SARS-CoV-2 Spike蛋白诱导人肾上皮细胞 周期阻滞及细胞凋亡
2021-09-02梁文翰毛莹莹贾敏胡志明李金龙李红卫
梁文翰, 毛莹莹, 贾敏, 胡志明, 李金龙, 李红卫*
(1.南方医科大学 检验与生物技术学院, 广东 广州 510515;2.南方医科大学 皮肤病医院/广东省皮肤病医院 检验科, 广东 广州 510515)
自2019年12月,由新型冠状病毒SARS-CoV-2引起的严重急性呼吸系统疾病迅速流行,成为影响几乎所有国家的流行病[1].在短时间内,这种感染已导致全球大流行,并已成为对当今人类社会生存的重大威胁[2-4].根据世界卫生组织公布信息,截至2020年11月17日,全球已有超过5 400万确证病例,造成超过132万人死亡.
冠状病毒由一个大而多样的包膜正链RNA病毒家族组成,其基因组长约27~32 kb[5].SARS-CoV-2在结构上主要有四种结构蛋白:膜蛋白(membrane protein,M蛋白),核衣壳蛋白(nucleocapsid protein,N蛋白),包膜蛋白(envelope protein,E蛋白)和刺突蛋白(spike protein,S蛋白)[6].其中SARS-CoV-2 S蛋白由2个亚基组成:S1亚基介导与宿主细胞受体结合,S2亚基介导病毒膜和细胞膜的融合,其中受体结合位区(receptor-binding domain, RBD)位于S1亚基上[7-8].SARS-CoV-2 S蛋白以三聚体的形式组成病毒颗粒外膜表面的刺突,其主要负责识别宿主细胞表面受体血管紧张素转移酶2(angiotensin I-converting enzyme 2, ACE2),从而介导与宿主细胞的融合[9-11].研究表明,SARS-CoV-2的S蛋白与ACE2的亲和力更强,约比SARS的结合能力要高出10~20倍,这提示表达ACE2的细胞更容易受到SARS-CoV-2 的感染[7].
SARS-CoV-2 感染引起的COVID-19引起了全球的关注,随着研究的不断深入,SARS-CoV-2感染与全身多脏器受损间的关系及SARS-CoV-2感染的机制陆续得到揭示[12].文献报道证实COVID-19患者最常见的肾脏功能异常是轻度至中度蛋白尿[13].只有一小部分血浆蛋白被肾小球过滤,并且大多数被近端肾小管有效地吸收,因此在正常尿液中基本上没有蛋白出现.肾小球滤过屏障取决于其三个组成部分上皮细胞、肾小球基底膜和足细胞的功能[14],而最近已经确定了SARS-CoV-2与足细胞的亲和力,其可能影响肾小球滤过功能并引起滤过血浆蛋白增加[15].然而SARS-CoV-2感染引发急性肾衰竭的相关机制仍有待研究.HK-2细胞是人肾小管上皮细胞,高表达ACE2,属于SARS-CoV-2 的易感细胞,而HEK293T(以下简称293T)细胞属于人肾上皮细胞系,来源于人胚胎肾细胞,其几乎不表达胞外配体所需要的受体,是科学研究最常用的肾上皮细胞之一[16].本研究利用SARS-CoV-2 S蛋白重组慢病毒载体,探讨其诱导的SARS-CoV-2 S蛋白过表达对肾上皮细胞的生长的影响及相关机制,为研究SARS-CoV-2 感染后期发生肾功能损伤的原因提供实验依据.
1 材料与方法
1.1 试剂与仪器
HK-2细胞株、293T细胞株购自ATCC,慢病毒载体质粒由南方医科大学生物治疗研究所保存.SARS-CoV-2 S蛋白质粒(VG40589-UT)购自北京义翘神州有限公司.限制性内切酶Nhe I和Mlu I、T4 DNA连接酶均购自美国New England Biolabs公司; CCK-8细胞活力检测试剂盒购自东京同仁公司;细胞基础培养基、体积分数为0.25%的Trypsin(EDTA)、PBS粉末和胎牛血清均购自Gibco;细菌培养用抗生素、琼脂粉、100×双抗购自美国海克隆公司;蛋白Marker、DNA Marker购自北京康润科技有限公司; Trizol裂解液、Primer STAR、逆转录试剂和SYBR Premix Ex TaqTM均购自宝生物工程(大连)有限公司;细胞周期流式检测试剂盒和细胞凋亡流式检测试剂盒购自杭州联科生物技术股份有限公司;Caspase-3抑制剂购自美国Santa公司,Caspase-8,Caspase-9抑制剂购自美国Abcam公司;使用的一抗均购自CST公司;HRP标记抗羊鼠二抗购自杭州弗德生物科技有限公司.
