病毒异戊二烯化修饰及生物学功能研究进展
2021-08-31吴海燕宋影影刘英楠陈鸿军
王 俊,吴海燕,宋影影,刘英楠,陈鸿军
(中国农业科学院上海兽医研究所,上海 200241)
1 概述
蛋白质异戊二烯化修饰是将15个碳原子的法尼基(C15H25)或20个碳原子的香叶基(C15H23)不可逆地共价连接到蛋白质的C基末端半胱氨酸上的一种翻译后修饰[1]。被修饰的蛋白质其C基末端都含有一个保守的CAAX模体,其中C为半胱氨酸,A为疏水性氨基酸,X为任意氨基酸。最近研究证明:C基末端含有延伸的CXXXX和缩短的CXX模体的蛋白也可能是非经典的异戊二烯化修饰酶的底物[2-3]。一般而言,连接异戊烯基的蛋白,其羧基端的CAAX结构中的AAX可以被CAAX蛋白内切酶切割,随后羧基被甲基化修饰(GGTase-2修饰的蛋白不被切割和甲基化)。甲羟戊酸途径和蛋白质异戊二烯化修饰通路如图1所示。

图1 甲羟戊酸途径和蛋白质异戊二烯化修饰通路Fig.1 Mevalonate pathway and protein prenylation
蛋白质的异戊二烯化修饰分别由相应的转移酶FTase和GGTase(GGTase-1、GGTase-2和GGTase3)催化完成[4]。FTase和GGTase-1均为异二聚体,由α亚基和β亚基构成,α亚基由同一个基因FNTA编码,β亚基分别由FNTB和PGGT1B编码。这两种酶识别C末端含有CAAX基序的蛋白质,其中C为半胱氨酸,A为脂肪族氨基酸,X为任意氨基酸。X残基决定了底物蛋白质被法尼基化还是香叶基化。当X为丝氨酸、甲硫氨酸、丙氨酸或谷氨酰胺时,蛋白质发生法尼基化。当X为亮氨酸或异亮氨酸时,蛋白质发生香叶基化。究竟发生哪种修饰,并不是绝对的。例如:当X为苯丙氨酸时,蛋白质既能发生法尼基化,也能发生香叶基化。当某种异戊二烯含量有限时,两种修饰也可能发生互换。例如:当用FTI抑制FTase时,K-Ras蛋白可以发生香叶基化修饰。
发生典型法尼基化修饰后的蛋白质包括许多Ras小G蛋白家族成员(H-Ras、K-Ras、N-Ras、Ras2、Rap2、RhoB、RhoE、Rheb、TC10、TC21)、核纤层蛋白LaminA和LaminB。动粒蛋白CENP-E和CENP-F、cGMP磷酸二酯酶α、G蛋白的γ亚基变体、DnaJ热休克蛋白同源物、视紫红质激酶和过氧化物酶体膜蛋白Pex19和PxF等,GGTase-1修饰的蛋白质底物主要是一些小G蛋白(Rac1、Rac2、RalA、Rap1A、Rap1B、RhoA、RhoB、RhoC、Cdc42、Rab8、Rab11、Rab13等)、G蛋白γ亚基变体、cGMP磷酸二酯酶β和植物的钙调蛋白CaM53等[5]。目前已发现哺乳动物、植物、昆虫、真菌、病毒、原生动物等编码的蛋白质都可以发生异戊二烯化修饰[6],但蛋白质异戊烯基修饰在病毒中鲜有报道。
2 异戊二烯化修饰与病毒
