绢蒿荒漠粪甲虫多样性特征及其对粪便内种子的二次分配
2021-08-21曹佳敏郭亚亚李娜娜孙海荣车昭碧鲁为华
曹佳敏,郭亚亚,李娜娜,孙海荣,车昭碧,鲁为华,2*
(1.石河子大学动物科技学院,新疆 石河子832003;2.省部共建绵羊遗传改良与健康养殖国家重点实验室,新疆 石河子832003)
牲畜粪便是天然草地放牧生态系统中重要的养分来源之一[1]。放牧家畜通过消化只能吸收利用所采食牧草营养成分的一部分,其余60%~90%的养分以粪便和尿液的方式返还到草原生态系统[2],粪便在草地上的堆积能够有效提高粪堆下土壤水分、有机质、氮、磷和钾的含量,降低土壤pH值,利于周围植物生长[3],但是草食动物新鲜粪便质地黏稠,如无食粪昆虫参与分解,粪便破碎化和分解速度缓慢,会造成粪便下植物缺乏氧气和光照而死亡,同时,粪便堆积导致粪便下植物生长受阻,甚至会波及粪堆外围3~5 cm处,造成粪便下及其周边范围内的草地生产力降低,并且导致牲畜拒绝取食粪便周围的牧草[4-5],据统计,家畜排泄物占据放牧地30%~40%的面积[6],如果粪便不能迅速分解,会损失大片的放牧区域,降低草地放牧利用率,因此,排泄到草地上的粪便必须及时分解才能实现草地的有效保护利用,同时实现养分的正常循环。
粪甲虫作为食粪动物最大的类群,主要包括金龟科(Scarabaeidae)、蜉金龟科(Aphodiidae)和粪金龟科(Geotrupidae),同时,根据粪便处理方式将粪甲虫分为4个功能类群:a)远生型粪甲虫(roller)将粪便团成球状,滚到一定距离后掘洞并把粪球埋到洞中,作为食物球或孵化球;b)外生型粪甲虫(tunneler)直接在粪便下掘洞,将粪便搬运到洞中作为食物球,或产卵于其中,作为孵化球,成为后代繁殖场所和食物来源;c)内生型粪甲虫(dweller)直接在粪中取食,一旦找到粪便便一头扎进粪便中,其整个生活史中的卵、幼虫和蛹发育都发生在粪便中[7];d)偷窃寄生型粪甲虫(kleptoparasites)使用其他粪甲虫的资源,寄生于粪球中。