超净台(双人单面)购自力康生物医疗科技控股有限公司;CO2细胞培养箱购自美国赛默飞世尔公司;凝胶成像仪购自美国GE ImageQuant公司;倒置显微镜购自日本奥林巴斯公司;小型高速冷冻离心机(5910型)购自德国艾本德(Eppendorf)公司;超低温冰箱购自美国赛默飞世尔公司;FACSCelestaTM流式细胞仪购自美国BD公司.
1.2 研究方法
1.2.1 细胞培养
HK-2细胞使用DMEM/F12培养基(含体积分数为10%的胎牛血清、体积分数为1%的青霉素和体积分数为1%的链霉素),置于37 ℃,CO2体积分数为5%的培养箱中培养;293T细胞用 DMEM培养基(含体积分数为10%的胎牛血清、体积分数为1%的青霉素和体积分数为1%的链霉素),置于37 ℃,CO2体积分数为5%的培养箱中培养.
1.2.2 目的基因及引物的设计与合成
根据北京义翘神州有限公司提供的SARS-CoV-2 S蛋白质粒(VG40589-UT)的基因序列,在5’及3’端分别加入Nhe Ⅰ和Mlu Ⅰ酶切位点,同时设计相关的引物序列(表1),交由擎科生物科技有限公司合成.

表1 引物及序列Table 1 Primers and sequences
1.2.3 载体质粒的构建
SARS-CoV-2 S蛋白质粒(VG40589-UT)作为模板用PCR扩增出S基因全长,用Nhe Ⅰ和Mlu Ⅰ对PCR产物和慢病毒载体质粒pLV-CMV-IRES-eGFP进行双酶切,将酶切所得产物凝胶电泳后按片段大小进行切胶回收.将回收到的S片段和载体片段用T4 DNA连接酶16℃过夜连接,将连接产物转化至DH5α 感受态细胞,第二天从细菌平板挑取单克隆菌落进行扩增并提取质粒,将重组质粒命名为pLV-CMV-S-IRES-eGFP,重组质粒经酶切及测序鉴定.
1.2.4 慢病毒载体的包装、浓缩及滴度测定
病毒包装使用三质粒系统,将pLV-CMV-S-IRES-eGFP与辅助质粒VSVG和辅助质粒psPAX2按体积比4∶1∶3的比例转染至293T细胞,在37 ℃,CO2体积分数为5%的细胞培养箱中培养48 h后,收集细胞培养上清并用100KD超滤管进行超滤浓缩,得到慢病毒LV-CMV-IRES-eGFP (以下简称为LV-GFP)以及慢病毒LV-CMV-S-IRES-eGFP (以下简称为LV-S).用RT-PCR检测两种病毒的感染复数(multiplicity of infection,MOI),保存于-80 ℃冰箱.
1.2.5 建立慢病毒感染细胞模型
选择状态良好的HK-2及293T细胞以1×106/孔均匀铺于6孔板中,在37 ℃,CO2体积分数为5%的环境中培养24 h.在第2天,弃去培养基,以MOI为100加入稀释好的LV-S病毒液-聚凝胺(polybrene终质量浓度为8 mg/L)培养基混合液,将细胞培养板放入培养箱中,12 h后更换培养基继续培养.72 h后使用荧光显微镜观察细胞感染荧光蛋白表达情况.
1.2.6 细胞活力检测
(1)EdU法检测细胞增殖 在细胞感染慢病毒后,将培养基换成含10 μmol/L EdU培养液孵育2 h,PBS缓冲液清洗,多聚甲醛固定细胞30 min,经甘氨酸脱色和体积分数为0.5%的TritonX-100通透,后续步骤按照说明书进行.每个孔随机取3个200倍视野,荧光显微镜下观察计数并拍照.
(2)CCK-8法检测细胞增殖 以每孔1×104个细胞接种到96孔板中,细胞贴壁后按MOI为100加入慢病毒,72 h后加入100 μL含体积分数为10%的CCK-8细胞增殖检测试剂的新鲜培养基,孵育1 h,用酶标仪读取450 nm各孔的吸光度.