本文从Uniprot-KB数据库、病毒基因组数据库、流感病毒数据库、非洲猪瘟病毒数据库中下载了人类和动物相关病毒蛋白序列,将这些蛋白按不同的病毒科分类分别输入异戊二烯化算法预测服务器网站PrePS(https://mendel.imp.ac.at/PrePS/)进行预测[7],结果如下表1。

表1 人类和动物相关病毒蛋白异戊二烯化修饰预测Table 1 Prenylation prediction of human and animal viral proteins
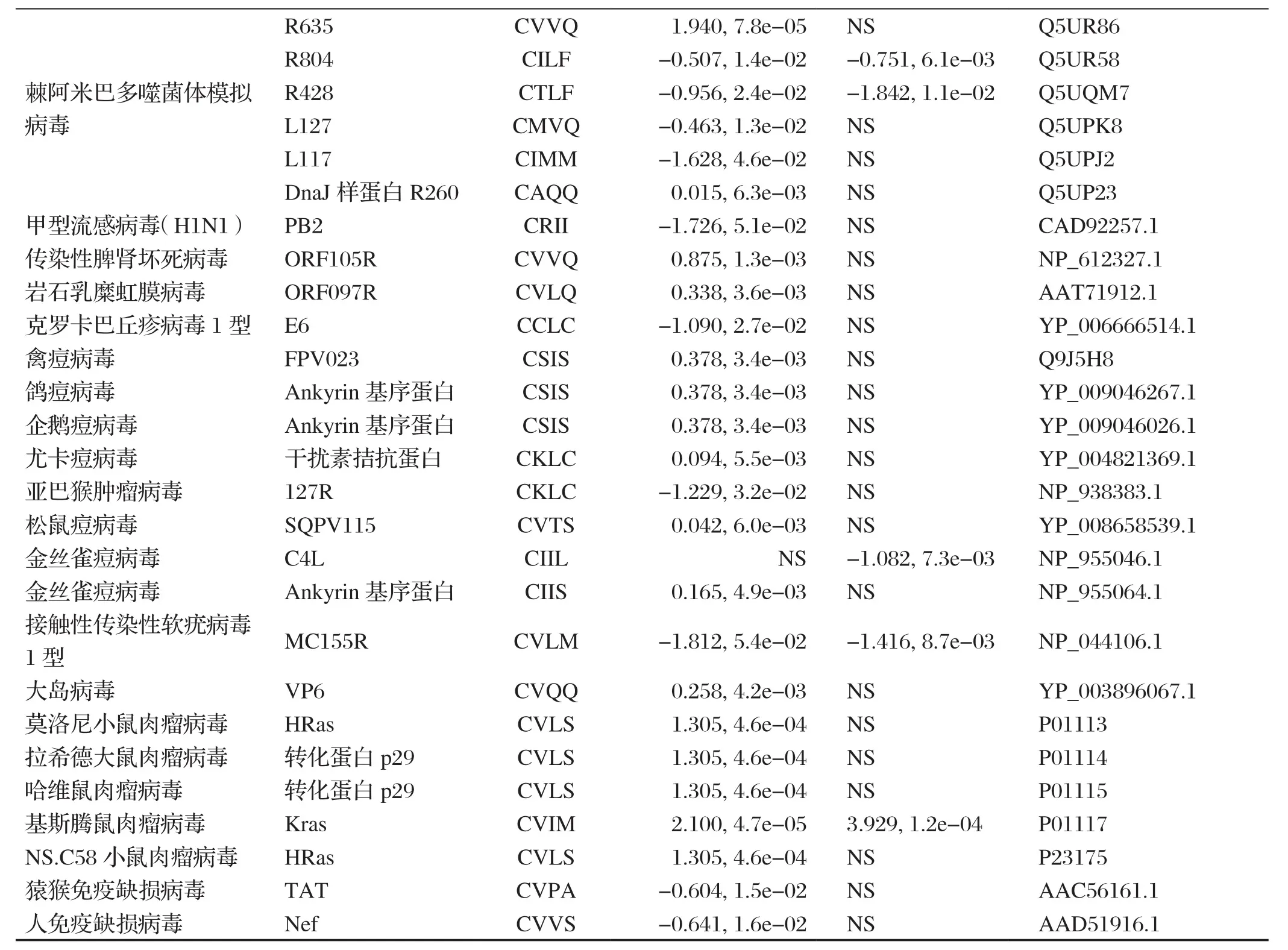
(续上表)
由于研究技术方法的限制,目前已经鉴定的异戊二烯化修饰病毒蛋白较少。早期研究人员通过放射性标记和脂质抗体检测技术证明了丁型肝炎病毒大抗原蛋白HDAg[8]和伪狂犬病病毒US2蛋白[9]是异戊二烯化修饰蛋白。通过应用甲羟戊酸合成酶抑制剂他汀,越来越多的研究发现胆固醇合成通路和中间代谢产物异戊二烯基对病毒至关重要。但大多数研究人员认为,病毒对异戊二烯化的依赖是其通过修饰宿主蛋白影响病毒生存。
与之不同的是,本文预测发现有16个科的人和动物相关病毒蛋白具有很高的置信度可能发生异戊二烯化修饰,尤其是腺病毒科、嗜肝DNA病毒科、疱疹病毒科、巨大病毒拟菌病毒科、虹彩病毒科、痘病毒科和逆转录病毒科的多种病毒蛋白可能发生异戊二烯化修饰,其中又以大基因组病毒最多。本文结合已有的对病毒与异戊二烯化修饰的知识和预测的病毒蛋白修饰结果,重点对与人类健康和畜牧养殖业密切相关的几种病毒和异戊二烯化修饰的关系进行介绍。
2.1 伪狂犬病病毒(Pseudorabies virus,PRV)伪狂犬病病毒编码的衣壳蛋白US2不含跨膜区和定位信号,但在感染细胞中却定位于细胞质膜和胞质囊泡表面。Amanda等[9]证明了PRV US2蛋白的C末端可以发生异戊二烯化修饰,进而通过与Tubulin结合被运输到细胞质膜上,该修饰对其亚细胞定位是必需的。进一步研究发现,在成熟的病毒粒子中US2蛋白是非异戊二烯化修饰的,提示非修饰的US2蛋白在PRV感染过程中结合微管蛋白辅助病毒粒子脱壳并进一步帮助其他衣壳蛋白进入细胞核,而异戊稀基修饰的US2蛋白则发挥不同的功能。本文预测发现,牛疱疹病毒US2蛋白也可能发生异戊二烯化修饰,但其可能是不同于PRV US2的法尼基化,而是偏好亮氨酸的香叶基化修饰。
2.2 非洲猪瘟病毒(African swine fever virus,ASFV)早期研究发现,异戊烯基修饰抑制剂能够有效抑制ASFV的入侵、复制和释放。Quetglas等[16]用异戊烯基抑制剂处理巨噬细胞后发现ASFV入侵效率降低,病毒在巨噬细胞中的复制减少,病毒粒子形态发生受到抑制,大量囊膜前体和未成熟病毒粒子聚集在细胞膜周围,病毒工厂大小减小。研究人员认为,异戊烯基化通过影响宿主细胞小GTPase的异戊二烯化修饰,从而对病毒的生命周期产生影响。有趣的是,ASFV编码B318L是反式香叶基焦磷酸合成酶,能够催化合成GGPP和长链异戊稀基底物[17]。并且,在ASFV感染的细胞中GGPP升高25倍[18]。该基因对ASFV在猪原代巨噬细胞中的复制是必需的。目前尚无研究证实ASFV蛋白能够被异戊烯基化修饰,本文预测到ASFV编码L83L和I9R等病毒蛋白可能是异戊二烯化底物蛋白。而B318L可能是通过合成GGPP底物促进宿主或病毒蛋白的异戊二烯化修饰,从而影响病毒粒子的组装、成熟和释放。
