若尔盖高原高寒草甸地表能量交换和蒸散研究*
2021-08-25郭小璇姚志生胡正华郑循华
郭小璇,王 凯,李 磊,张 寒,3,马 磊,4,姚志生,张 伟,胡正华,郑循华,5
(1.南京信息工程大学气象灾害预报预警与评估协同创新中心/应用气象学院,南京 210044;2.中国科学院大气物理研究所大气边界层物理和大气化学国家重点实验室,北京 100029;3.天津师范大学地理与环境科学学院,天津 300387;4.卡尔斯鲁厄理工大学气象与气候研究所大气环境研究部,加尔米施-帕滕基兴 82467,德国;5.中国科学院大学地球与行星科学学院,北京 100049;6.河北省气象技术装备中心,石家庄050021)
青藏高原地表过程(如能量平衡、地表蒸散)的机理机制研究及其与大气过程的耦合模拟研究,对于东亚和全球气候变化研究具有重要意义[1-3]。准确可靠的地表过程观测数据,尤其水热变化特征的长期观测资料,是开展上述研究工作的基础。高寒草甸是青藏高原分布最为广泛的生态系统,总面积超过50%,在草地中,高寒草甸占比约41%[4],因此高寒草甸生态系统是高原能量和水分循环的重要组成部分。近年来,一些学者在青藏高原陆续开展了高寒草甸能量交换和蒸散的观测研究,这些研究的观测站点大多数位于海北[5-17]和高原中南部[12,18-20]的温带或亚寒带气候区,也有一些位于高原东部的亚寒带半湿润或湿润区[21-27],以及云南境内的高原东南延伸地带[28-29]。上述研究发现,高寒草甸地气能量交换和地表蒸散过程受气温、辐射、降水和地表因子的共同影响,由于地理位置、海拔和气候带的差异,不同高寒草甸地表能量交换和蒸散的变化特征以及它们对环境因子的响应都存在很大差异。
若尔盖高原地处青藏高原东北边缘,平均海拔3500m,是高原上降水较为充沛的地区之一,植被覆盖状况较好,分布有大面积的高寒草甸生态系统,是青藏高原重要的天然牧场和生态屏障。近40a 来,若尔盖高原气候呈现明显的暖干化趋势,加之人为活动影响,生态环境发生显著变化,使该区域成为近年来高原气候研究的热点[30-31]。因此,有必要通过观测研究深入认识该区域高寒草甸生态系统的地表能量交换和蒸散过程。然而,受高寒地区环境条件的限制,目前在若尔盖地区的地面通量观测资料非常缺乏,仅有一个站点的周年观测报道[22-23]。地面原位观测通量数据的缺乏,会限制陆面模式中上述相关过程参数化方案的发展,影响模式对高原地表能量和水循环过程的模拟能力[2]。此外,基于卫星遥感反演的地面资料(如蒸散),受到冬季地表积雪和夏季局地强烈对流产生的云层的严重影响[32],需要可靠的地面观测数据对其可靠性进行验证。
基于上述背景,本研究以若尔盖高原一处典型高寒草甸作为研究对象,采用涡动相关法开展地表能量交换通量和蒸散的周年观测,旨在揭示该类型草甸生态系统地表能量交换和蒸散的季节变化特征,探讨生物和环境因子对这些过程的影响机制,为准确理解若尔盖高原草甸生态系统地气间水热交换过程提供更充分的观测依据,并为高原陆面模式参数化方案的发展和卫星遥感反演资料的验证提供可靠的地面观测数据。
1 材料与方法
1.1 研究站点概况