1.2.7 流式细胞术检测细胞周期和凋亡
(1)检测细胞周期 经离心收集1×106细胞,用PBS缓冲液洗2次,加入-20 ℃预冷的体积分数为70%的乙醇,低温固定过夜.离心后收集细胞,PBS水化,清洗1次,加入1 mL DNA staining solution和10 μL PI 混匀后 4 ℃避光孵育30 min,应用标准程序进行流式细胞仪检测.
(2)检测细胞凋亡 经离心收集1×106细胞,用PBS缓冲液洗2次,按说明书加入Annexin V-FITC,室温孵育15 min,用Binding Buffer洗1次,重悬细胞,加入PI染色液,避光孵育10 min,应用标准程序进行流式细胞仪检测.
1.2.8 RT-PCR检测细胞周期及凋亡的基因表达
用RNAiso Plus提取HK-2及293T细胞总RNA,酶标仪测得RNA质量浓度.以每反应体系1 μg加入RNA,以37 ℃持续15 min, 85 ℃持续5 min进行反转录反应;然后按照试剂盒说明书设置标准PCR程序.每个样本设置3个复孔,采用2-ΔΔCt法计算目的基因的相对表达量,该实验重复3次并取其均值.
1.2.9 Western Blot检测细胞相关蛋白表达
在细胞感染慢病毒72 h后,提取HK-2及293T细胞总蛋白,在质量分数为10%的凝胶中以200 V,40 min的条件电泳,并以200 mA,1.5 h的条件电转移到PVDF膜上.质量分数5%的脱脂奶粉封闭后,以V抗体∶V抗体稀释液=1∶1 000稀释一抗,并置于4 ℃冰箱孵育过夜,洗涤后,以V抗体∶V抗体稀释液=1∶10 000稀释二抗并室温孵育1 h. 使用ECL发光液检测各组细胞中周期、凋亡相关蛋白的表达量,利用Image quant软件测量发光强度.使用Image J软件对蛋白质条带灰度进行分析.
1.3 统计学分析
使用SPSS 20.0统计软件分析,采用t检验或单因素方差分析进行比较,P<0.05为差异有统计学意义.
2 结果
2.1 SARS-CoV-2 S慢病毒载体构建及鉴定结果
重组质粒pLV-CMV-S-IRES-eGFP质粒酶切鉴定的凝胶电泳结果(图1 A)表明,挑取的单克隆质粒DNA均被切割成两段条带,其中小片段大小约为3 900 bp,与原始目的基因质粒序列长度相同,且经测序后结果显示序列无突变.说明重组质粒pLV-CMV-S-IRES-eGFP构建成功.重组慢病毒包装结果显示,细胞在24 h、48 h、72 h均能表达绿色荧光(图1 B).Western blot结果显示S蛋白成功过表达(图1 C),用RT-PCR法测定其病毒滴度,结果显示为3.85×108IU/mL.这表明慢病毒载体包装成功,并能介导S蛋白在细胞内的表达.
2.2 细胞活力分析
LV-S感染HK-2及293T细胞72 h后,EdU检测细胞增殖,结果显示,与对照组LV-GFP对比,HK-2及293T细胞增殖显著减少(P<0.05)(图2 A, B).CCK-8结果显示,LV-S组与对照组LV-GFP对比,细胞活力明显减少(图2 C).这提示SARS-CoV-2 S蛋白可以显著抑制细胞增殖.
2.3 重组慢病毒载体介导的SARS-CoV-2 S蛋白表达
流式细胞术的结果表明,与对照组细胞相比,过表达SARS-CoV-2 S蛋白的293T细胞的G2/M期占比从6.87%上升到12.48%,HK-2细胞的G2/M期占比从7.54%增加到14.75%(图3 A, B),差异具有统计学意义(P<0.01).过表达SARS-CoV-2 S后,293T和HK-2细胞的Cyclin B1的基因和蛋白水平均明显下降,表明SARS-CoV-2 S蛋白可以诱导细胞发生G2/M期阻滞(图3C, D).
