NaCl胁迫下施加外源钾对多枝柽柳生理的影响
2021-09-12陈亚辉张文韬宋志忠杨庆山张晓勉姜姜
陈亚辉 张文韬 宋志忠 杨庆山 张晓勉 姜姜

摘要:为提高多枝柽柳的耐盐性,探讨K+加入后多枝柽柳在生理方面的变化,以1/2 Hoagland营养液培养为空白组,以含200 mmol/L NaCl的1/2 Hoagland营养液培养和含200 mmol/L NaCl+10 mmol/L KCl的1/2 Hoagland营养液培养为试验组。对处理后7、15、30 d的多枝柽柳新鲜叶片和新生根进行采样,测得其生理指标。试验结果表明:相较于200 mmol/L NaCl处理,200mmol/L NaCl +10 mmol/L KCl处理组多枝柽柳叶绿素含量高,根系活力增强;同时H2O2和MDA含量明显降低,SOD、CAT、POD等抗氧化酶活性明显升高,脯氨酸和可溶性糖等渗透调节物质含量也明显升高。可见,加入外源K+明显提升了多枝柽柳耐盐性,为钾肥在盐碱地使用提升植物耐盐能力提供了理论依据。
关键词:多枝柽柳;盐胁迫;K+;生理特性;耐盐能力
中图分类号: S727.23;S718.43 文献标志码: A
文章编号:1002-1302(2021)15-0142-05
收稿日期:2021-08-13
基金项目:国家自然科学基金面上项目(编号:32071612);山东省农业良种工程项目(编号:2019LZGC009)。
作者简介:陈亚辉(1990—),男,江苏泰州人,博士研究生,助理研究员,主要从事植物生理及生态修复研究。E-mail:chenyahui01@163.com。
通信作者:姜 姜,博士,教授,主要从事植物生理及生态修复研究。E-mail: ecologyjiang@gmail.com。
盐渍土是在全球分布广泛的土壤资源,因其盐分含量高,土壤理化性质差,严重危害植物的生长发育[1-3]。近年来,受人为活动的影响,盐渍土面积不断扩大,如何利用鹽渍土成为亟待解决的重要环境问题。目前,为了尽最大可能利用该类型土壤资源,学者们对如何改良利用盐渍土进行了多方面的探讨。学者们提出的解决方式大致可以分为两大类[4]。一类是通过优化盐碱土的土壤结构,满足植物生长需求。例如,王会等提出通过秸秆还田改善盐化土壤团粒体的稳定性[5];张济世等通过试验表明施用的6种土壤改良剂均能显著提高小麦产量[6]。另一类是从植物生理的角度看待问题,提高植物本身对盐碱土的适应性,增加盐渍土壤的利用率。刘梅等研究发现,施用氮肥时,硝态营养比铵态营养更有利于提高油菜和水稻的耐盐性[7]。也有学者研究表明[8-9],丛枝菌根可以与大部分高等植物共生,提高植物的耐盐能力。