研究站点位于四川省若尔盖县红星镇境内的一处高寒草甸(34°03'32"N,102°43'47"E,图1),海拔3231m,地处青藏高原东部的亚寒带湿润气候区。位于研究站点南部约 80km 的若尔盖县气象站1980-2012年的气象观测资料显示,该地区年平均气温为1.6±0.7℃(平均值±1 倍标准差,下同),最冷月出现在1月,平均气温为-9.6±1.6℃,最热月出现在7月,平均气温为11.2±0.9℃,年平均降水量为649±94mm,约 85%的降水发生在生长季。长期(2000-2013年)物候观测表明,该地区生长季通常开始于4月中上旬,结束于10月中下旬[33]。生长季的划分依据为,平均气温连续5d 大于0℃视为生长季的起始日期,平均气温连续5d 小于0℃则为生长季的结束日期。研究样地土壤类型为亚高寒草甸粉沙壤土,主要优势物种为矮嵩草(Kobresia humilis)和干生苔草(Carex aridula)。样地在6月初-7月初被用作早夏牧场,10月中旬-12月下旬则为冬季牧场,其余时间不放牧,放牧家畜主要为高原牦牛和藏系绵羊,放牧强度约为3.3 羊单位·hm-2·a-1。观测期内数据显示,该草甸样地在生长盛期植被盖度约为93%,地上净初级生产力为354.9g·m-2,土壤(0-10cm)有机碳含量为70.4g·kg-1[34]。综合上述指标,研究样地为健康的高寒草甸生态系统[35-37]。
1.2 通量观测方法
观测时间为2013年11月1日-2014年10月31日。涡动相关通量观测系统由一台三维超声风温仪(CAST3,Campbell Scientific Inc.,USA)、一个红外开路CO2/H2O 分析仪(LI-7500,LI-COR,USA)和一个数据采集器(CR5000,Campbell Scientific Inc.,USA)组成,系统供电由太阳能板和蓄电池提供,传感器安装高度为2m,观测塔半径200m 内下垫面平坦均一。CR5000 上还连接有其它探头,每半小时测量总辐射(Rs)、净辐射(Rn)、表层(5cm)土壤温度(Ts)、不同深度(5、10、20 和40cm)土壤体积含水量(VWC)和土壤热通量(G)等气象环境要素。由于其它试验安排和设备限制,在试验样地西南方向3km 处的一块湿地样地上,设有一套微型自动气象站,为本研究提供气温(超声风温仪无有效数据期间)和降雨量数据。降雪量的观测点位于样地西北方10km 处的郎木寺镇,由实验人员手动采集获取。基于观测得到的10Hz 涡动数据,根据式(1)、式(2)[38]计算半小时平均感热通量(H)和潜热通量(LE)。
1.3 数据处理
(1)数据质量控制
数据前处理和通量计算的步骤包括:原始涡动数据去野点[39];坐标轴二次旋转[40];最大协方差法矫正风速和密度数据间的时间差;平均去趋势法计算脉动;利用式(1)和式(2)计算半小时平均通量;通量的高频和低频衰减矫正[41];水热密度效应矫正[42]。上述计算步骤由EddyPro 软件(6.2.0 版本,LI-COR Biosciences,美国)完成。
通量数据的质量控制步骤包括:剔除仪器维护和故障及降水期间的通量(LI-7500 输出AGC 值大于65);删除平稳性检验和湍流发展状况检验[43]综合等级为2,以及夜间摩擦风速小于0.1m·s-1的半小时通量;删除小于-20W·m-2的潜热通量;以一天为时间窗口,删除偏离平均通量大于3 倍标准差的半小时通量。
(2)数据插补
为估算季节和年尺度的累积通量,按以下方法插补缺失的半小时通量:≤2h 的空缺,用其两端的有效通量进行线性内插;大于2h 小于4d 的空缺,用邻近前后各5d 相同时段有效通量的平均值进行填补(平均日变化法);大于4d 的空缺,多由仪器或供电故障导致,选取空缺邻近时段的观测数据,建立感热通量与净辐射、潜热通量与气温的线性拟合关系,采用模拟值对日值进行插补。
(3)蒸散量计算