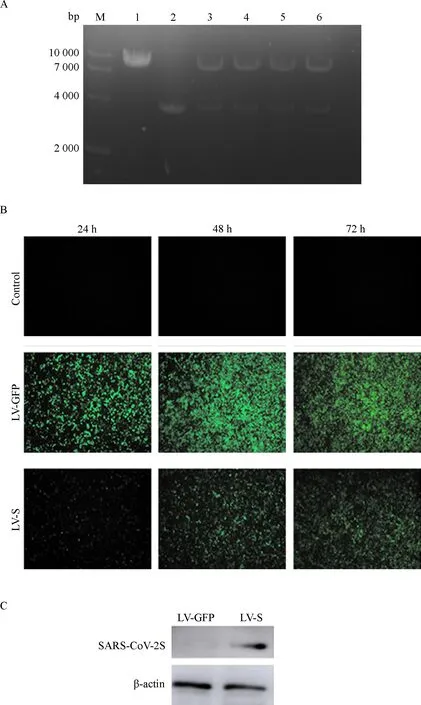
A:pLV-CMV-S-IRES-eGFP酶切鉴定结果, M:DL 10000 DNA 标记物,1:pLV-CMV-S-IRES-eGFP (Nhe Ⅰ & Mlu Ⅰ), 2:SARS-CoV-2 S (Nhe Ⅰ & Mlu Ⅰ),3~6: pLV-CMV-S-IRES-eGFP (Nhe Ⅰ & Mlu Ⅰ); B:质粒转染293T细胞,包装慢病毒,检测24、48、72 h的荧光表达(×40); C:Western blot检测SARS-CoV-2 S蛋白表达.
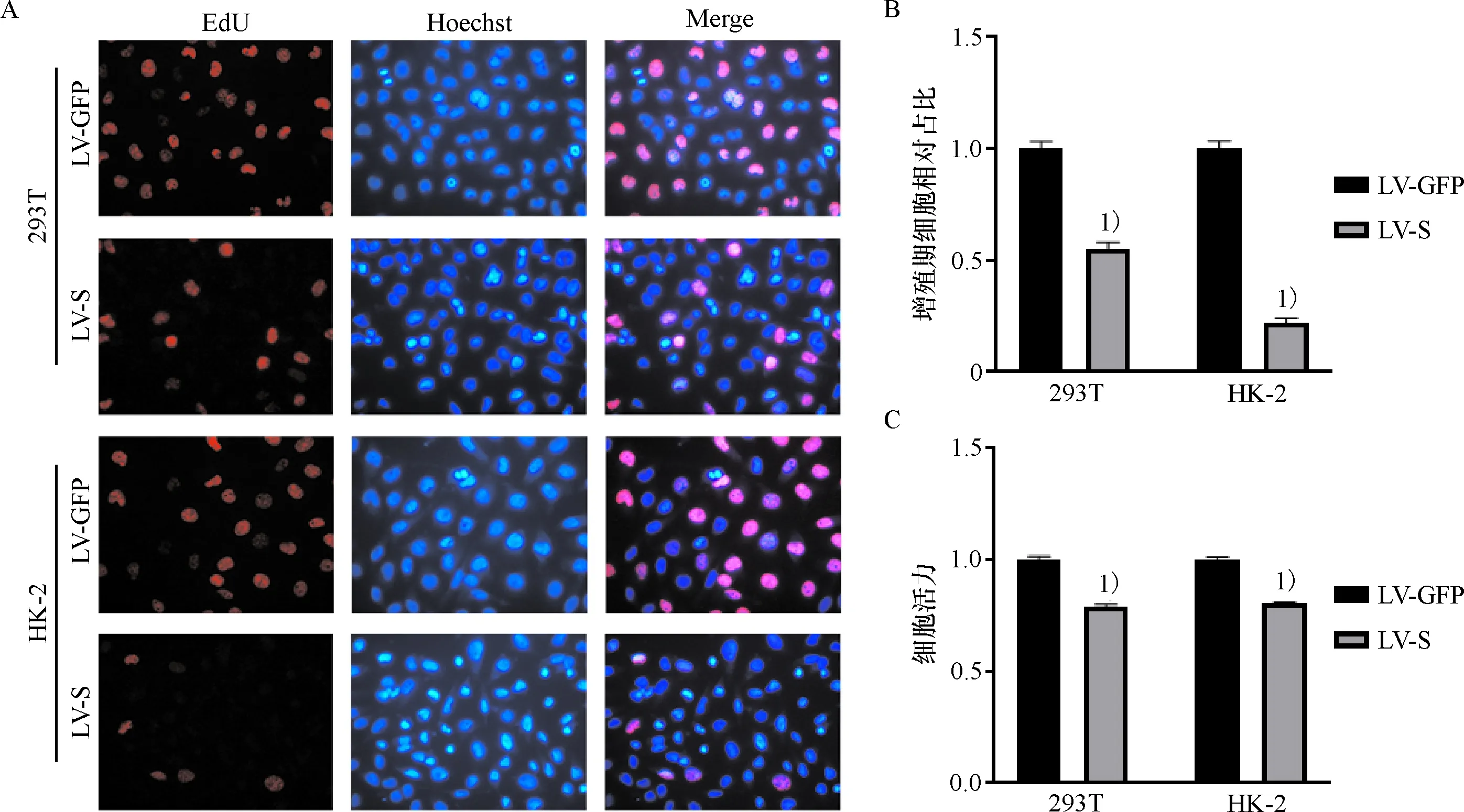
A:EdU检测HK-2细胞及293T细胞增殖(×200);B:对EdU实验进行统计分析;C:CCK-8检测细胞活力.1)与LV-GFP组相比,P<0.05.
