键生育期[J].环境科学, 2020, 41(9): 4218-4225.
Hu Y D, Zhou H, Gu J F, Huo Y, Deng P H, Wei B Y, Liu J, Liao B H.Key growth stage of Pb accumulation in rice through a hydroponic experiment with Pb stress[J]., 2020, 41(9): 4218-4225.(in Chinese with English abstract)
[36] 黄冬芬, 奚岭林, 杨立年, 王志琴, 杨建昌.不同耐镉基因型水稻农艺和生理性状的比较研究[J].作物学报, 2008(5): 809-817.
Huang D F, Xi L L, Yang L N, Wang Z Q, Yang J C.Comparisons in agronomic and physiological traits of rice genotypes differing in cadmium-tolerance[J]., 2008(5): 809-817.(in Chinese with English abstract)
[37] 贾沛菡.水稻对镉的吸收受不同介质条件与生育期的影响及其与籽粒积累镉的关系[D].杭州: 浙江大学, 2019.
Jia P H.The factors that affecting Cd uptake and its relationship with Cd accumulation in grains of rice[D].Hangzhou: Zhejiang University, 2019.(in Chinese with English abstract)
[38] Maria G,Martina L.Comparison of uptake and distribution of cadmium in different cultivars of bread and durum wheat[J]., 2004, 44(2): 501- 507.
[39] 喻华, 上官宇先, 涂仕华, 秦鱼生, 陈琨, 陈道全, 刘前聪.水稻籽粒中镉的来源[J].中国农业科学, 2018, 51(10): 1940-1947.
Yu H, Shang G Y X, Tu S H, Qin Y S, Chen K, Chen D Q, Liu Q C.Sources of cadmium accumulated in rice grain[J]., 2018, 51(10): 1940-1947.(in Chinese with English abstract)
[40] 程旺大, 张国平, 姚海根, DOMINY P, 王润屹.晚粳稻籽粒中砷、镉、铬、镍、铅等重金属含量的品种和粒位效应[J].中国水稻科学, 2005, 18(3): 273-279.
Cheng W D, Zhang G P, Yao H G, Dominy P, Wang R Q.Effect of grain position in a panicle and varieties on As, Cd, Cr, Ni, Pb contents in grains of late japonica rice[J]., 2005, 18(3): 273-279.(in Chinese with English abstract)
[41] 陈京都, 何理, 林忠成, 戴其根, 张军, 郭保卫, 许露生, 张洪程.不同生育期类型水稻对镉积累的研究[J].生态与农村环境学报, 2013, 29(3): 390-393.
Chen J D, He L, Lin Z C, Dai Q G, Zhang J, Guo B W, Xu L S, Zhang H C.Cd accumulation in japonica rice relative to growth type[J]., 2013, 29(3): 390-393.(in Chinese with English abstract)
[42] 吴启堂, 陈卢, 王广寿.水稻不同品种对Cd吸收累积的差异和机理研究[J].生态学报,1999(1): 106-109.
Wu Q T, Chen L, Wang G S.Difference and mechanism of Cd uptake and accumulation in different rice varieties[J]., 1999(1): 106-109.(in Chinese with English abstract)
[43] 黄冬芬, 王志琴, 刘立军, 杨建昌.镉对水稻产量和品质的影响[J].热带作物学报, 2010, 31(1): 19-24.
Huang D F, Wang Z Q, Liu L J, Yang J C.Effects of cadmium on the rice yield and grain quality[J]., 2010, 31 (1): 19-24.(in Chinese with English abstract)
[44] 王凯荣, 龚惠群.不同生育期镉胁迫对两种水稻的生长、镉吸收及糙米镉含量的影响[J].生态环境, 2006(6): 1197-1203.
Wang K R, Gong H Q.Effects of cadmium exposures in different stages on plant growth, cd uptake and cd concentrations in brown rice of a hybrid and conventional rice variety [J]., 2006(6): 1197-1203.(in Chinese with English abstract)
[45] 丁园, 宗良纲, 徐晓炎, 刘光荣.镉污染对水稻不同生育期生长和品质的影响[J].生态环境学报, 2009, 18(1): 183-186.