根据潜热通量和水的汽化潜热(λ=2.45MJ·kg-1),利用式(3)计算得到植被的实际蒸散量(ET)。为评价蒸散是否受到水分限制,引入Priestley-Taylor 系数αPT,其数值为ET 与平衡蒸散量(ETeq)之比,ETeq反映在没有水分限制及没有水平和垂直对流情况下,由有效能量所决定的最大蒸散量[44]。
式中,γ 是干湿表常数(kPa·K-1),Δ 是饱和水汽压随温度变化曲线的斜率(kPa·K-1)。
(4)能量闭合率(EBR)和波文比(β)计算
(5)地表参数计算
地表反照率定义为出射和入射太阳短波辐射之比。在计算反照率的日平均值时,只取10:00-16:00时段内的半小时数值,以去除太阳高度角变化对计算结果的影响。
避免异常值干扰,在计算地表导度(gc)、空气动力学导度(ga)和地表-大气解耦系数(Ω)的日平均值时只选取10:00-14:00 时段且无降水的半小时数值。
地表导度(gc,m·s-1)是描述水汽经由地表(包含土壤、冠层空间和叶片气孔)进入大气的能力的最常用参数[45],可以利用实际观测的 ET 和对Penman-Monteith 公式[46]进行变换来计算gc,即
式中,ga为空气动力学导度(m·s-1)。
空气动力学导度(ga,m·s-1)的大小取决于下垫面的空气动力学特性和大气湍流强度,计算方法为[46]
式中,U 和u*分别为水平风速和摩擦风速,单位均为m·s-1。
解耦系数(Ω)用于描述地表与大气之间的耦合程度[47]。当两者完全不耦合时(Ω = 1,ga<< gc),ET 只受有效能量(太阳辐射)控制。当两者完全耦合时(Ω = 0,ga>> gc),ET 只受gc等土壤表面导度因子和VPD 控制,而不受辐射控制。Ω 的表达式为
2 结果与分析
2.1 观测期环境因子变化
由图2a 可见,观测期内气温(Ta)平均为3.0℃,最低和最高日平均气温分别为-15.3℃和18.0℃,分别出现在2013年12月16日和2014年7月21日(图2a)。根据前文给出的划分依据,选取2014年4月8日-10月31日为生长季,其余时间段为非生长季。生长季和非生长季的平均气温分别为9.7℃和-3.7℃。表层(5cm)土壤温度(Ts)的动态变化与气温相似,年平均值为6.3℃,日平均最低值和最高值分别为-4.2℃和19.3℃,土壤冻结期为12月上旬-翌年3月中旬。图2b 显示,观测期内年总降水量(PPT)为682.7mm,高于若尔盖县气象站33a 平均降水量(649.0mm),降水的季节分布不均,84%发生在生长季,日最大降水量为33.3mm·d-1(8月8日)。饱和水汽压差(VPD)与气温有相似的季节变化特征,但波动幅度比气温大得多,日最大值为0.98kPa,出现于7月25日。图2c 表明,表层(5cm、10cm 和20cm)土壤受冬季冻结影响,体积含水量(VWC)从11月开始陆续大幅下降,并稳定在0.1 左右,40cm土壤VWC 则变化较小。随着土壤解冻,土壤中液态水含量增加,各层VWC 从3月中上旬开始逐渐增加,表层土壤增加最为迅速,其随后的变化与降水事件有较好一致性。6-7月降水量显著低于多年平均水平,对土壤水的补充不足,20cm 以上VWC 呈现持续下降趋势,而40cm 的VWC 未有显著变化。8-9月降水事件频发,VWC 开始缓慢增加,9月中上旬开始,20cm 和40cm 土壤VWC 迅速增加,并超过5cm 和10cm 的VWC,且持续到观测期结束。由图2d 可见,总辐射(Rs)呈现春夏高、秋冬低的季节变化趋势,生长季平均值为217.3W·m-2,年平均值为194W·m-2,总量为6118MJ·m-2。
