闽楠种实性状表型多样性与环境地理因子的关系1)
2021-09-07欧汉彪董利军李娟林建勇梁瑞龙
欧汉彪 董利军 李娟 林建勇 梁瑞龙
(广西壮族自治区林业科学研究院,南宁,530002) (融水县国营贝江河林场) (广西壮族自治区林业科学研究院)
种实表型是植物生命史的一项重要特征,决定了天然建群及群落演替。由于不同种源分布范围广,地理气候差异大,长期自然选择过程中,遗传、环境多样性综合影响植物性状,使其具有丰富的表型多样性[1]。林木表型变异主要来源于3个因素:林木不同的生长环境、林木间的遗传差异、林木基因型与林木生长环境间的相互作用[2]。植物种实的表型性状是植物分类学上重要的辨别依据,具有稳定的遗传特征,不易受区域环境影响,有力的反映群体遗传规律、环境变异特征。目前,国内外研究了文冠果(Xanthocerassorbifolia)[3]、蒜头果(Malaniaoleifera)[4]、蒙古栎(Quercusmongolica)[5]、西伯利亚杏(Armeniacasibirica)[6]、油茶(Camelliaoleifera)[7]、无患子(Sapindusmukorossi)[8]、砂生槐(Sophoramoorcroftiana)[9]、枫香(Liquidambarformosana)[10]等阔叶树种的种实表型性状,为不同树种种质资源收集及利用、遗传选育奠定了基础。
闽楠(Phoebebournei)是樟科(Lauraceae)楠属植物,主要分布长江中下游以南,如福建、浙江、江西、湖北、湖南、四川、贵州、重庆、广东、广西等地区,是我国特有的珍贵用材树种。闽楠花期在4月,果实于10月中旬—11月中旬成熟,果形呈椭圆形或长圆形,长1.1~1.5 cm,直径6~7 mm,成熟时由青变紫黑,表面光滑。国内对闽楠进行了大量的研究,包含造林[11]、栽培[12]、天然林[13-14]、光合特性[15]、木材材性[16]、遗传基因[17]等方面,李娟等[18]对9个闽楠种源的种子形态特征、营养成分进行了研究,但未研究环境地理因子对9个闽楠种源种实的影响。