钾离子作为植物生长发育所必需的一价阳离子,在植物细胞生长及代谢中发挥重要作用[10]。近年来,随着高通量测序的发展,不少学者从分子生物学角度提出,钾离子通过影响某些基因的转录、表达和信号转导水平在植物抗逆性中发挥重要作用[11-12]。有学者报道植物的耐盐能力与植物中的钾离子通道和转运蛋白密切相关[13-14]。多枝柽柳(T. ramosissima Lcdcb.)属于广泛使用的盐生植物,具有较为系统的耐盐机制[15-16]。鲁艳等研究发现低浓度(≤100 mmol/L)的NaCl胁迫会促进多枝柽柳生长,高浓度(≥200 mmol/L 的NaCl)下则会抑制其生长[17]。
本试验以多枝柽柳为对象,使用含不同成分的1/2 Hoagland营养液进行处理,测定相关生理指标,探讨盐胁迫下K+处理对多枝柽柳植物生理特性的影响,为钾肥在盐碱地使用提供相关理论依据。
1 材料与方法
1.1 材料
供试多枝柽柳来自山东省林业科学院基地,选取5个月苗龄、长势相似的扦插苗,试验于2019年10月至2021年5月在南京林业大学林学院国家重点实验室展开。将扦插苗用蒸馏水洗净根部,放于24孔水培箱(尺寸为40 cm×30 cm×16 cm),用 1/2 Hoagland营养液,置于温度(26±2) ℃、相对湿度50%~60%的温室大棚中,培养2个月后进行试验。
1.2 方法
采用1/2 Hoagland营养液培养的多枝柽柳为CK组,用含200 mmol/L NaCl的1/2 Hoagland营养液和含200 mmol/L NaCl+10 mmol/L KCl的1/2 Hoagland营养液培养的多枝柽柳为处理组,每3 d更换1次培养液,试验设置3组,每组3次重复。分别在处理后7、15、30 d对多枝柽柳新鲜叶片和新生根进行采样。
1.3 生理指标测定
总叶绿素含量采用95%乙醇与丙酮混合溶液浸泡法[18]测定;根系活力采用TTC比色法[18-19]测定;H2O2含量采用Sergive等的方法[20]测定;MDA含量采用硫代巴比妥酸法[21]测定;SOD活性采用氮蓝四唑还原法[22]测定;POD活性采用愈创木酚法[23]测定;CAT活性采用Aebi的方法[24]测定;脯氨酸含量按照张殿忠等的方法[25]测定;可溶性糖含量采用蒽酮比色法[26]测定。
1.4 数据处理
采用Excel进行数据统计和计算,利用SPSS 26.0进行显著性差异分析,使用Origin 2018软件制图。
2 结果与分析
2.1 NaCl胁迫下施加外源钾对多枝柽柳叶片中总叶绿素含量的影响
由图1可知,在30 d内,CK组的总叶绿素含量没有显著性变化且高于处理组;处理组总叶绿素含量随时间的变化呈减少趋势;在7 d时,200 mmol/L NaCl处理组总叶绿素含量最低,在15 d和30 d时,处理组叶绿素含量显著低于CK组,200 mmol/L NaCl+10 mmol/L KCl处理组的叶绿素含量显著高于200 mmol/L NaCl处理组。