2.2 观测期地表参数变化
由图3 可见,冬季地表反照率显著高于其它季节,频繁出现脉冲式增加,受降雪和地表积雪影响,全年最大值为0.80,出现于2月17日(图3a)。秋冬季的无积雪时段,反照率平均值约为0.20。随着春季植被萌发生长,覆盖度增加,反照率逐渐下降,夏季达到全年最低值,平均约0.16。地表反照率全年平均为0.22,与海北和玛曲高寒草甸观测结果相近[22,48]。
地表导度(gc)全年平均为8.13mm·s-1,并表现出显著的季节变化(图3b)。非生长季变化幅度较小,平均为3.16mm·s-1。进入生长季后gc显著增加,但在5月中旬达到第一个高峰后持续下降趋势,于7月下旬达到最低点;随后gc迅速增加,于8月上旬达到全年第二个峰值,进入秋季后,又呈下降趋势,生长季平均值为11.67mm·s-1。相比gc,空气动力学导度(ga)则无显著的季节变化,其数值显著大于gc(P<0.01),全年平均为20.96mm·s-1。解耦系数(Ω)表现出与gc相似的季节变化趋势(图3c),全年平均为0.43。非生长季平均值为0.23,冬季降雪后,Ω值出现小幅增加。4月中旬,Ω 迅速上升,生长季平均为0.57,有两个峰值,分别出现在5月和8月。
2.3 观测期能量交换分析
2.3.1 能量通量的季节变化
图4 给出了经过插补后的日平均净辐射通量(Rn)、感热通量(H)、潜热通量(LE)和土壤热通量(G)的季节动态变化。净辐射和土壤热通量为正代表能量向下传播,感热和潜热通量为正代表能量向上传播。感热通量的有效数据比例为67%,由于数据存储故障导致的数据缺失率约为13%(2013年11月22-26日和2014年3月19-30日)。潜热通量有效数据比例为55%,其中,数据存储故障和LI-7500故障(2014年1月26日-3月30日)共同引起的数据缺失约占27%。
净辐射通量(Rn)在春夏季的波动显著大于秋冬季,日平均值变化范围为8.3~218.8W·m-2,年平均值为94.5W·m-2,总量为2980.2MJ·m-2(图4a)。土壤热通量(G)的量级最小,年平均值为1.2W·m-2,季节变化趋势为冬季低、夏季高,日平均值的极值为-20.0 和29.5W·m-2;2月末G 开始由负转正,大气开始向土壤输送热量,随后即伴随着表层土壤的融化(图4a);10月中旬之后,G 完全变为负值,即土壤向大气传输热量。
感热通量(H)日平均值的极值为-10.7 和60.6W·m-2,年平均为21.0W·m-2,仅在冬季个别时段为负值,其它时间均为正值(图4b)。非生长季,H 和潜热通量(LE)都处于全年最低水平,且两者量级接近(表1)。3月中旬,随着地表接收到的能量增多,H 缓慢增加,于5月初出现全年第一个高峰。随后,由于植被迅速生长,土壤的热源效应减弱,H的强度逐步下降。7月下旬,H 迎来全年第二个高峰,这一时期H 升高主要有两个原因。首先,6-7月降水偏少,辐射很强,土壤含水量和热容量有所下降,地表的热源效应得以加强,这与一些学者在半干旱草原[49]和高寒草甸[6,23]生态系统上获得的观测结果类似;其次,6月中旬-7月中旬,样地被用作为夏季牧场,牲畜啃食使地上生物量有所减少,影响地表反照率和表层土壤容重,也在一定程度上增加了H。进入8月后,迟来的降水改变了地表能量分配状况,H 迅速减小,至观测结束,H 变化幅度较小。
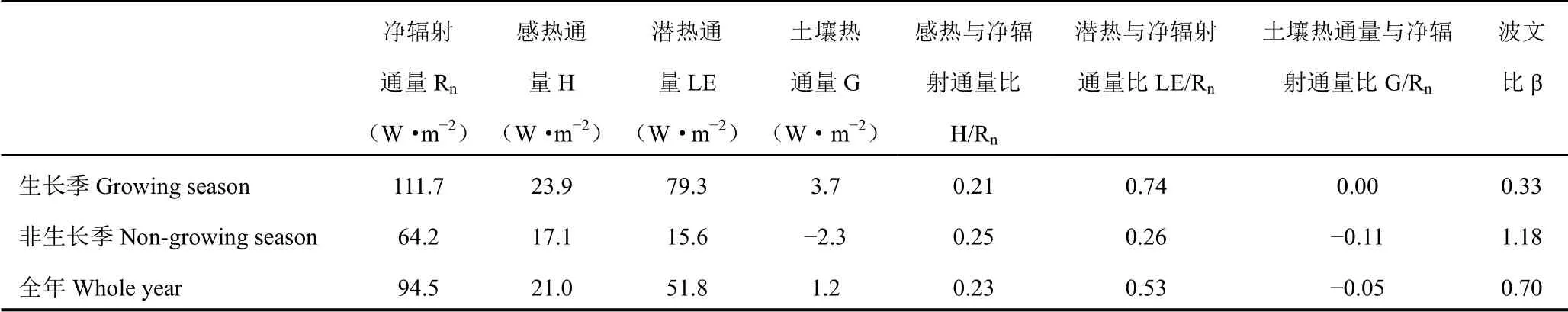