本研究对闽楠天然分布区内14个种源的种实表型性状进行研究,探索种源间及种源内变异来源的规律,研究闽楠种实与环境地理因子的关系,探究区域环境对表型性状的影响机制,以期为闽楠种质资源保存、利用、开发提供参考。
1 材料与方法
1.1 群体选择及试验材料采集
于2016—2019年,采集全国闽楠天然分布范围内9省(区、市)35个种源365个单株的闽楠树果实。母树选择结实良好、生长健壮的优良单株,株距100 m以上。果实成熟时,各单株采果2 kg以上,带回实验室进行果实表型数据测量,之后洗净果皮、果肉,自然阴干,进行种子表型数据及种子质量测量。由于闽楠天然种群数量极少,种群规模存在差异,同时满足至少5株株距100 m及单株采果2 kg的种群有限,所以从35个种源中选取14个具有代表性且满足条件的天然群体,平均每个种源采集8.2株,12月完成全部果种的采集,各种源地理因子见表1。
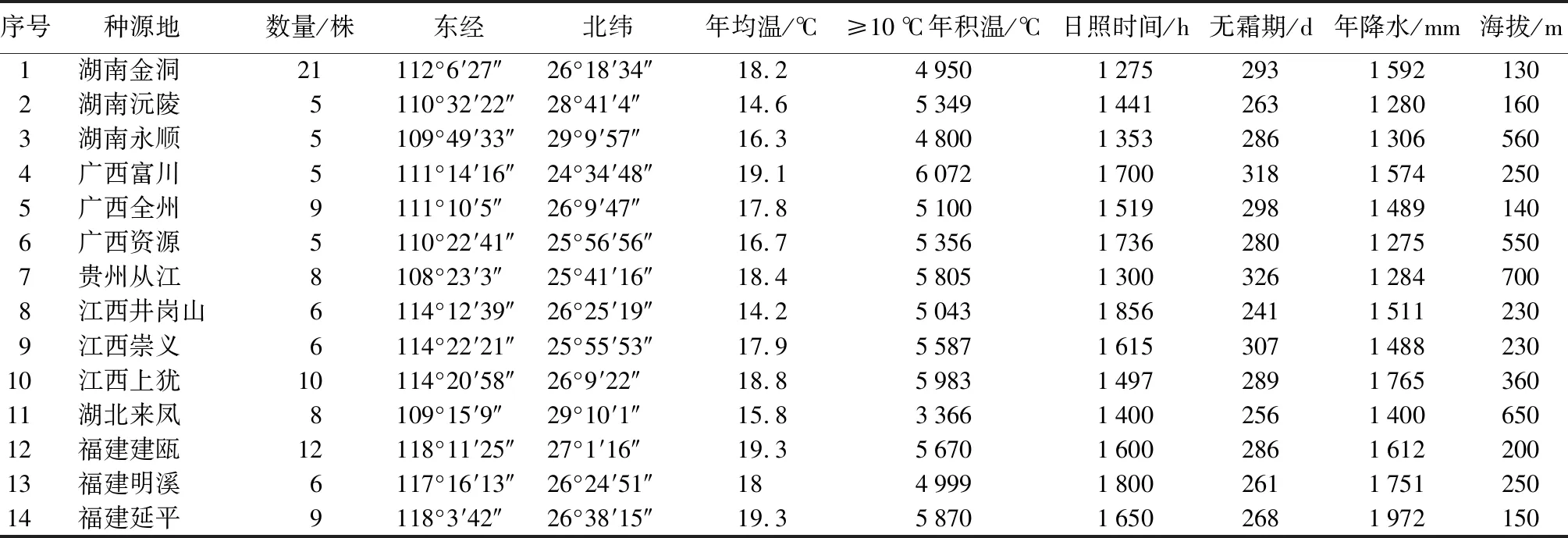
表1 闽楠14个种源地理气候因子
1.2 表型测定
每个种源选取单株数量如表1所示,每个单株随机取60个果实,用游标卡尺对果实、种子的纵径、横径进行测量,随机取1 000粒种子测量种子质量,重复3次。
种形指数=种子纵径/种子横径;
果形指数=果实纵径/果实横径。
1.3 数据分析
采用SAS 9.4软件进行巢式分析、LSD多重分析,采用Canoco5.0软件进行RDA分析,采用R3.6.1软件分析热图。
2 结果与分析
2.1 闽楠种实表型性状
巢式方差分析将闽楠种实性状变异分为种源间、种源内、机误3个部分,以每部分的方差分量比例反映闽楠的变异来源。