2.4 SARS-CoV-2 S蛋白对HK-2细胞及293T细胞凋亡、自噬水平的影响
流式细胞术结果显示,与对照组相比,HK-2细胞过表达S蛋白后,细胞凋亡比例显著增多(从8.61%到27.6%), 293T细胞过表达S蛋白后,细胞凋亡显著增多(从2.04%上升到4.52%)(图4 A,P<0.01).Western blot结果表明,与LV-GFP感染的细胞相比,LV-S感染组细胞p53、Bax、Caspase-3、Caspase-8和Caspase-9蛋白表达均显著升高(图4 B),与之相反,Bcl-2的表达则显著下降.RT-PCR结果显示S蛋白过表达后细胞的Bax基因表达上升,Bcl-2 表达下降(图4 C).提示SARS-CoV-2 S蛋白可以诱导HK-2及293T细胞发生凋亡.
进一步探讨SARS-CoV-2 S诱导细胞发生凋亡的相关机制,采用Caspase-3、Caspase-8和Caspase-9抑制剂处理细胞并检测其凋亡的变化情况.结果表明,应用Caspase-3抑制剂和Caspase-9抑制剂之后, Caspase-3和Caspase-9蛋白水平显著下降.而应用Caspase-8抑制剂之后,Caspase-8表达未发生明显变化.上述结果提示SARS-CoV-2 S蛋白主要可能通过调节Caspase-9的内源性凋亡途径,进而激活下游的Caspase-3凋亡执行蛋白,诱导HK-2及HEK-293T细胞发生凋亡(图4 D).
自噬在细胞本身的代谢需要和某些细胞器的更新过程中发挥重要作用,本研究检测自噬相关蛋白的表达,结果显示自噬的相关蛋白Beclin 1与LC3 B表达上升,p62表达下降,提示SARS-CoV-2 S蛋白导致了HK-2及HEK-293T细胞自噬增强(图4 E).
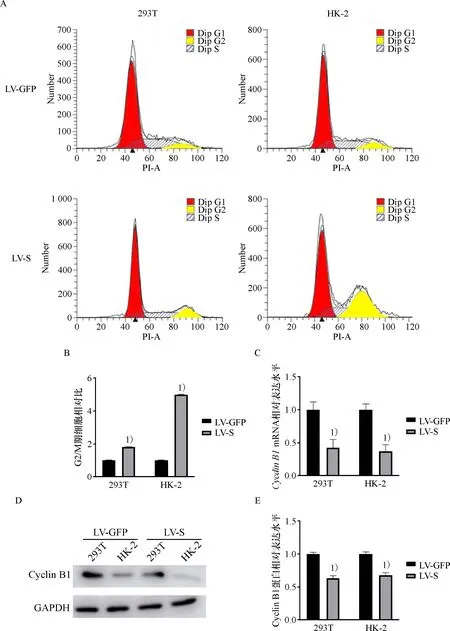
A:流式细胞术检测HK-2及293T细胞的细胞周期;B:G2/M期HK-2及293T细胞的百分比;C:感染的HK-2及293T细胞中Cyclin B1基因的相对水平; D、E:感染的HK-2及293T细胞中Cyclin B1的表达水平; 1)相较于LV-eGFP,P<0.05.

A: 流式细胞术检测SARS-CoV-2 S蛋白导致HK-2及293T细胞凋亡; B: SARS-CoV-2 S蛋白诱导HK-2及293T细胞凋亡相关蛋白表达; C: SARS-CoV-2 S蛋白诱导HK-2及293T细胞 Bax,Bcl-2 mRNA表达, 1)与LV-eGFP比P<0.05; D: Caspase抑制剂对SARS-CoV-2 S诱导的HK-2及293T细胞凋亡的抑制作用;E: SARS-CoV-2导致HK-2及293T细胞自噬相关蛋白表达水平.