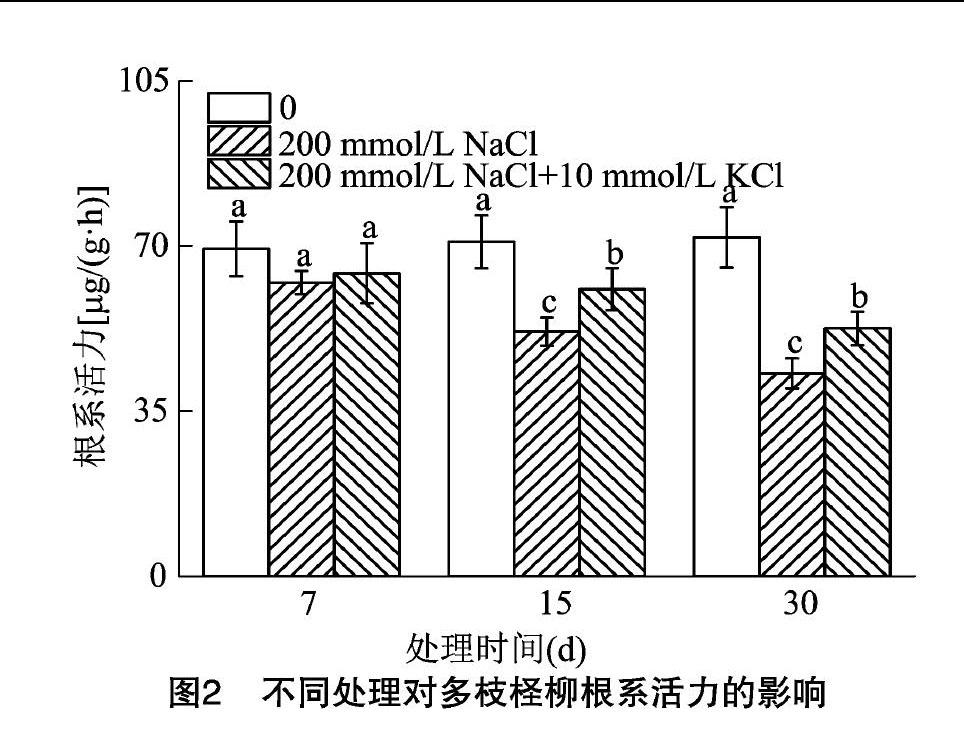
2.2 NaCl胁迫下施加外源钾对多枝柽柳根系活力的影响
由图2可知,在30 d内,CK组的根系活力呈缓慢增加趋势;处理组随时间的变化,根系活力呈减少趋势;其中30 d时,200 mmol/L NaCl处理组的根系活力下降最多。在7 d时,处理组相较于CK组根系活力下降,但无显著性变化。在15 d和30 d时,处理组对比CK组根系活力显著下降,且200 mmol/L NaCl+10 mmol/L KCl处理组的根系活力显著高于200 mmol/L NaCl处理组。
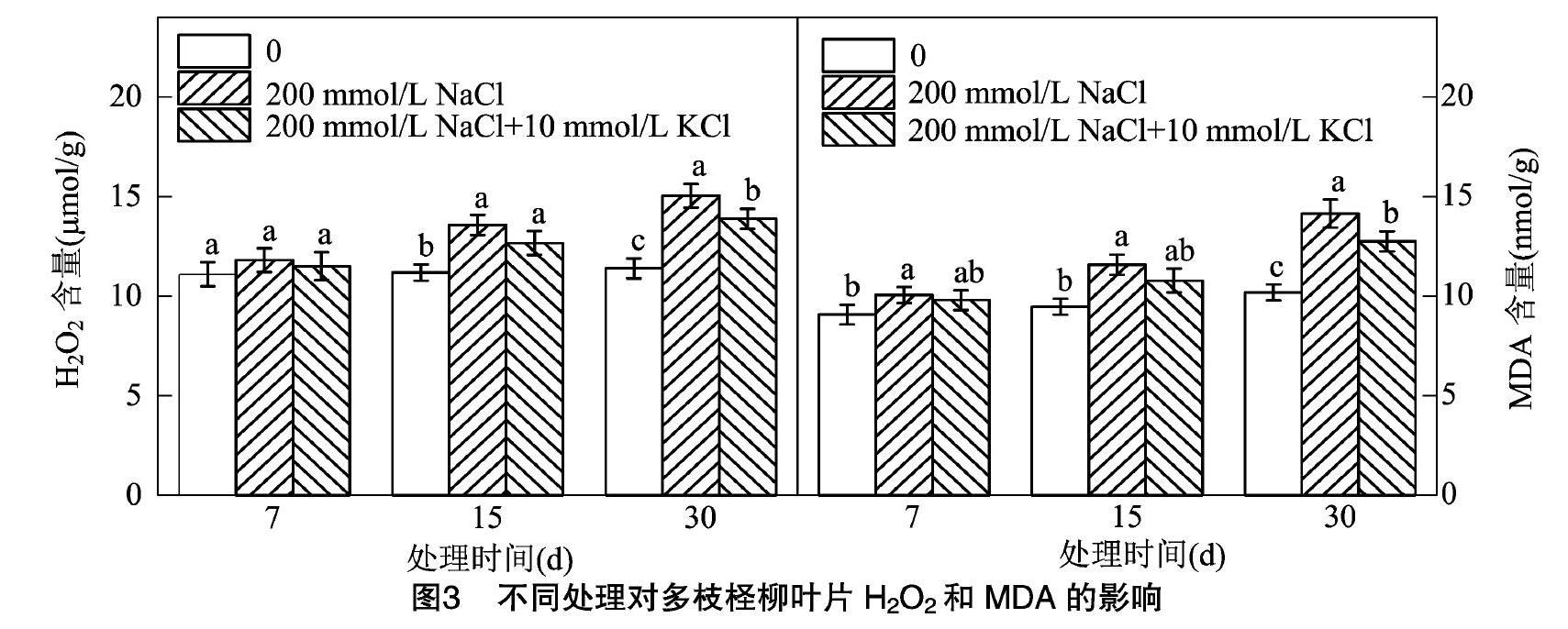
2.3 NaCl胁迫下施加外源钾对多枝柽柳叶片中H2O2与MDA含量的影响
由图3可知,在30 d内, CK组的H2O2含量没有显著性变化;处理组H2O2含量随时间变化呈增加趋势;在7 d时,处理组对比CK组H2O2含量略微增加,无显著性差异。在15 d和30 d时,处理组H2O2含量对比CK组H2O2含量增加明显, 且2种
处理组间差异也显著,200 mmol/L NaCl处理组的H2O2含量增加最多。
在30 d内,CK组的MDA含量呈缓慢增加趋势,但无显著性变化;处理组MDA含量随时间的变化呈增加趋势;在7 d和15 d时,200 mmol/L NaCl处理组比CK组MDA含量高,且有显著性差异;200 mmol/L NaCl+10 mmol/L KCl对比CK组MDA含量略微增加,但无显著性差异。在30 d时,不同处理组对比CK组MDA含量增加明显,且不同处理组间差异显著。200 mmol/L NaCl处理组MDA含量高于200 mmol/L NaCl+10 mmol/L KCl处理组。
2.4 NaCl胁迫下施加外源钾对多枝柽柳叶片抗氧化酶活性的影响
由图4可知,30 d内CK组SOD活性变化不明显;处理组SOD活性随时间变化呈增加趋势;其中,200 mmol/L NaCl+10 mmol/L KCl處理组SOD活性增加最多,且处理组对比CK组SOD活性增加较多,有显著性差异。此外,在30 d内3种处理下POD活性呈缓慢增加趋势;其中,200 mmol/L NaCl处理组POD活性增加最多。在7 d和15 d时,处理组比CK组POD活性较高,且有显著性差异;但处理组间差异变化不明显。在30 d时,处理组显著高于CK组POD活性。同时,30 d内CK组CAT活性变化不明显;处理组随时间的变化,CAT活性呈上升趋势;其中,200 mmol/L NaCl处理组CAT活性增加最多。在7 d时,处理组比CK组CAT活性较高,但无显著性变化。在15 d和30 d时,处理组比CK组CAT活性较高,有显著性差异。值得注意的是,在30 d时200 mmol/L NaCl+10mmol/L KCl处理下3种抗氧化酶活性均显著高于200 mmol/L NaCl。