由表2可以看出,闽楠种实的表型形状在种源间、种源内均表现出极显著差异。闽楠种实性状的表型分化系数处于52.43%~84.72%,均值为70.83%,果实纵径的表型分化系数最大(84.72%),种子横径的最小(52.43%)。种源间的方差分量占总变异的46.31%,种源内占18.93%,随机误差占34.76%,表明闽楠种实表型变异主要为种源间变异,种源间种实表型多样性大于种源内。
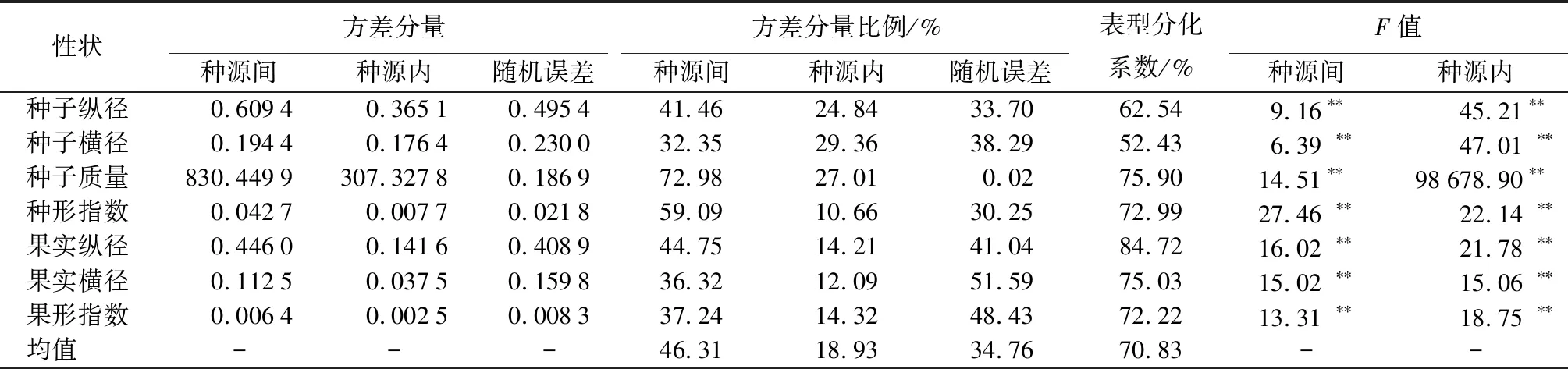
表2 闽楠树种子与果实方差分量及种群表型分化系数
对种源内种实各性状进行多重比较,由表3可知,所有种实表型性状在种源内均存在显著差异,其中福建明溪的种子纵径最长,湖南永顺的最短;广西全州的种子横径最宽,福建建瓯的最窄;湖北来凤的种子质量最重,江西井岗山的最轻;福建建瓯的种形指数最高,贵州从江的最低;江西崇义的果实纵径最长,湖南永顺的最短;福建建瓯、延平的果实横径最宽,江西井岗山的最窄;江西崇义的果形指数最高,湖南永顺的最低。
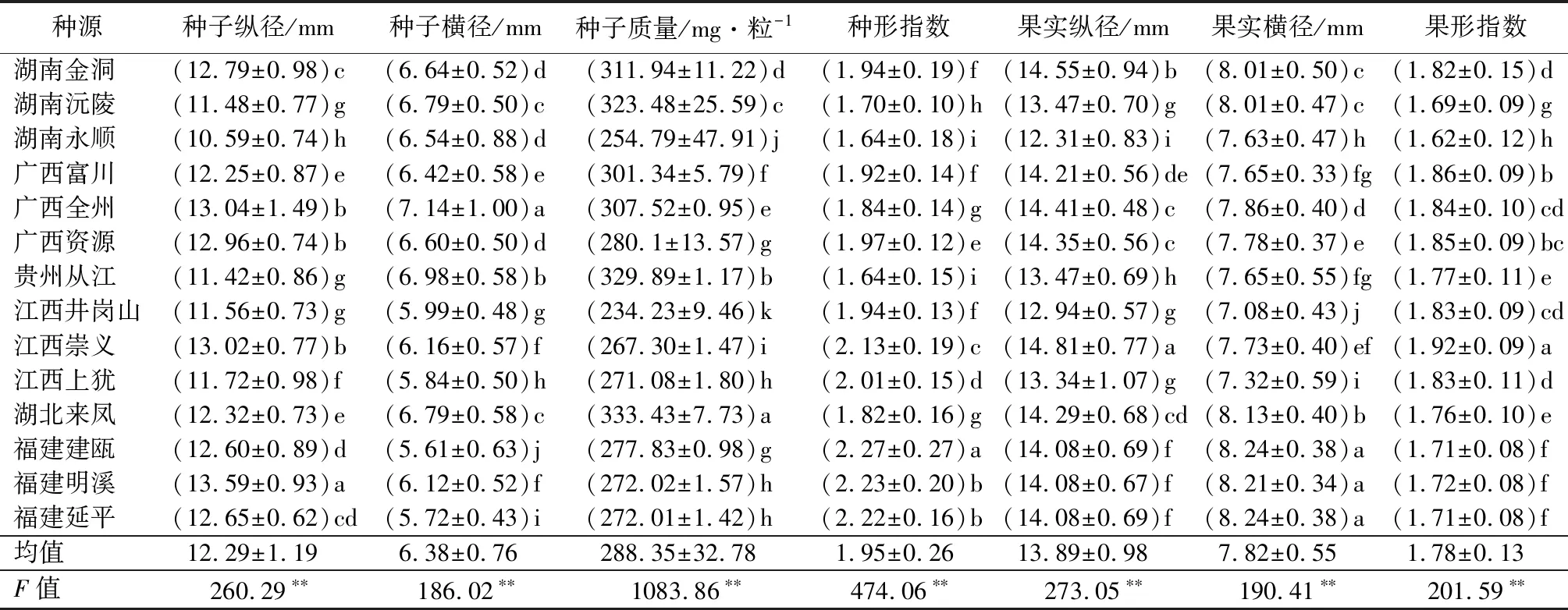
表3 闽楠种实性状种群内的多重比较
对种源间的闽楠种实表型性状变异进行分析(表4),结果表明:所有性状中,种形指数的变异系数最高(13.33%),果实横径的变异系数最低(7.03%)。不同种源间种子纵径、横径、种子质量、种形指数的变异系数均高于果实纵径、横径、果形指数,表明闽楠种源间种子变异程度高于果实。
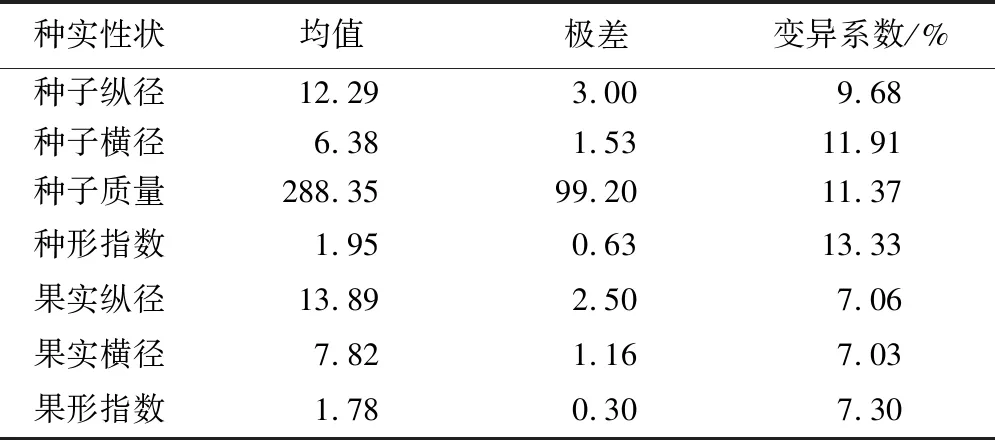
表4 闽楠种实性状的变异系数
对种源内闽楠种实表型性状变异进行分析(表5)。结果表明,不同性状的变异系数均值变化幅度为0.31%~13.96%。种源内各性状的变异系数与种源间的存在差异,其中,种子横径最大(9.23%),种子质量最小(2.13%),表明种子横径变异最丰富,种子质量性状最稳定。从种实性状来看,种子质量的极差最大(7.60%),其中湖南沅陵的种子质量最大,广西全州的最小。果形指数极差最小(3.91%),其中湖南金洞的果形指数最大,江西崇义的最小。不同种源存在不同程度的变异,其中湖南永顺种源的平均变异系数最高,为7.62%;福建延平最低,为4.93%。福建延平每个性状的变异系数均小于湖南永顺,表明湖南永顺种源的变异最为丰富,而福建延平种源的变异最稳定。