2.3 丙型肝炎病毒(Hepatitis C virus,HCV)研究表明,HCV的有效复制依赖于宿主蛋白FBXL2(F-box蛋白,SCF复合物组成部分之一)的香叶基化修饰。HCV蛋白NS5A能特异性地和香叶基化修饰的FBXL2发生相互作用,且与IP3R3形成三重复合物,从而使IP3R3靶向泛素-蛋白酶体系统降解,抑制IP3R3介导的Ca2+释放和其诱导的细胞凋亡,最终促进HCV的复制[19]。预测发现2~6型HCV的NS5A蛋白C末端是CSMS,可能发生法尼基化修饰(FT值为-1.727,p值为5.1e-02)。NS5A还能和Viperin相互作用,该作用发生在脂滴表面和复制复合体,并且依赖于NS5A的N末端两亲性螺旋和C末端[20-21],表明法尼基化修饰可能在其中发挥重要作用。
2.4 丁型肝炎病毒(Hepatitis delta virus,HDV)Glenn等[8]首先鉴定到HDV的 大抗原蛋白HDAg羧基末端CPRQ基序能够发生异戊二烯化修饰,进而促进HDAg与HBsAg结合和病毒粒子的组装[10-11]。研究表明,CPRQ基序的法尼基化修饰对其核定位、抑制RNA复制和病毒粒子的组装是必需的[12]。法尼基转移酶抑制剂Lonafranib能够显著抑制HDV的组装[13-14]。目前,Lonafranib已经在120多名HDV患者中服用,并且正在进行三期临床试验[15]。值得注意的是,Lonafranib已被美国食品及药物管理局和欧洲药品管理局指定为治疗早衰症的孤儿药。
2.5 小鼠白血病病毒(Murine leukemia virus,MuLV)尽管没有预测到MuLV编码蛋白可以发生异戊二烯化修饰,但Overmeyer等[22]研究发现他汀能够通过抑制Rab1p和Rab6p的香叶基化修饰,抑制了细胞蛋白从内质网向高尔基体的转运系统,使得MuLV前体囊膜糖蛋白gPr90env不能迁移到高尔基体进行酶解加工,最终抑制了病毒粒子的组装。
2.6 人免疫缺陷病毒(Human immunodeficiency virus,HIV)研究表明,他汀能够通过上调CD4+T淋巴细胞中细胞周期依赖性激酶抑制因子p21抑制HIV的感染和复制[23]。他汀也能够抑制HIV-1潜伏感染的U1细胞释放病毒粒子,该作用与他汀抑制了胆固醇的合成无关,而是通过抑制蛋白质香叶基化修饰[24]。当回补GGPP时,能够逆转他汀对HIV-1释放的抑制作用。有趣的是,尽管小GTPase对HIV的复制很重要,然而,预测到HIV编码的Nef蛋白可能发生异戊二烯化修饰。