粪甲虫通过取食和搬运改变粪便内部形状,使其内部穿有小孔,加速粪便内部空气流通,从而有利于微生物的取食和粪便的自然分解,微生物对粪便分解主要是对液体部分进行采食和养分的释放,此外,大型食草动物对粪便的踩踏有利于粪便破碎使得碎屑进入土壤,进而加速粪便分解,粪便经过分解进入生态系统,从而参与生态系统的物质循环和能量流动[8-10]。尽管不同功能类群的粪甲虫对粪便处理方式不一样,但最终能够快速改变粪便的形状,使其破碎化,加快粪便的分解速度[11]。
草食动物在采食植物营养体的同时,会将成熟的种子一同吞咽并通过消化道后随粪便排放到草地上,从而实现种子的消化道传播(endozoochorous)。粪甲虫是绢蒿荒漠家畜粪便的主要分解者,在扮演分解者这一角色的同时,它们又会对粪便内的种子进行二次传播。因此,粪甲虫会决定粪便内种子的命运和去向。由于粪甲虫有多个种类和生态功能类型,并且其种类和数量在时间和空间上存在特定的变化规律,这种变化规律又会对粪便的分解及粪便内种子的二次传播产生影响,在人为建立起来的绢蒿荒漠草地放牧系统中,由于有严格的季节选择性[12],使草食家畜密集性地集中在秋季放牧采食、粪便排泄,这种高强度、密集性的放牧过程显然也会影响草地植物种子的传播[13],此外,在组成相对简单的荒漠生态系统中,不仅粪便的分解过程强烈依赖于粪甲虫,而且粪便内种子的再分配也由于缺乏其他有效的动物传播媒介而对粪甲虫有强烈的依赖性。本研究主要通过不同放牧季节(主要是春秋两季)的野外调查和模拟控制试验,以期阐明:1)不同季节绢蒿荒漠的粪甲虫的种类和数量特征及其数量变化规律是什么?2)各类粪甲虫对粪便的进驻和取食特征如何?3)通过模拟试验,探讨种子在粪便内被粪甲虫二次分配的特征。
1 材料与方法
1.1 研究区概况
研究地点选取石河子紫泥泉种羊场的典型绢蒿荒漠放牧地(一大滩)(E 85°49'-85°58',N 43°26'-45°20'),分布海拔980~1120 m。该牧场为重要的春秋牧场,放牧时间包括4-6月(春季)、9-11月(秋季)两个时间段。属于典型温带荒漠干旱气候,年平均温度8℃,月平均最高温度集中在7月,为26.4℃。年降水集中在4、5、8月,年平均降水量180~200 mm。土壤为淡栗钙土。植被组成以伊犁绢蒿(Seriphidium transiliense)为建群种,伴生种有草原苔草(Carex liparocarpos)、羊茅(Festuca ovina)、新疆针茅(Stipa sareptana)、骆驼蓬(Peganum harmala)以及一年生草本角果藜(Ceratocarpusarenarius)和猪毛菜(Salsola collina),植被结构组成较为复杂。