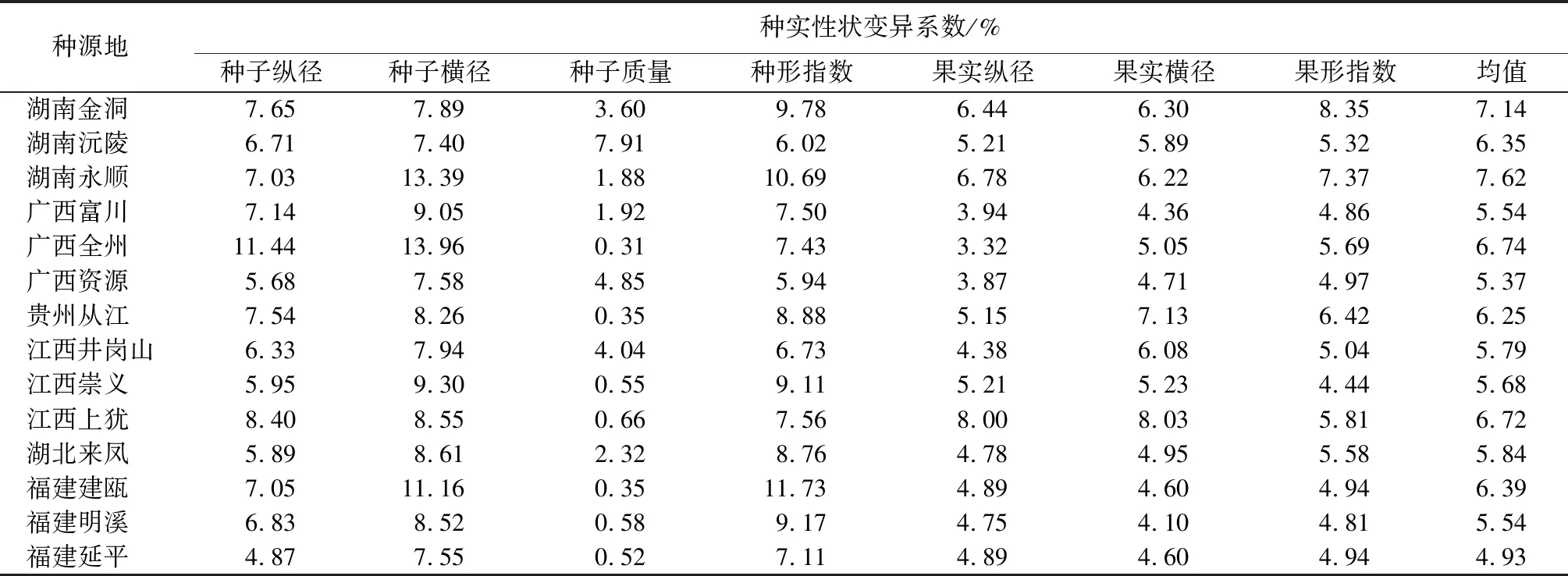
表5 闽楠种实性状种群内的变异系数
2.2 闽楠种实表型性状与环境地理因子相关性
乔木适应生长过程中,不同种源的种实表型性状指标受地理环境因子的综合影响。为了更加直观地表现种实表型性状指标与地理环境因子之间的关系,对其进行冗余分析。根据RDA模型对种实性状特征及地理环境因子进行排序(表6)。Monte Carlo 检验表明,典范轴整体显著性检验表现为极显著(P<0.01),表明RDA排序结果可信,可以较好地解释种实性状与环境地理因子间的关系[18]。年降水相关性极显著,解释率达29.3%,贡献率达55.6%,表明年降水与闽楠种实表型性状相关性最强,为关键影响因素;经度、海拔、日照时间相关性显著,贡献率均处于10%左右,表明经度、海拔、降水与闽楠种实表型性状相关性次之,为主要影响因素;其他地理环境因子相关性不显著,对闽楠种实表型性状无显著影响。
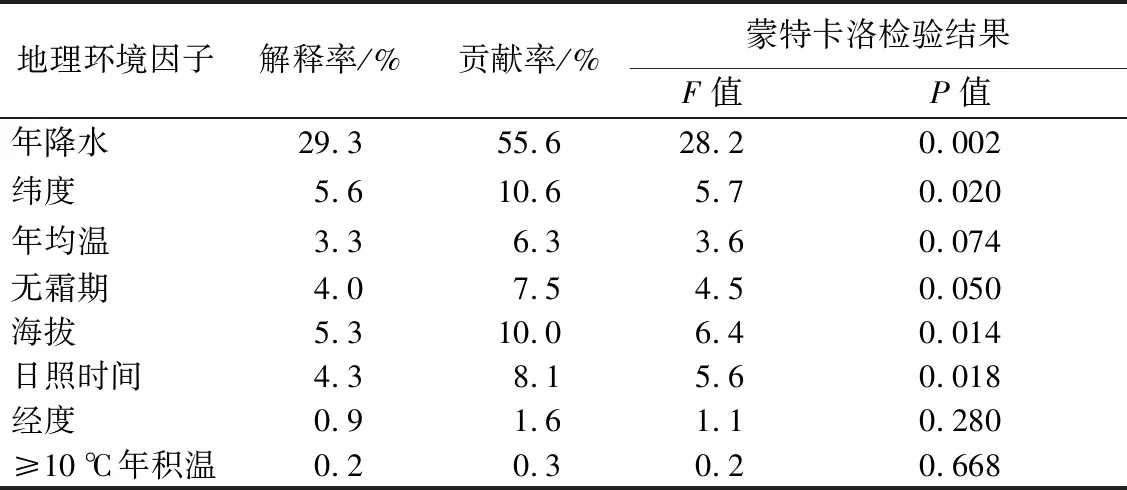
表6 基于RDA模型8种地理气候因子的贡献率和相关性
RDA双序图能较直观的反映闽楠表型性状与地理环境因子的关系(图1),红色空心箭头为环境地理因子,蓝色实心箭头为种实特性,红色箭头连线长度为地理环境因子对种实性状影响的大小,箭头连线间夹角的余弦值表示相关性,样方连线距离为相似程度。所有地理环境因子对闽楠表型性状因子前2轴的总解释率为52.7%(第1轴为52.62%,第2轴为 0.08%)。种子横径、种子质量与年降水、经度、日照时间、≥10 ℃年积温呈负相关,与纬度、海拔、无霜期呈正相关。果实横径、果实纵径、种子纵径、果形指数、种形指数与年均温、年降水、经度、日照时间呈正相关,与纬度、海拔呈负相关。