Ding Y, Zong L G, Xu X Y, Liu G R.Effect of cadmium on the growth and quality of rice (L.) In different growth period[J]., 2009, 18(1): 183-186.(in Chinese with English abstract)
[46] Uraguchi S, Fujiwara T.Cadmium transport and tolerance in rice: Perspectives for reducing grain cadmium accumulation[J]., 2012, 5(1): 5.
[47] 赵步洪, 张洪熙, 奚岭林, 朱庆森, 杨建昌.杂交水稻不同器官镉浓度与累积量[J].中国水稻科学, 2006(3): 306-312.
Zhao B H, Zhang H X, Xi L L, Zhu Q S, Yang J C.Cadmium concentration and accumulation in different organs of hybrid rice[J]., 2006, 20 (3): 306-312.(in Chinese with English abstract)
[48] 刘侯俊, 梁吉哲, 韩晓日, 李军, 芦俊俊, 张素静, 冯璐, 马晓明.东北地区不同水稻品种对Cd的累积特性研究[J].农业环境科学学报, 2011, 30(2): 220-227.
Liu H J, Liang J Z, Han X R, Li j, Lu J J, Zhang S J, Feng L, Ma X M.Accumulation and distribution of cadmium in different rice cultivars of Northeastern China[J]., 2011, 30(2): 220-227.(in Chinese with English abstract)
[49] Nishizono H, Ichikawa H, Suziki S, Ishii F.The role of the root cell wall in heavy metal tolerance of[J]., 1987, 101(1): 15-20.
[50] 朱智伟, 陈铭学, 牟仁祥, 曹赵云, 张卫星, 林晓燕.水稻镉代谢与控制研究进展[J].中国农业科学, 2014, 47(18): 3633-3640.
Zhu Z W, Chen M X, Mou R X, Cao Z Y, Zhang W X, Lin X Y.Advances in research of cadmium metabolism and control in rice plants[J]., 2014, 47(18): 3633-3640.(in Chinese with English abstract)
[51] Li H, Luo N, Li Y W, Cai Q Y, Li H Y, Mo C H, Wong M H.Cadmium in rice: Transport mechanisms, influencing factors, and minimizing measures[J]., 2017, 224: 622-630.
[52] 刘利, 郝小花, 田连福, 戴小军, 梁满中, 李东屏, 陈良碧.植物吸收、转运和积累镉的机理研究进展[J].生命科学研究,2015, 19(2): 176-184.
Liu L, Hao X H, Tian L F, Dai X J, Liang M Z, Li D P, Chen B L.Research progresses on the mechanism of Cd absorption, transport and accumulation in plant[J]., 2015, 19(2): 176-184.(in Chinese with English abstract)
[53] 周志波, 易亚科, 陈光辉.水稻Cd吸收、转运机理研究进展[J].作物杂志, 2017(1): 14-19.
Zhou Z B, Yi Y K, Chen G H.Advances in Cd uptake and transport in rice[J]., 2017(1): 14-19.(in Chinese with English abstract)
[54] Tian S K, Lu L L, Zhang J, Wang K, Brown P, He Z L, Liang J, Yang X E.Calcium protects roots ofH.against cadmium-induced oxidative stress[J]., 2011, 84(1): 63-69.
[55] Pence N S, Larsen P B, Ebbs S D, Letham D L, Lasat M M, Garvin D F, Eide D, Kochian L V.The molecular physiology of heavy metal transport in the Zn/Cd hyperaccumulator[J]., 2000, 97(9): 4956-4960.
[56] 孙敏.水稻植株中镉区室化关键螯合物的鉴定与分析[D].北京: 中国农业科学院, 2010.
Sun M.Characterization and analysis of chelates involved in compartmentation of cadmium in rice plants[D].Beijing: Chinese Academy of Agricultural Sciences, 2010.(in Chinese with English abstract)
[57] Guo J B, Xu W Z, Ma M.The assembly of metals chelation by thiols and vacuolar compartmentalization conferred increased tolerance to and accumulation of cadmium and arsenic in transgenic[J]., 2012, 199: 309-313.
[58] Uraguchi S, Fujiwara T.Rice breaks ground for cadmium-free cereals[J]., 2013, 16(3): 328-334.
[59] Uraguchi S, Mori S, Kuramata M, Kawasaki A, Arao T, Ishikawa S.Root-to-shoot Cd translocation via the xylem is the major process determining shoot and grain cadmium accumulation in rice[J]., 2009, 60(9): 2677-2688.