1.2 试验设计及样品采集
1.2.1 粪甲虫季节性群落组成和多样性分析 研究时间为2019年4-10月,研究区为典型的绢蒿荒漠放牧地,为了尽可能覆盖试验区域,保证在绢蒿荒漠中对物种多样性调查的准确性。利用GPS全球定位系统,在空间上将放牧地按海拔每升高20 m为一个调查梯度设置8个不同的海拔。分别为980、1000、1020、1040、1060、1080、1100和1120 m。在4-9月中每个月连续5 d对放牧区域进行调查。调查时首先在放牧区域内的每一调查梯度上设置垂直于调查梯度的50 m×50 m的3个重复样地,在每个样地内设置10个5 m×5 m的样方,对每个样方内马、牛的粪便类型、单位面积重量进行计算、统计,然后分别对不同家畜粪便内及粪便下10 cm深土壤内所存在的粪甲虫种类进行手捡、记数,制作标本带回实验室,通过《中国北方常见金龟子彩色图鉴》[14]和李久文[15]的方法进行分类鉴定。
采用计数法对粪便中粪甲虫的数量进行确定,将粪甲虫装入不密封的管中饥饿12 h排除体内残渣,在水浴锅中加热,待虫子僵直后取出放置于无水乙醇中,置于4℃冰箱中保存,用体式显微镜对粪甲虫进行拍摄并测量记录体尺数据。用形态学分类法对粪甲虫进行分类和鉴定。
1.2.2 粪甲虫进驻粪便行为观察 粪甲虫的行为学研究在典型绢蒿荒漠放牧地进行,时间为6、10月。在清晨收集牛的新鲜粪便,粪便收集后进行充分搅拌混匀。每份粪便为500 g,设置在直径为30 cm的圆形区域,间隔2 m各6份,共需等量粪便12份。在9:00将粪便放置在指定区域,同时开始用摄影机进行连续拍摄观察。从9:00开始,每隔15 min对环境条件进行记录,包括温度、湿度、风速、光照强度。试验开始后,在2、6、12、24、48 h时采用全收粪法收集前5份粪便,直至粪便内再无任何粪甲虫个体存在,将最后一份粪便小心回收装袋。