2.5 NaCl胁迫下施加外源钾对多枝柽柳叶片渗透调节物质含量的影响
通过图5可以看出,30 d内CK组脯氨酸含量变化不明显;处理组随时间变化,脯氨酸含量呈增加趋势;其中,200 mmol/L NaCl+10 mmol/L KCl处理组脯氨酸含量增加最多。在30 d时,200 mmol/L NaCl+10 mmol/L KCl处理组对比CK组脯氨酸含量增加显著,且高于200 mmol/L NaCl处理组。同时,30 d内CK组可溶性糖含量呈缓慢增加趋势;而处理组随时间变化,可溶性糖含量呈增加趋势;其中200 mmol/L NaCl+10 mmol/L KCl处理组可溶性糖含量增加最多。在30 d中,200 mmol/L NaCl+10 mmol/L KCl处理组显著高于其他处理组。
3 讨论与结论
3.1 讨论
植物幼苗在钾素的作用下能够促进光能的转化利用和传递。田晓莉等发现缺钾会导致棉花生物量的降低和早衰[27]。王晓光等研究发现缺钾会导致大豆叶面积减少,叶绿素含量降低[28]。本试验研究表明,200 mmol/L NaCl+10 mmol/L KCl处理组的叶绿素含量显著高于200 mmol/L NaCl处理组。钾作为植物必需的三大营养元素之一,几乎参与整个植物生长发育和生理代谢的过程[12]。钾与氮和磷的不同之处在于,钾不参与植物体内任何有
机物质的构成,却对植物的生长发育起着不可缺少的作用。根系活力体现的是植物根系新陈代谢能力的强弱[29],也是反映植物抗逆能力的重要指标[30]。本试验研究表明,多枝柽柳在200 mmol/L NaCl+10 mmol/L KCl处理组的根系活力显著高于200 mmol/L NaCl处理组,说明K+可明显提升盐环境下植物的新陈代谢能力。
同时,本试验结果也证明了施加外源K+后,植株应对盐胁迫产生活性氧的能力明显提高。正常情况下,植物体内ROS的产生与清除是处于动态平衡状态,不会影响植物的生长发育[31],但在逆境条件下,植物细胞这种平衡会被打破,产生大量的活性氧,这些活性氧以其极强的氧化性造成细胞膜脂过氧化反应,从而导致膜系统损伤和细胞氧化[32-33]。H2O2是ROS的一种,而MDA是脂膜过氧化的最终产物。超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、过氧化物酶(POD)是参与H2O2酶促反应清除系统的重要成员。