表1 各观测时期能量通量平均值、能量分配和波文比Table 1 Means of energy fluxes,energy partitioning and Bowen ratio during different measurement periods
LE 的季节变幅大于H,日平均值范围为3.4~144.3W·m-2,年平均值为51.7W·m-2(图4),生长季LE 显著高于非生长季,也显著大于H(表1)。3月之前,LE 的日平均值都在30W·m-2以下。4月开始逐渐增加,并超过H,于6月2日达到全年的峰值,表明LE 开始主导向大气的能量传输。两个方面影响了这个时期LE 和H 的相对变化:其一,草甸植被开始萌发新芽,大气短波辐射对地表的加热效应被逐渐削弱,土壤的热源效应相应减弱;其二,叶面积指数增加,蒸腾作用逐渐增强,同时由于气温升高、降雨充沛等因素,土壤的蒸腾也迅速增加[50-52]。受季风影响,6-8月是若尔盖高原地区降水最多的月份,本应是一年中潜热通量(蒸散)最强的季节,但在6月初之后,LE 并未继续增加,其强度一直维持在5月的水平(图4b),这是由于6-7月的降水量仅为多年(1980-2012年)平均水平的52%,降水亏缺导致这两个月的LE 强度受到抑制。8月后,LE稍有增加,但变化趋势平稳。9月之后,随着气温继续下降,植被逐渐停止生长,LE 和蒸散强度随之减弱。至10月末,LE 和H 的量级又趋于接近。
2.3.2 能量通量的日变化
图5 显示,能量通量各组分在生长季和非生长季均表现出“单峰型”的日变化趋势。生长季和非生长季内所有日期能量通量日变化平均值均表现为日出后逐渐增大、午后达到日最大值的规律,但出现时间不一,Rn和H 为13:00 左右,LE 为14:00左右,而G 的日变化相对滞后,在16:00 后才出现极大值。随后各项指标逐渐降低到日出前的水平,但出现的时间也不同。在生长季,H 全天都低于LE,在非生长季,H 只在白天时段略高于LE,其它时段则相反。
2.3.3 能量通量与环境因子的关系
对H 和LE 与环境因子分别进行线性回归分析,结果见表2。由表可见,不管哪个时期,辐射(尤其是Rn)都是H 最主要的控制因子,其次为VPD,温度(Ta和Ts)只在非生长季与H 存在显著相关关系。对于LE,最重要的影响因子都是温度和Rn,次要影响因子是VPD。有研究[53-54]发现,风速大小显著影响LE 的强度,与本研究结果不一致,这是因为本研究用于回归分析的是日平均风速,它在季节尺度上无显著的变化趋势。VWC 只在生长季对LE 有显著的影响作用,由于冬季表层土壤冻结,基于频域反射原理获取的土壤含水量数据失真,所以未对非生长季数据进行分析。
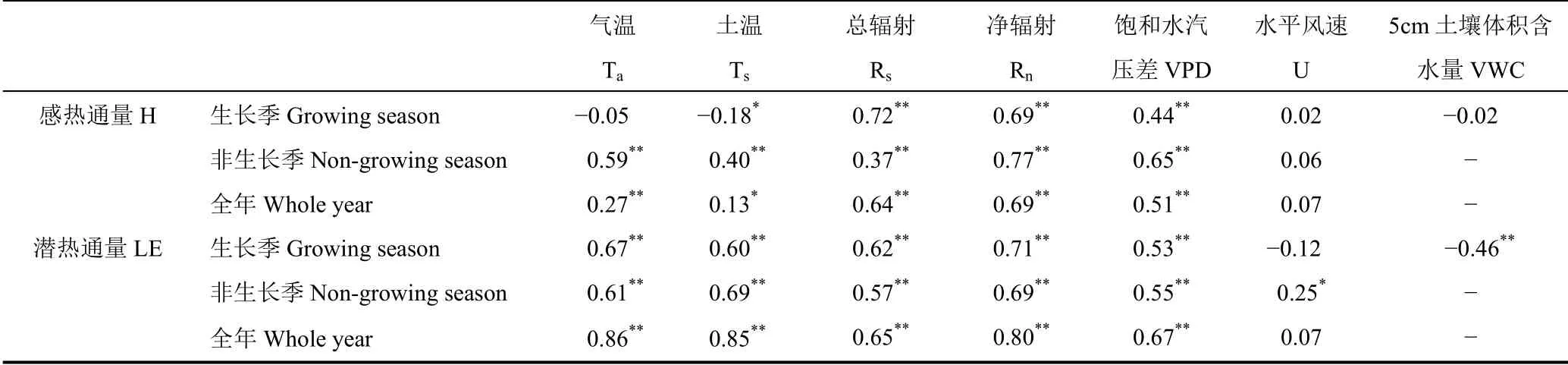
表2 感热通量和潜热通量与环境因子的线性相关系数Table 2 Linear correlation coefficients between the sensible heat flux(H)and latent heat flux(LE)and the environmental factors
Rn是生长季能量通量的主要控制因子,为研究不同干旱级别下它对能量通量的影响,将VPD 划分为不同等级,将每一等级的H 和LE 与Rn进行线性相关分析。如表3 所示,在湿润环境下Rn对H 的影响最大,相关系数最高,随着VPD 增加,两者的相关性逐渐变小,当VPD>0.8kPa 时,两者无显著相关关系。LE 对Rn的响应不受VPD 变化而变化,当VPD为0~0.8kPa 时,两者始终显著相关,只有在极干旱情况下(VPD>0.8kPa)才表现为不相关,但这一结果可能受样本量较小的影响。