分析种实表型性状间相关性可知,种子横径与种子质量呈正相关,种子越宽质量越重;种子横径、种子质量与种形指数呈负相关,种子越狭长质量越轻。果实横径、果实纵径、种子纵径、果形指数4个性状间呈正相关,但果实纵径和果形指数相关性不明显,既果实变异较小,符合表5分析结果。
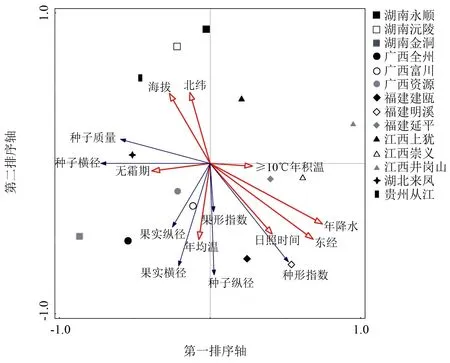
图1 闽楠种实表型性状与地理环境因子RDA双序图
对种源和种实表型性状进行热图分析(图2)。种源共分为3大类,湖南永顺、江西井岗山、江西上犹3个种源为第1类;贵州从江、湖南沅陵、湖北来凤、广西全州、湖南金洞5个种源为第2类;福建明溪、福建建瓯、福建延平、江西崇义、广西富川、广西资源6个种源为第3类。从颜色梯度、相似程度分析种源间、性状间的相似性及差异性。第1类果实横径、种子纵径、果实纵径、种子质量同第2类、第3类相比,均偏向“-2”颜色梯度,表现为种实较瘦小;第2类种形指数、种子纵径、果实纵径同第3类相比,偏向“-2”颜色梯度,种子横径、种子质量同第1类、第3类相比,偏向“2”颜色梯度,表现为种实较宽阔饱满;第3类种子纵径、果实纵径整体颜色更偏向“2”梯度,表现为种实较狭长。
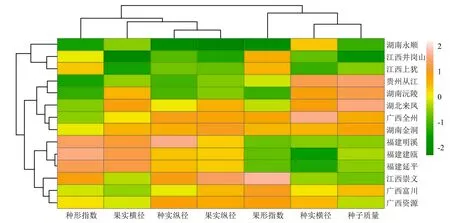
图2 闽楠种实表型性状热图
3 结论与讨论
对闽楠14个天然种源的7个种实表型性状进行研究,发现闽楠在种源间、种源内都存在极显著差异,且种源间变异比种源内变异更丰富,这与李娟等[19]对闽楠不同地理种群遗传多样性ISSR研究结果一致,反映出闽楠长期的地理隔绝阻碍了基因交流,也是植物对环境选择、适应的结果。闽楠的平均表型分化系数为70.83%,种源间的方差分量比例(46.31%)远高于种源内的(18.93%)。