[60] Kobayashi N I, Tanoi K, Hirose A,Nakanishi T M.Characterization of rapid intervascular transport of cadmium in rice stem by radioisotope imaging[J]., 2013, 64(2): 507-517.
[61] Ueno D, Koyama E, Yamaji N, Ma J F.Physiological, genetic, and molecular characterization of a high-Cd- accumulating rice cultivar, Jarjan[J]., 2011, 62(7): 2265-2272.
[62] 杨肖娥, 龙新宪, 倪吾钟.超积累植物吸收重金属的生理及分子机制[J].植物营养与肥料学报, 2002(1): 8-15.
Yang X E, Long X X, Ni W Z.Physiological and molecular mechanisms of heavy metal uptake by hyper- accumulating plants[J]., 2002(1): 8-15.(in Chinese with English abstract)
[63] Fujimaki S, Suzui N, Ishioka N S, Kawachi N, Ito S, Chino M, Nakamura S.Tracing cadmium from culture to spikelet: Noninvasive imaging and quantitative characterization of absorption, transport, and accumulation of cadmium in an intact rice plant[J]., 2010, 152(4): 1796-1806.
[64] Tanaka K, Fujimaki S, Fujiwara T, Yoneyama T, Hayashi H.Quantitative estimation of the contribution of the phloem in cadmium transport to grains in rice plants (L.)[J]., 2007, 53(1): 72-77.
[65] Rodda M S, Li G, Reid R J.The timing of grain Cd accumulation in rice plants: The relative importance of remobilisation within the plant and root Cd uptake post-flowering [J]., 2011, 347(1-2): 105-114.
[66] Kato M, Ishikawa S, Inagaki K, Chiba K, Hayashi H, Yanagisawa S, Yoneyama T.Possible chemical forms of cadmium and varietal differences in cadmium concen- trations in the phloem sap of rice plants(L.)[J]., 2010, 56(6): 839-847.
[67] Nakanishi H, Ogawa I, Ishimaru Y, Mori S, Nishizawa N K.Iron deficiency enhances cadmium uptake and translocation mediated by the Fe2+transporters OsIRT1 and OsIRT2 in rice[J]., 2006, 52(4): 464-469.
[68] Sasaki A, Yamaji N, Yokosho K, Ma J F.Nramp5 is a major transporter responsible for manganese and cadmium uptake in rice[J]., 2012, 24(5): 2155-2167.
[69] Ishikawa S, Ishimaru Y, Igura M, Kuramata M, Abe T, Senoura T, Hase Y, Arao T, Nishizawa N K, Nakanishi H.Ion-beam irradiation, gene identification, and marker- assisted breeding in the development of low-cadmium rice[J]., 2012, 109(47): 19166-19171.
[70] Yang M, Zhang Y Y, Zhang L J, Hu J T, Zhang X, Lu K, Dong H X, Wang D J, Zhao F J, Huang C F, Lian X M.OsNRAMP5 contributes to manganese translocation and distribution in rice shoots[J]., 2014, 65(17): 4849-4861.
[71] Namiko S N, Mikako M, Nobushige N, Tomohiko K, Yasuo N, Kenji S, Hidekazu T, Akio W, Hiromori A.Mutations in rice () heavy metal ATPase 2 (OsHMA2) restrict the translocation of zinc and cadmium[J]., 2012, 53(1): 213-224.
[72] Sasaki A, Yamaji N, Ma J F.Overexpression of OsHMA3 enhances Cd tolerance and expression of Zn transporter genes in rice[J]., 2014, 65(20): 6013-6021.
[73] Zhao F Y, Han M M, Zhang S Y, Wang K, Zhang C R, Liu T, Liu W.Hydrogen peroxide-mediated growth of the root system occurs via auxin signaling modification and variations in the expression of cell-cycle genes in rice seedlings exposed to cadmium stress[J]., 2012, 54(12): 991-1006.
[74] Lee S, An G.Over-expression ofleads to increased iron and zinc accumulations in rice[J]., 2009, 32(4): 408-416.