表3 生长季不同饱和水汽压差水平下感热通量和潜热通量与净辐射通量的线性相关系数Table 3 Linear correlation coefficients between the sensible heat flux(H)and latent heat flux(LE)and the net radiation under different levels of vapor pressure deficit(VPD)during the growing season
2.3.4 能量分配
表1 列出了各个时期能量通量的大小、能量分配状况和波文比的平均值。各项能量通量在生长季都显著高于非生长季。净辐射作为所有能量通量的来源,全年分配给感热通量的比例(H/Rn)为23%,非生长季的H/Rn大于生长季。相反,分配给潜热通量的比例(LE/Rn)则是生长季更高,为74%,全年平均为53%。土壤热通量与净辐射之比(G/Rn)全年平均值为-0.05。波文比β 取决于H 和LE 能量分配比例的变化,全年平均值为0.74,生长季平均值(0.33)显著低于非生长季(1.18)。
由图6 可见,逐月能量通量平均值也表现出与表1 类似的趋势(图6)。H/Rn和LE/Rn月平均值的变化范围分别为0.16~0.33 和0.20~0.91,在非生长季,H 的能量分配比例略大,但总体与LE 相当,从4月开始,LE/Rn开始大于H/Rn,并持续到10月,表明LE 在这一时期占绝对主导地位。G/Rn月平均值在-0.45~0.12,最小值和最大值分别出现在12月和3月,分别与表层土壤冻结和融化时间一致(图2a)。β 也呈现冬春高、夏秋低的规律;11月-翌年3月,β 大于1,4-10月,β 远小于1,月平均值的极大值和极小值分别出现在1月和8月,分别为2.04 和0.18(图6)。β 与环境因子的相关性表明,土壤温度是β的首要调控因子,尤其在生长季,两者相关系数达0.65(P < 0.01)。
根据式(6)得到全年、生长季和非生长季的能量闭合率,平均值分别为0.77、0.85 和0.44,冬季能量闭合率较低,全年结果与全球FLUXNET 站点报道的能量闭合率的平均值(0.79)较为接近[55]。
2.4 观测期地表蒸散变化
地表蒸散表现出显著的季节变化(图7),生长季 ET 显著高于非生长季,年平均蒸散量为1.82mm·d-1,变化范围为0.12~5.09mm·d-1,最小值和最大值分别出现在12月22日和6月2日。7日滑动平均线更直观呈现了蒸散的季节变化趋势,12月和1月为全年最低,平均蒸散量约为0.35mm·d-1,2-3月间缓慢增加;进入生长季后,ET 迅速增加,4月和5月的平均值分别达到1.81 和2.87mm·d-1;进入生长旺季(6-8月),ET 保持较大的速率,平均值为3.40mm·d-1;进入秋季后ET 呈逐步下降趋势,9月和10月的平均蒸散量分别为2.56 和1.73mm·d-1。
蒸散包括土壤蒸发和植被蒸腾,受空气动力过程和生物过程的共同作用[56],这些过程的相互关系可用导度参数ga和gc以及解耦系数Ω 来解释。在非生长季,蒸散过程由地表蒸发主导,ga显著大于gc,Ω 平均值为0.23(图3),表明ET 更多受gc等土壤表面导度因子的控制,有效能(太阳辐射)对ET 的影响很小。进入生长季后,Ω 显著增加,多数时间都大于0.5,表明辐射主导了蒸散过程,gc的控制变为次要因素。7月底,Ω 出现低谷,原因是降水量少,草甸生态系统处于水分匮缺状态,增加了地表和大气间的耦合程度。