研究表明,耐盐植物为了适应盐环境,主要做出2种大的改变:一是通过其膜系统的稳定性来维持对其他离子的选择吸收功能;二是通过代谢变化,产生脯氨酸、可溶性糖等渗透调节物质,这些物质都有很高的水溶性,能够降低细胞内水势[34]。本试验结果表明:相较于200 mmol/L NaCl处理,200 mmol/L NaCl+10 mmol/L KCl处理组H2O2和MDA含量明显降低,SOD、CAT、POD等抗氧化酶活性明显升高。脯氨酸,可溶性糖等渗透调节物质含量明显升高。本试验只是从植物生理的角度解释了这一现象,至于是柽柳植物中的哪些基因起到调控作用还需要通过分子生物学手段进行深入挖掘。
3.2 结论
通过比较盐胁迫条件下,外源施加K+对多枝柽柳根系活力、抗氧化酶活性和渗透调节物质含量的影响,发现K+可明显提升盐环境下多枝柽柳的新陈代谢能力,并缓解盐胁迫对多枝柽柳的伤害。
参考文献:
[1]Ghassemi F,Jakeman A J,Nix H A. Salinisation of land and water resources:human causes,extent,management and case studies[M]. Canberra:UNSW Press,1995:1-3.
[2]李建国,濮励杰,朱 明,等. 土壤盐渍化研究现状及未来研究热点[J]. 地理学报,2012,67(9):1233-1245.
[3]俞仁培,陈德明. 我国盐渍土资源及其开发利用[J]. 土壤通报,1999(4):3-5.
[4]李 星,李凯锋. 土壤盐渍化开发利用研究进展[J]. 农业科技通讯,2018(6):244-246.
[5]王 会,何 伟,段福建,等. 秸秆还田对盐渍土团聚体稳定性及碳氮含量的影响[J]. 农业工程学报,2019,35(4):124-131.
[6]张济世,于波涛,张金凤,等. 不同改良剂对滨海盐渍土土壤理化性质和小麦生长的影响[J]. 植物营养与肥料学报,2017,23(3):704-711.
[7]刘 梅,郑青松,刘兆普,等. 盐胁迫下氮素形态对油菜和水稻幼苗离子运输和分布的影响[J]. 植物营养与肥料学报,2015,21(1):181-189.
[8]Juniper S,Abbott L K. Soil salinity delays germination and limits growth of hyphae from propagules of arbuscular mycorrhizal fungi[J]. Mycorrhiza,2006,16(5):371-379.
[9]徐 瑶,樊 艳,俞云鹤,等. 丛枝菌根真菌对盐胁迫下红花幼苗生长及耐盐生理指标的影响[J]. 生态学杂志,2014,33(12):3395-3402.
[10]周峰,李平华,王宝山. K+稳态与植物耐盐性的关系[J]. 植物生理学通讯,2003(1):67-70.
[11]杨玲琴,刘 敬,李 魏,等. 植物钾离子通道AKT1的研究进展[J]. 生物技术通报,2019,35(4):94-100.