[75] Yu C L, Sun C D, Shen C J, Wang S K, Liu F, Liu Y, Chen Y L, Li C Y, Qian Q, Bibek A, Markus G, Jiang D A, Qi Y H.The auxin transporter, OsAUX1, is involved in primary root and root hair elongation and in Cd stress responses in rice (L.)[J]., 2015, 83(5): 818-830.
[76] Ishimaru Y, Bashir K, Nakanishi H, Nishizawa N K.The role of rice phenolics efflux transporter in solubilizing apoplasmic iron[J]., 2011, 6(10): 1624-1626.
[77] Takahashi R, Ishimaru Y, Senoura T, Shimo H, Ishikawa S, Arao T, Nakanishi H, Nishizawa N K.The OsNRAMP1 iron transporter is involved in Cd accumulation in rice[J]., 2011, 62(14): 4843-4850.
[78] Ishimaru Y, Takahashi R, Bashir K, Shimo H, Senoura T, Sugimoto K, Ono K, Yano M, Ishikawa S, Arao T, Nakanishi H, Nishizawa N K.Characterizing the role of rice NRAMP5 in manganese, iron and cadmium transport[J]., 2012, 2(1): 286.
[79] Shimo H, Ishimaru Y, An G, Yamakawa T, Nakanishi H, Nishizawa N K.Low cadmium (LCD), a novel gene related to cadmium tolerance and accumulation in rice[J]., 2011, 62(15): 5727-5734.
[80] Lan H X, Wang Z F, Wang Q H, Wang M M, Bao Y M, Huang J, Zhang H S.Characterization of a vacuolar zinc transporter OZT1 in rice (L.)[J]., 2013, 40(2): 1201-1210.
[81] ODA K, OTANI M, Uraguchi S, Akihiro T, Fujiwara T.Rice ABCG43 is Cd inducible and confers Cd tolerance on yeast[J]., 2011, 75(6): 1211-1213.
[82] Song W Y, Lee H S, Jin S R, Ko D, Martinoia E, Lee Y, An G, Ahn S N.Rice PCR1 influences grain weight and Zn accumulation in grains[J]., 2015, 38(11): 2327-2339.
[83] Hao X H, Zeng M, Wang J, Zeng Z W, Dai J L, Xie Z J, Yang Y Z, Tian L F, Chen L B, Li D P.A node-expressed transporter OsCCX2 is involved in grain cadmium accumulation of rice[J]., 2018, 9: 476.
[84] Luo J S, Huang J, Zeng D L, Peng J S, Zhang G B, Ma H L, Guan Y, Yi H Y, Fu Y L, Han B, Lin H X, Qian Q, Gong J M.A defensin-like protein drives cadmium efflux and allocation in rice[J]., 2018, 9(1): 645.
[85] Uraguchi S, Kamiya T, Sakamoto T, Kasai K, Sato Y, Nagamura Y, Yoshida A, Kyozuka J, Ishikawa S, Fujiwara T.Low-affinity cation transporter (OsLCT1) regulates cadmium transport into rice grains[J]., 2011, 108(52): 20959-20964.
[86] Moons A.Ospdr9, which encodes a PDR-type ABC transporter, is induced by heavy metals, hypoxic stress and redox perturbations in rice roots[J]., 2003, 553(3): 370-376.
[87] Chen W R, Feng Y, Chao Y E.Genomic analysis and expression pattern of OsZIP1, OsZIP3, and OsZIP4 in two rice () genotypes with different zinc efficiency[J]., 2008, 55(3): 400-409.
[88] Nakamura T, Yamaguchi Y, Sano H.Four rice genes encoding cysteine synthase: isolation and differential responses to sulfur, nitrogen and light[J]., 1999, 229(1-2): 155.
[89] Yeh C M, Hsiao L J, Huang H J.Cadmium activates a mitogen-activated protein kinase gene and MBP kinases in rice.[J]., 2004, 45(9): 1306-1312.
[90] Cheng L J, Wang F, Shou H X, Huang F L, Zheng L Q, He F, Li J H, Zhao F J, Ueno D, Ma J F, Wu P.Mutation in nicotianamine aminotransferase stimulated the Fe(II) acquisition system and led to iron accumulation in rice[J]., 2007, 145(4): 1647-1657.
[91] Wang C H, Guo W L, Ye S, Wei P C, Ow D W.Reduction of Cd in rice through expression of OXS3-like gene fragments[J]., 2016, 9(2): 301-304.