通过观察,明确粪甲虫的到达时间、到达顺序以及不同的粪甲虫是如何处理粪便的。结合不同时间段对粪甲虫的收集,确定粪便中粪甲虫的数量高峰。
1.2.3 粪甲虫进驻对粪便内种子的分配 用圆形塑料珠作种子模拟物[16]。种子模拟物比真实种子更可取,这样可以防止粪便中的种子与自然存在的种子相混淆。使用了3种不同直径的珠子:小(1.3~1.9 mm;每重复n=100),中(3.3~3.7 mm;n=40)和大(4.8~5.8 mm;n=5)。试验当天,在研究区放置500 g新鲜粪便,其中带有嵌入的种子模拟物。粪便沉积至无粪甲虫活动采集土壤样品。在每个采样点,在以粪便为圆心半径为20 cm的圆形区域内以0~1 cm、2~5 cm、5~10 cm深度对土壤层进行采样。通过筛分土壤样品对珠子进行计数,以计算在不同深度被粪甲虫掩埋的珠子的比例。本研究认为未收回的珠子是被粪甲虫传播垂直距离大于10 cm或水平距离大于20 cm。
1.3 数据处理
利用Excel软件计算多样性指标Margalef丰富度指数(d Ma)、Simpson指数(D)、Shannon-Wiener指数(H')、Pielou均匀度指数(J)。
2 结果与分析
2.1 粪甲虫季节性群落组成和多样性分析
在8个海拔梯度覆盖整个研究区采集到食粪昆虫14797头。其中捕获粪甲虫分属3科17种,蜉金龟科9种:黄缘蜉 金龟(Aphodius sublsmbatus)、黑蜉金龟(A.breviassulus)、血斑蜉金龟(A.haemorrhoidalis)、红亮蜉金龟(A.impunctatus)、黑格(三斑)蜉金龟(Aphodiidae nigrotessellatus)、直蜉金龟(A.rectus)、蜉金龟科一种(Aphodiidae sp.1)蜉金龟科一种(Aphodiidae sp.2)、蜉金龟科一种(Aphodiidae sp.3);金龟科7种:小驼嗡蜣螂(Onthophagusgibbulus)、独行嗡蜣螂(O.solivagus)、三开蜣螂(Copristripartitus)、北方 蜣 螂(C.lunaris)、粗 纹 弯 边 蜣 螂(Gymnopleurus flayellates)、金龟科一种(Scarabaeidae sp.1)、金龟科一种(Scarabaeidae sp.2);粪 金 龟 科1种 :粪 金 龟(Geotrupeslaeristriatus)。