与其他物种相比,闽楠的平均表型分化系数低于夏蜡梅(Calycanthuschinensis)的(89.30%)[20]、高于枫香的(57.55%)[10]、五角枫(Acerpictumsubsp.mono)的(48.42%)[21]、云南含笑(Micheliayunnanensis)的(24.38%)[22]、皂荚(Gleditsiasinensis)的(20.42%)[23]、元宝枫(Acertruncatum)的(11.46%)[24],表明闽楠的种实表型性状分化属于偏高水平,种源间变异较丰富。
植物种实受遗传和环境的双重影响,表现出不同程度的变异。本研究中闽楠的平均变异系数为6.14%,与其他物种相比,闽楠的平均变异系数高于红砂(Reaumuriasoongarica)的(1.93%)[25],低于枫香的(15.83%)[10]、元宝枫的(11.46%)[24]、国槐(Amorphafruticosa)的(9.40%)[26]、江南油杉(Keteleeriafortuneivar.cyclolepis)的(7.25%)[27],表明闽楠果实与种子表型性状变异相对稳定。闽楠种子纵径(7.04%)、横径(9.23%)平均变异系数高于果实纵径(5.12%)、横径(5.52%)的,其果实表型性状比种子更稳定,这与关于无患子[8]、江南油杉的研究结果一致,与皂荚[23]、思茅松(Pinuskesiyavar.langbianensis)[28]的研究结果相反。种实是植物建群及天然更新的主要因素,其较大的变异程度体现出闽楠适应生殖的繁育策略[10]。
不同的物种对环境因子存在不同的反映,产生的变异趋势也不同。研究表明,影响枫香种实的主要地理气候因子为1月平均气温、海拔、纬度[10]。川西云杉(Picealikiangensisvar.rubescens)的球果性状随经度增加而变大[29];红砂种子表型性状根据地理位置由西到东,种子质量、种子长、种子长宽比均有所增大,根据地理位置由北到南,种子长、种子宽、种子长宽比匀有所降低[25];元宝枫翅果表型性状根据地理位置由西向东有变长、变宽的趋势[24];青海云杉(PiceacrassifoliaKom)种实性状根据地理位置从东向西,球果逐渐变大变重[30];苦楝(Meliaazedarach)在南部、东部种群的种粒比北部、西部种群的小[31]。