3 讨论
数据显示,大多数COVID-19肺炎患者发生肾脏功能异常,虽然这些患者的肾脏功能异常往往在3周内消失[17],但住院的COVID-19患者中有很大一部分患有急性肾损伤,且该类人群的预后较差.在患有肾脏疾病和肥胖的人群中,SARS-CoV-2感染更容易引起多脏器损伤,导致COVID-19死亡率上升[18].本研究结果证实过表达SARS-CoV-2 S蛋白能引起HK-2细胞活力抑制且发生G2/M期阻滞,且过表达SARS-CoV-2 S蛋白能激活Caspase-8、Caspase-9和Caspase-3诱导HK-2细胞发生凋亡.本研究结果提示,SARS-CoV-2 S蛋白在SARS-CoV-2 感染诱导的肾脏损伤中可能发挥重要作用.
细胞周期蛋白B1(Cyclin B1)主要参与细胞从G2期进入M期的调节,在细胞周期循环过程中如果出现Cyclin B1表达降低则可能引起细胞周期阻滞[19].本研究结果显示重组慢病毒载体介导的SARS-CoV-2 S蛋白表达导致HK-2及293T 细胞Cyclin B1 mRNA含量显著下降,Western blot结果也正是其蛋白表达量也显著下调.
已有文献报道多种病毒蛋白能诱导细胞发生凋亡,例如流感病毒的NS1蛋白[20],丙型肝炎病毒的E1/E2蛋白[21]和EB病毒的EBNA3C的N端[22],但是上述蛋白在诱导细胞凋亡的机制往往各不相同[23].
大量研究表明p53是调节细胞凋亡的一个重要蛋白,p53主要通过抑制细胞的有丝分裂信号和生存信号,促进凋亡信号的发生从而诱导细胞凋亡[24].据以往文献报道p53能上调Bcl-2凋亡家族蛋白Bax表达,同时抑制抗凋亡蛋白Bcl-2的转录.因此,p53调节的Bax表达上调同时伴随着Bcl-2表达下调,引发细胞内BAX/Bcl-2比例失衡,最终导致细胞凋亡[25].本研究结果表明,SARS-CoV-2 S蛋白过表达不仅上调了p53蛋白的表达,同时增加了胞内Bax的含量,下调了Bcl-2的表达,上述结果显示p53蛋白参与了SARS-CoV-2 S 介导的HK-2细胞凋亡.研究还发现SARS-CoV-2 S过表达的HK-2及293T细胞中cleaved caspase-3、8、9的含量显著升高,然而仅caspase-3和caspase-9抑制剂能抑制细胞凋亡,结果提示SARS-CoV-2 S通过内源性凋亡通路诱导细胞发生凋亡.
自噬是一个吞噬自身细胞质蛋白或细胞器,并使其包被进入囊泡,然后与溶酶体融合从而形成自噬溶酶体,进而降解囊泡内容物的过程,借此实现细胞代谢需要和细胞器更新[26].在生理或者病理的过程中都可见到自噬的发生,因此自噬所起的作用尚未完全阐明.Beclin1 和LC3 B作为自噬的调控因子和效应因子,在自噬过程中被激活从而引起细胞自噬,p62作为自噬的负调节因子,负向调节自噬流生成[27].本研究发现SARS-CoV-2 S蛋白过表达不仅上调了Beclin 1和LC3 B的表达,说明细胞内自噬流被激活,同时胞内p62减少,说明胞内自噬流通畅,并未受到抑制,提示自噬参与了SARS-CoV-2 S 蛋白介导的HK-2细胞凋亡.
综上,在体内环境下SARS-CoV-2 S蛋白对肾脏功能的影响机制尚不清楚,本研究发现SARS-CoV-2 S蛋白过表达能引起肾小管上皮细胞增殖减弱和凋亡,为进一步研究SARS-CoV-2 的肾脏损伤机制,靶点以及如何影响肾脏功能提供了方向.
作者贡献声明
梁文翰:实验设计者和实验研究执行,数据分析,论文初稿的写作;毛莹莹:实验设计和试验结果分析;贾敏:实验设计和试验结果分析;李金龙:参与论文审阅,指导论文写作;胡志明:参与论文审阅,指导论文写作;李红卫:项目的构思指导论文写作.
利益冲突声明
本研究未受到企业、公司等第三方资助,无利益冲突.