研究区4月牛粪生境中的优势种为金龟科的三开蜣螂,个体数占牛粪生境的34.09%。5月优势种为蜉金龟科的黄缘蜉金龟,个体数占牛粪生境的22.21%。6月优势种为蜉金龟科的红亮蜉金龟,个体数占牛粪生境的21.67%。7月优势种为金龟科的北方蜣螂,个体数占牛粪生境的30.42%。8-10月优势种均为蜉金龟科的直蜉金龟,个体数分别占各自生境49.90%、93.55%、99.6%(图1)。
6月牛粪中捕获的食粪昆虫个体数在7个月中仅排第5位,但拥有最多的物种和丰富度(图2),Simp⁃son指数、Shannon-Wiener指数、均匀度指数也高于其他月份(表1)。8月牛粪中捕获食粪昆虫个体数最少,但各项指数不是最低的。10月牛粪中捕获的食粪昆虫个体数量最多(图1),但各项指数均最低。不同月份物种的丰富度与物种的数量呈正相关,物种的数量越多其测度值也越高。各月牛粪中食粪昆虫的物种数与各地的丰富度是相一致的:6月>5月>7月>4月>8月>9月>10月(表1)。在物种总数相同的条件下,多样性指数与各物种个体数量的平均程度呈正相关。物种的均匀度指数则受各物种个体数量分布的均匀性影响。因此,尽管10月有最大的个体数量,但因各物种的个体数量分布极不均匀,即直蜉金龟占了捕获个体数的绝大部分,致使其多样性和均匀度指数均最低。

表1 不同月份牛粪中食粪昆虫的多样性Table 1 The diversity of insects in cattle dung in different months
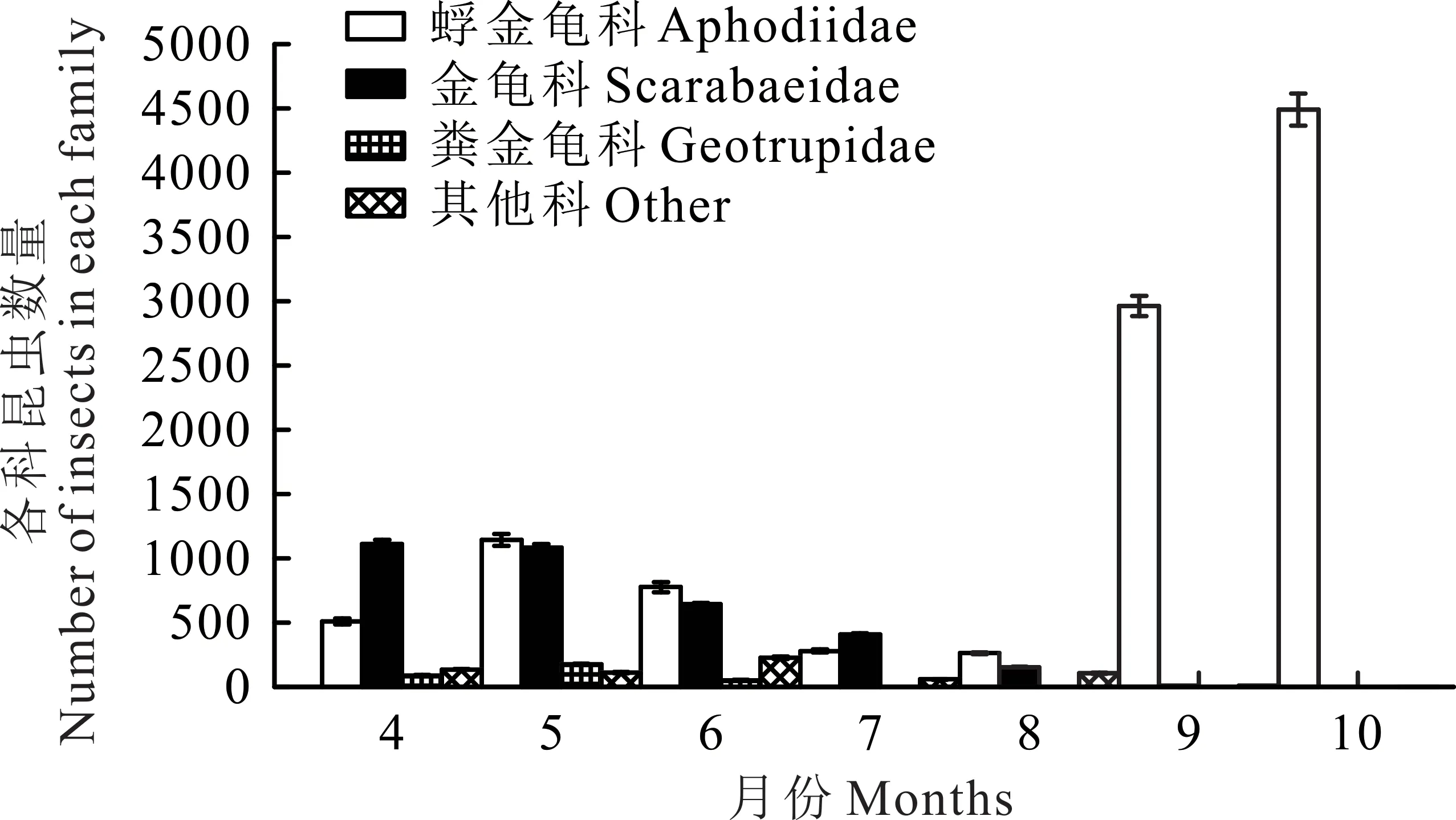
图1 各科昆虫不同月份数量变化Fig.1 Insects of different families in different months
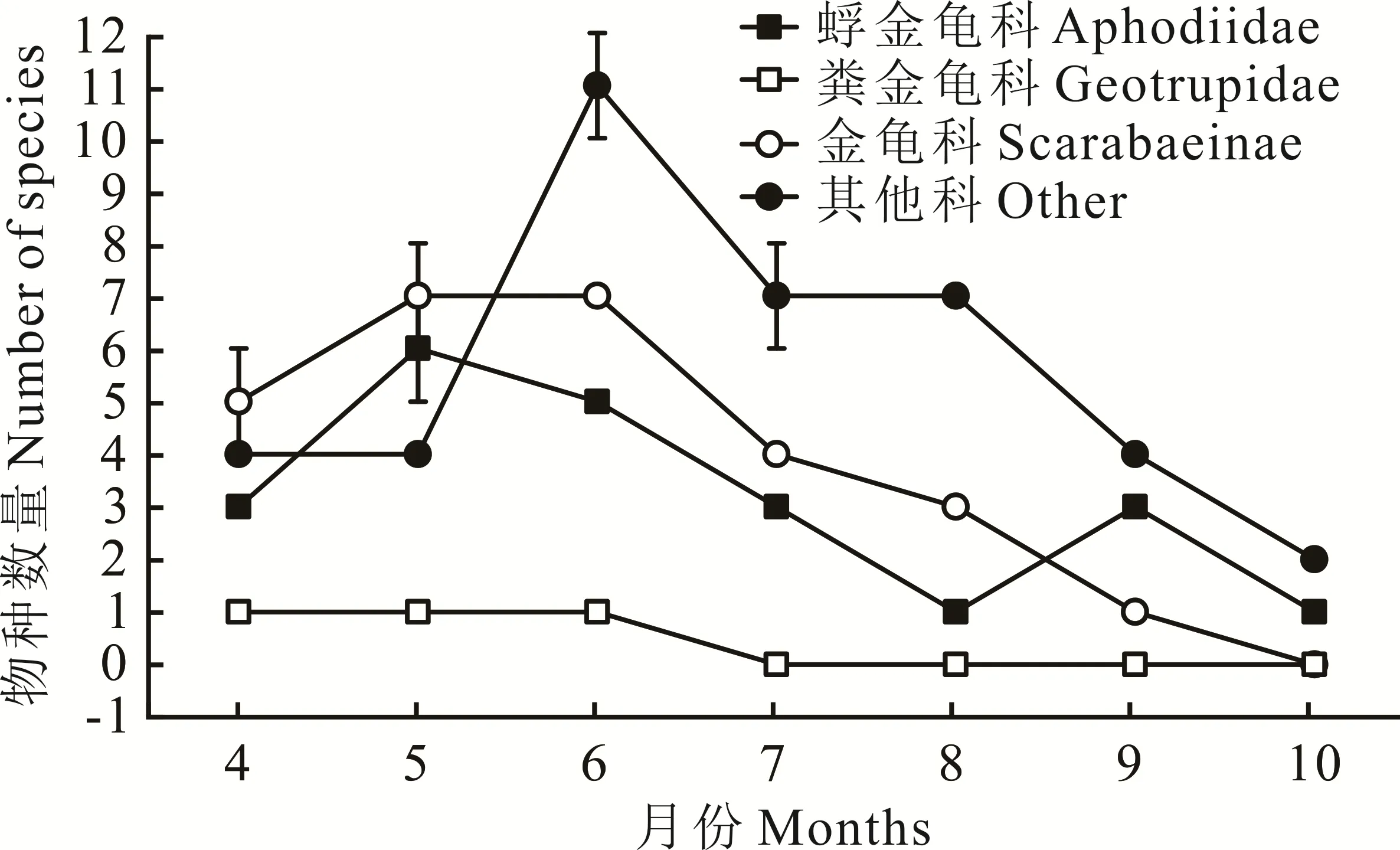
图2 各科昆虫不同月份种类动态变化Fig.2 The dynamic change of insects of differ ent families in different months