[92] Miyadate H, Adachi S, Hiraizumi A, Tezuka K, Nakazawa N, Kawamoto T, Katou K, Kodama I, Sakurai K, Takahashi H, Namiko S N, Watanabe A, Fujimura T, Akagi H.OsHMA3, a P1B-type of ATPase affects root-to-shoot cadmium translocation in rice by mediating efflux into vacuoles[J]., 2011, 189(1): 190-199.
[93] Ueno D, Yamaji N, Kono I, Huang C F, Ando T, Yano M, Ma J F.Gene limiting cadmium accumulation in rice[J]., 2010, 107(38): 16500-16505.
[94] Yan J L, Wang P T, Wang P, Yang M, Lian X M, Tang Z, Huang C F, Salt D E, Zhao F J.A loss-of-function allele of OsHMA3 associated with high cadmium accumulation in shoots and grain of Japonica rice cultivars[J]., 2016, 39(9): 1941-1954.
[95] Ye X X, Ma Y B, Sun B.Influence of soil type and genotype on Cd bioavailability and uptake by rice and implications for food safety.[J]., 2012, 24(9): 1647-1654.
[96] 鄂志国, 张玉屏, 王磊.水稻镉胁迫应答分子机制研究进展[J].中国水稻科学,2013, 27(5): 539-544.
E Z G, Zhang Y P, Wang L.Molecular mechanism of rice responses to cadmium stress., 2013, 27(5): 539-544.(in Chinese with English abstract)
[97] 黄新元, 赵方杰.植物防御素调控水稻镉积累的新机制[J].植物学报, 2018, 53(4): 451-455.
Huang X Y, Zhao F J.A defensin-like protein regulates cadmium accumulation in rice [J]., 2018, 53(4): 451-455.(in Chinese with English abstract)
[98] 宗良纲, 徐晓炎.水稻对土壤中镉的吸收及其调控措施[J].生态学杂志, 2004(3): 120-123.
Zong L G, Xu X Y.Cadmium absorption of rice from soils and remediations [J]., 2004(3): 120-123.(in Chinese with English abstract)
[99] Rehman M Z, Rizwan M, Ghafoor A, Naeem A, Ali S, Sabir M, Qayyum M F.Effect of inorganic amendments for in situ stabilization of cadmium in contaminated soils and its phyto-availability to wheat and rice under rotation[J]., 2015, 22(21): 16897-16906.
[100]徐爱春, 陈益泰.镉污染土壤根际环境的调节与植物修复研究进展[J].中国土壤与肥料,2007(2): 1-6.
Xu A C, Chen Y T.Regulation of rhizosphere and phytoremediation of Cd contaminated soil and its research advances[J]., 2007(2): 1-6.(in Chinese with English abstract)
[101]黄凯, 张杏锋, 李丹.改良剂修复重金属污染土壤的研究进展[J].江苏农业科学, 2014, 42(1): 292-296.
Huang K, Zhang X F, Li D.Research progress on remediation of heavy metal contaminated soil by amendments[J]., 2014, 42(1): 292-296.(in Chinese with English abstract)
[102]Lin X Y, Mou R X, Cao Z Y, Xu P, Wu X L, Zhu Z W, Chen M X.Characterization of cadmium-resistant bacteria and their potential for reducing accumulation of cadmium in rice grains[J]., 2016, 569: 97-104.
[103]Lin H M, Fang C X, Li Y Z, Lin W W, He J Y, Lin R Y, Lin W X.Cadmium-stress mitigation through gene expression of rice and silicon addition[J]., 2017, 81(1): 91-101.
[104]Yang Y J, Chen R J, Fu G F, Xiong J, Tao L X.Phosphate deprivation decreases cadmium (Cd) uptake but enhances sensitivity to Cd by increasing iron (Fe) uptake and inhibiting phytochelatins synthesis in rice ()[J]., 2016, 38(1): 28.
[105]Kanu A S, Ashraf U, Mo Z W, Sabir S U R, Baggie I, Charley C S, Tang X R.Calcium amendment improved the performance of fragrant rice and reduced metal uptake under cadmium toxicity[J].,2019, 26(24): 24748-24757.