推测异戊二烯化修饰通过促进Nef蛋白靶向反式高尔基体网络劫持多种细胞内转运调控因子,从而减少细胞膜表面的MHC-1类分子[25],利于病毒的感染和生存。值得注意的是,HIV-1的前体囊膜糖蛋白gp160和MuLV的囊膜糖蛋白具有相似的切割位点,并且也依赖从内质网转运到高尔基体上进行切割成熟[22]。因此,推测香叶基化修饰可能通过作用于Rab蛋白促进内质网向高尔基体转运并最终促进HIV-1的囊膜糖蛋白的酶解成熟。
2.7 γ疱疹病毒(Murine gammaherpesvirus 68,MHV68)尽管尚无证据表明γ疱疹病毒编码相关蛋白能够发生异戊二烯化修饰,但Lange等[26]发现MHV68的复制需要胆固醇代谢的中间产物对宿主蛋白进行异戊二烯化修饰,进而促进病毒复制。利用他汀抑制胆固醇合成途径及其中间代谢产物异戊二稀基的产生,能够呈显著抑制MHV68在原代巨噬细胞中的复制和在腹膜腔渗出细胞中的激活。
2.8 巨细胞病毒(Cytomegalovirus,CMV)Potena等[27]研究发现氟伐他汀能够显著抑制CMV在人脐静脉内皮细胞中的复制、成熟病毒粒子的产生、抗原基因的表达和病毒DNA的合成。有趣的是,CMV的复制依赖于NF-κB信号通路,而他汀能够有效阻断CMV诱导的NF-κB的激活。有意思的是,用PrePS程序预测到CMV编码的IRL9蛋白极有可能发生法尼基化修饰(FT值为0.187,p值为4.7e-03)。IRL9蛋白目前功能未知,因此,猜测他汀可能通过抑制该蛋白的法尼基化修饰,最终抑制了NF-κB信号通路和CMV复制。
2.9 腺病毒E1A蛋白大量研究表明,腺病毒E1A蛋白通过其N末端CR结构域与p300和Rb等细胞蛋白结合发挥细胞转化功能,而其C末端则通过结合转录共抑制因子CtBP抑制了E1A蛋白N末端的转化活性[28]。E1A蛋白C末端异戊二烯化修饰可能通过促进其膜定位增强与CtBP的结合,从而限制E1A蛋白的致癌活性。有趣的是,在较古老的腺病毒中E1A蛋白并不能发生异戊二烯化修饰[29],提示腺病毒的异戊二烯化修饰可能是其进化过程中新出现的事件。
3 总结和展望
异戊二烯化作为一种通过共价连接异戊二烯基团到蛋白质上,从而将其靶向到细胞膜结构上的翻译后修饰,其不仅在细胞的正常生命活动的多个过程汇总发挥关键作用,同时也影响了病毒生命周期的多个过程。虽然目前仅有丁型肝炎病毒HDAg和伪狂犬病病毒US2蛋白被证明可以发生异戊二烯化修饰,但预测仍有很多威胁人类公众卫生和健康的病毒以及危害家禽养殖业和畜牧业的动物病毒编码较多蛋白可能发生异戊二烯化修饰。
令人振奋的是,针对HDAg法尼基化的抑制剂Lonafranib对HDV患者具有较好的治疗效果。这提示可通过药物干预病毒蛋白的脂质修饰来防控动物传染病,或可能通过针对相关修饰蛋白上的基序突变开发新型减毒活疫苗等。因此,仍需进一步探究病毒蛋白的异戊二烯化修饰及在病毒各个生命周期过程中发挥的生物学作用。