[12]陆 潭,陈华涛,沈振国,等. 植物钾通道与钾转运体研究进展[J]. 华北农学报,2019,34(增刊1):372-379.
[13]Fernandez J M,Schroeder J I,Hedrich R. Potassium-selective single channels in guard cell protoplasts of Vicia faba[J]. Nature,1984,312(5992):361-362.
[14]Dennison K L,Robertson W R,Lewis B D,et al. Functions of AKT1 and AKT2 potassium channels determined by studies of single and double mutants of Arabidopsis[J]. Plant Physiol,2001,127(3):1012-1019.
[15]李 芊. 新疆柽柳属植物抗盐机理研究[D]. 乌鲁木齐:新疆农业大学,2002.
[16]刘铭庭. 中国柽柳属植物综合研究图文集[M]. 乌鲁木齐:新疆科学技术出版社,2014:1-79.
[17]鲁 艳,雷加强,曾凡江,等. NaCl處理对多枝柽柳(Tamarix ramosissima)生长及生理的影响[J]. 中国沙漠,2014,34(6):1509-1515.
[18]王学奎. 植物生理生化实验原理和技术[M]. 北京:高等教育出版社,2006.
[19]朱秀云,梁 梦,马 玉. 根系活力的测定(TTC法)实验综述报告[J]. 广东化工,2020,47(6):211-212.
[20]Sergiev I,Alexieva,Karanov E. Effect of spermine,atrazine and combination between them on some endogenous protective systems and stress markers in plants[J]. Comptes Rendus de IAcadmie Bulgare Sciences,1997,51:121-124.
[21]Kosugi H,Kikugawa K. Thiobarbituric acid reaction of aldehydes and oxidized lipids in glacial acetic acid[J]. Lipids,1985,20(12):915-921.
[22]Beauchamp C,Fridovich I. Superoxide dismutase:improved assays and an assay applicable to acrylamide gels[J]. Anal Biochem,1971,44(1):276-287.
[23]Chance B,Maehly A C. Assay of catalases and peroxidases[M]//Methods in enzymology. New York:Academic Press,1955:2,764-775.
[24]Aebi H. Catalase in vitro[M]//Methods in enzymology. New York:Academic Press,1984:105,121-126.
[25]张殿忠,汪沛洪,赵会贤. 测定小麦叶片游离脯氨酸含量的方法[J]. 植物生理学通讯,1990:62-65.
[26]李合生. 植物生理生化实验原理和技术[M]. 北京:高等教育出版社,2003.
[27]田晓莉,王刚卫,杨富强,等. 棉花不同类型品种耐低钾能力的差异[J]. 作物学报. 2008,34(10):1770-1780.
[28]王晓光,曹敏建,蒋文春,等. 钾肥对不同基因型大豆叶片生理功能的影响[J]. 大豆科学,2006,25(2):133-136.
[29]刘小刚,张富仓,杨启良,等. 玉米叶绿素、脯氨酸、根系活力对调亏灌溉和氮肥处理的响应[J]. 华北农学报,2009,24(4):106-111.
[30]田礼欣,李丽杰,刘 旋,等. 外源海藻糖对盐胁迫下玉米幼苗根系生长及生理特性的影响[J]. 江苏农业学报,2017,33(4):754-759.
[31]薛 鑫,张 芊,吴金霞. 植物体内活性氧的研究及其在植物抗逆方面的应用[J]. 生物技术通报,2013(10):6-11.
[32]赵 嫚,陈仕勇,李亚萍,等. 外源GABA对盐胁迫下金花菜种子萌发及幼苗抗氧化能力的影响[J]. 江苏农业学报,2021,37(2):310-316.
[33]姜卫兵,高光林,俞开锦,等. 水分胁迫对果树光合作用及同化代谢的影响研究进展[J]. 果树学报,2002,19(6):416-420.
[34]马焕成,王沙生. 胡杨膜系统的盐稳定性及盐胁迫下的代谢调节[J]. 西南林学院学报,1998,18(1):15-23.