Priestley-Taylor 系数全年平均值为0.75,变化范围在0.20~1.61,生长季显著高于非生长季(图7)。αPT的值越接近1,代表实际蒸散速率与平衡蒸散速率相近,蒸散不受水分限制[14,44,57]。冬季,αPT的平均值在0.5 左右,降雪发生后有小幅增加。随着雨季来临,αPT迅速增加,在生长季旺盛期,其数值在1上下波动,尽管6-7月由于降水偏少出现下降趋势,总体而言,这个时期草甸生态系统的水分供应相对充足,这有赖于前期较大的土壤储水量,也说明这一地区草甸生态系统发挥着重要的水源涵养作用,同时也表明蒸散在年尺度上的变化主要取决于年降水量的季节分布。
2.5 观测期水分收支分析
高寒草甸生态系统水的循环过程主要包括降水、蒸散、径流、土壤储水、地下水补充等,各项分量动态变化并达到水分收支的平衡。本研究通量观测的区域平坦,可忽略地表径流的输入和输出,降水和蒸散分别成为最大的输入项和输出项。
图8 给出了蒸散量和降水量的累积变化,观测期的绝大部分时间内(2014年9月中旬之前),累积蒸散量始终高于累积降水量。11月-翌年5月上旬,两者差值保持在30mm 以内,这个时期降水少、气温低,加之大部分时间表层土壤处于冻结状态,可用于蒸散的水分主要来自降雪和少量的土壤水。6-7月,蒸散的增长速度明显加快,两者差值拉大,于7月27日出现最大值(130.5mm),这两个月的降水量远低于多年平均水平,草甸生态系统处于水分匮缺状态,只能通过消耗表层土壤中储存的水分来维持较大的蒸散量;这一时期,各层土壤含水量都呈下降趋势,表层土壤的降幅超过深层土壤(图2c),表明随着时间推移,蒸散过程对土壤水的获取逐渐往深层发展。8-9月,降水频率和强度显著增加,并显著高于多年平均水平,累积ET 和累积P 的差异逐渐减小,至10月初,累积ET 被反超,周年观测结束时,两者分别达到665.8mm 和682.7mm,基本持平,比值(ET/P)为0.98。从水分平衡角度看,草甸生态系统接收到的降水,基本都以蒸散的方式返回大气。
8月之后,各层土壤的含水量因降雨得以补充,呈上升趋势,至周年观测结束时,0-50cm 土壤储水量较观测初始多45.0mm,减去年降水量与蒸散量之差,还有28.1mm 的盈余,由于观测样地的地下水埋藏很深,这部分盈余水量可能来自更深层(50cm以下)土壤在缺水期(6-7月)对表层土壤的补给。此外,还发现生长季日蒸散量与降水量之差(ET-P)与表层土壤(5cm)体积含水量之间存在极显著的负相关关系(P< 0.01),再次表明在干旱季节,土壤储水是高寒草甸蒸散的重要水源,在天山[58]和黑河流域[15]的高寒草甸生态系统也观测到类似的结果。
3 结论与讨论
3.1 讨论
通过搜集青藏高原高寒草甸生态系统开展的已有观测研究,发现共有10 个不同下垫面站点、24 个周年的观测数据[8,11,14-16,18,21,23,25-27],与这些研究相比,能量通量各分量都表现出相似的季节变化趋势,生长季潜热占主导,非生长季感热稍占优势,但在年蒸散量、能量分配和波文比的变化等方面存在一定差异。