江南油杉种实表型随着分布区海拔的升高,形状由椭圆形趋于圆形,形态逐渐饱满[27];岷江柏木(Cupressuschengiana)表现为高海拔地区球果质量较好,但种子在低海拔地区质量更优良[32]。本研究发现,闽楠种子表现为年降水丰富地区种子生长偏向狭长的变异模式。经度、海拔、日照时间是影响种实表型性状的次要因素。结合RDA双序图发现,经度、日照时间与种形指数呈正相关,与种子质量呈负相关。随经度增加,种子呈近椭趋势,高经度种子更饱满厚重。海拔与果形指数、种形指数均呈负相关,随海拔的增加,果实、种子呈近圆趋势。在日照时间较少、海拔较高、降水较少的地区,种子为了适应恶劣的环境,进化成宽阔厚重的种子,既能满足营养物质的积累,也能维持外在条件的抗压。
闽楠分布范围宽泛,不同分布区域具有不同气候条件。植物为了适应环境,其表型性状逐渐具有区域特征。本研究发现,湖南永顺、江西井冈山、江西上犹闽楠种实的表型性状表现为果种较瘦小,为第1类;贵州从江、湖南沅陵、湖北来凤、广西全州、湖南金洞的果种较宽阔饱满,为第2类;福建明溪、福建福州、福建延平、江西崇义、广西富川、广西资源的果种较狭长,为第3类。地理位置来看,江西南部为主的第1类种实主要位于湖南罗霄山脉和福建武夷山山脉之间,湖南永顺受武陵山山脉影响,其种实划分为第1类,第1类种实分布区日照时间较长、无霜期较短;湖南及周边的第2类种实分布区因湖南罗霄山脉与南岭隔绝,日照时间较短、海拔较高、降水较少;福建及东南线的第3类种实分布区因福建武夷山与湖南南岭山脉隔绝,降水较丰富、年均温较高、海拔较低。山系的存在影响闽楠异花授粉及种子传播,种源间无法进行基因交流,增加了种源间分化的可能性,导致种实表型性状的丰富变异。