[106]Cai Y, Lin L, Cheng W, Zhang G, Wu F.Genotypic dependent effect of exogenous glutathione on Cd-induced changes in cadmium and mineral uptake and accumulation in rice seedlings ()[J]., 2010, 56(11): 516-525.
[107]Feng R W, Wei C Y, Tu S X, Ding Y Z, Song Z G.A dual role of Se on Cd toxicity: Evidences from the uptake of Cd and some essential elements and the growth responses in paddy rice[J]., 2013, 151(1): 113-121.
[108]Tang L, Mao B G, Li Y K, Lv Q M, Zhang L P, Chen C Y, He H J, Wang W P, Zeng X F, Shao Y, Pan Y L, Hu Y Y, Peng Y, Fu X Q, Li H Q, Xia S T, Zhao B R.Knockout of OsNramp5 using the CRISPR/Cas9 system produces low Cd-accumulating indica rice without compromising yield[J]., 2017, 7(1): 14438.
[109]马卉, 焦小雨, 许学, 李娟, 倪大虎, 许蓉芳, 王钰, 汪秀峰.水稻重金属镉代谢的生理和分子机制研究进展[J].作物杂志, 2020(1): 1-8.
Ma H, Jiao X Y, Xu X, Li J, Ni D H, Xu R F, Wang Y, Wang X F.Advances in physiological and molecular mechanisms of cadmium metabolism in rice [J]., 2020(1): 1-8.(in Chinese with English abstract)
[110]Kulaeva O, Kliukova M, Afonin A, Sulima A, Zhukov V, Tikhonovich I.The role of plant antimicrobial peptides (AMPs) in response to biotic and abiotic environmental factors[J]., 2020, 65(2): 187-199.
[111]Odintsova T I, Slezina M P, Istomina E A.Defensins of grasses: A systematic review[J]., 2020, 10(7): 1029.
[112]Brunetti P, Zanella L, De P A, Di L D, Cecchetti V, Falasca G, Barbieri M, Altamura M M, Costantino P, Cardarelli M.Cadmium-inducible expression of the ABC-type transporter AtABCC3 increases phytochelatin- mediated cadmium tolerance in Arabidopsis[J]., 2015, 66(13): 3815-3829.
[113]Fu S, Lu Y S, Zhang X, Yang G Z, Chao D, Wang Z G, Shi M M, Chen J G, Chao D Y, Li R B, Ma J F, Xia J X.The ABC transporter ABCG36 is required for cadmium tolerance in rice[J]., 2019, 70(20): 5909-5918.
[114]Suksabye P, Pimthong A, Dhurakit P, Mekvichitsaeng P, Thiravetyan P.Effect of biochars and microorganisms on cadmium accumulation in rice grains grown in Cd-contaminated soil[J]., 2016, 23(2): 962-973.
[115]Sun L J, Wang J, Song K, Sun Y F, Qin Q, Xue Y.Transcriptome analysis of rice (L.) shoots responsive to cadmium stress[J]., 2019, 9(1-2): 10177.
Research Progresses on Response Mechanisms and Control Measures of Cadmium Stress in Rice
XU Xiaobo, AN Penghu, GUO Tianjiao, HAN Dan, JIA Wei, HUANG Wuxing*
(,,,;Corresponding author,:)
Soil cadmium pollution is one of the increasingly serious environmental problems.Rice is the bulk cereal crop with the highest cadmium uptake capacity.The accumulation of cadmium by rice and its transfer to the food chain pose a threat to human health.In recent years, a series of significant advances have been made in research on the uptake and accumulation processes of cadmium in rice and its influencing factors.In this review, three aspects were reviewed.First, the effects of cadmium on rice seed germination, growth and development, and yield.Second, the physiological and molecular mechanisms behind cadmium activation in soil, xylem loading and transport, and transfer of cadmium from phloem to grains.Third, the prevention and control measures such as variety selection, soil remediation, and agronomic measures.It is expected to provide theoretical references for research related to Cd stress in rice and to lay a basis for developing effective strategies to reduce Cd level in rice.
rice; cadmium; physiological mechanism; molecular mechanism; control and prevention measures
10.16819/j.1001-7216.2021.201209
2020-12-11;
2021-03-22。
河南省重点研发与推广专项(202102310026)。