上述高寒草甸通量观测研究的年蒸散量为470.6±60.8mm(平均值±标准偏差,下同),降水量为506.8±81.5mm,蒸散与降水之比(ET/P)为0.96 ±0.21。本观测研究所得的全年累积蒸散量(665.8mm)和降水量(682.7mm)分别比上述多站点平均值高41.5%和34.7%,同时,这两个数值也是所有高原站点周年观测中最大值。尽管如此,与其它站点相似,ET/P 接近1,表明青藏高原草甸生态系统的地表蒸散过程在年尺度上受水分限制,降水基本都通过蒸散形式返回大气。本研究站点观测周年的平均气温为3.0℃,高于上述多站点平均值(-1.0 ± 1.8℃),平均气温更高,意味着生长季更长,这是本研究蒸散量偏高的重要原因之一。
波文比的全年平均值为0.70,显著低于上述其它观测研究所报道的数值[5,8,11,16,23]。生长季大的蒸散速率导致这一时期波文比仅为0.33,潜热占绝对主导地位。非生长季H 和LE 强度很小,但数值接近,波文比仅为1.18,而其它研究中,尽管非生长季H和LE 量级也很小,但H 的数值经常几倍于LE,导致很大的波文比。其原因可归于本研究站点较高的土壤含水量和较低的地表反照率,尤其在生长季,反照率显著低于其它健康或退化高寒草甸的观测结果[6,11,16,23],致使地表接收到更多的辐射能量以分配给潜热,从而影响能量分配和波文比。陆面模式中能量分配的参数化方案是决定模式性能的重要因素[59-60],不同年份间气候和水分条件存在很大差异,并显著影响能量分配的季节变化特征,一周年的观测数据无法描述其响应规律,今后有条件可以开展多年连续通量观测研究,揭示能量交换和地表蒸散的年际变化特征,以及它们对不同气候和水分条件的响应,这些资料将为模式参数化方案提供可靠的基础数据。
放牧对草甸生态系统的能量交换和蒸散也会产生一定影响,放牧会改变土壤容重和地上生物量,从而改变土壤温湿度、地表粗糙度、地表导度、空气动力学导度等参数[61]。本研究中,在水分匮乏的6月初-7月初,草甸作为夏季牧场,牲畜踩踏会减小土壤孔隙度,减小地表导度,不利于土壤水分蒸发。同时,由于草场载畜量较合理,放牧时间不长,牲畜啃食并未显著改变植被覆盖度,一定程度的啃食也可降低植被的蒸腾需水,有利于土壤水分的储存。本研究还强调了土壤储存水对草甸生态系统水分收支的调节作用,采用模式描述和模拟高原草甸生态系统水循环时,必须充分考虑土壤水分迁移等过程。此外,本研究观测期间没有叶面积指数和生物量的动态监测,卫星遥感产品数据可作为有效的代用数据,用于进一步研究地表植被状况与能量分配、蒸散等过程的相互作用[62]。
3.2 结论
高寒草甸地表能量通量各组分呈明显的日变化和季节变化特征,辐射是感热通量的主要气象影响因子,潜热通量则受温度、辐射和饱和水汽压差共同影响。非生长季的蒸散过程主要受土壤表面导度因子控制,生长季则由辐射主导,土壤和植被表面导度因子的控制变为次要。在季节尺度上,蒸散的变化取决于降水分布,生长季降水匮乏会抑制蒸散,此时土壤储水成为蒸散的主要水源,对草甸生态系统水分收支起到调节作用。从全年来看,降水基本都以蒸散的方式返回大气。与青藏高原上同类观测研究相比,地表能量通量和蒸散都有相似的季节变化趋势,但观测到的波文比和蒸散量为最大值,这是气温、降水、地表植被等因素共同作用的结果。本研究为高原地区陆面模式参数化方案的发展和卫星遥感反演资料的验证提供了可靠的地面观测数据,下一步工作要关注地表通量的长期观测研究,并结合卫星遥感等其它观测方法,揭示高寒草甸能量交换和蒸散过程的年际变化特征及它们对不同气候和水分条件的响应。
致谢:衷心感谢中国科学院大气物理研究所的王晓龙及实验站其久甲先生和阿当先生在野外观测中所提供的帮助。