2.2 粪甲虫进驻粪便行为观察
行为观察试验在6月粪甲虫物种数最多和10月粪甲虫个体数最大的两个月进行。主要观察粪甲虫拜访粪便的方式,不同种类的粪甲虫到达粪便的先后顺序。不同时间段粪便中累计活动的粪甲虫数量以及被粪甲虫分解的粪便状态。6月粪甲虫种类多数量少,在粪便中活动时间主要集中在新鲜粪便放置的前24 h,数量高峰在粪便放置8 h左右(累计100+)。优势种为内生型粪甲虫红亮蜉金龟,其次是远生型粪甲虫粗纹弯边蜣螂。10月粪甲虫种类少数量多,在粪便中活动时间主要集中在新鲜粪便放置的前8 h,数量高峰在粪便放置4 h左右(累计2800+),粪便放置23 h粪甲虫完全结束在粪便中的活动。优势种为内生型粪甲虫直蜉金龟(表2)。
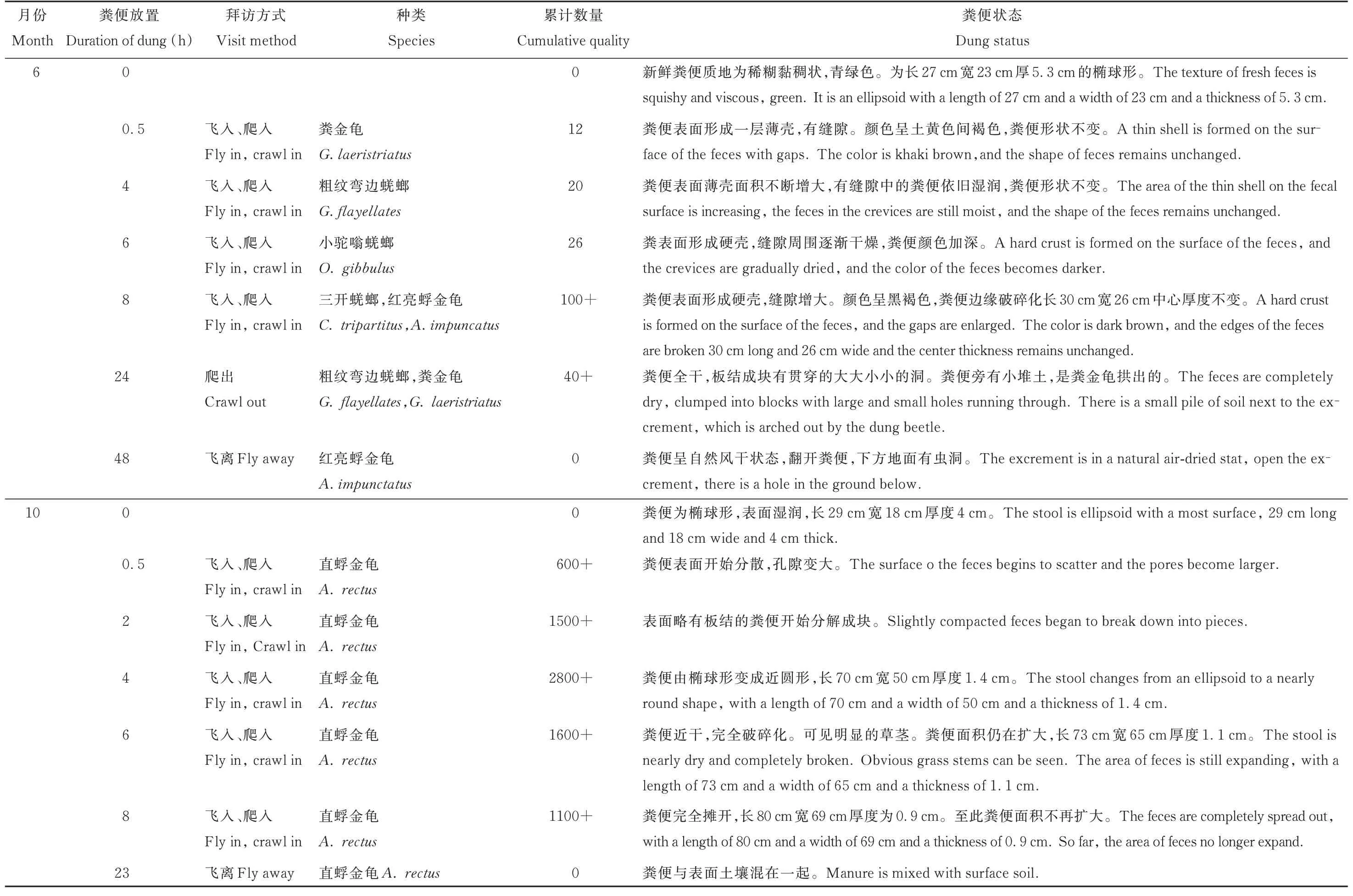
表2 粪甲虫进驻粪便行为观察Table 2 Observation on the behavior of dung beetle in d
6月的温度与粪甲虫数量没有显著相关性(表3);湿度与粪甲虫数量没有显著的相关性;风速与粪甲虫数量呈显著负相关(P<0.05);光照强度与粪甲虫数量呈显著正相关(P<0.05);10月温度与粪甲虫数量呈显著正相关(P<0.05);湿度与粪甲虫数量呈极显著负相关(P<0.01);风速与粪甲虫数量没有显著的相关性;光照强度与粪甲虫数量呈极显著正相关(P<0.01)。

表3 粪甲虫数量与环境因子之间的皮尔森相关系数Table 3 Pearson’s correlation coefficient between the number of dung beetles and environmental factors
2.3 粪甲虫进驻对粪便内种子的分配
80%的珠子被埋藏在水平距离20 cm以内及范围0~10 cm的土壤深度中(图3),主要处理珠子的粪甲虫为内生型粪甲虫、外生型粪甲虫和远生型粪甲虫。有20%的珠子被粪甲虫埋藏到10 cm以下或移动到水平范围20 cm以外,主要处理珠子的粪甲虫为远生型粪甲虫。3种尺寸的珠子在0~1 cm、2~5 cm、5~10 cm的埋藏深度中均表现出显著性差异(P<0.05)。在0~1 cm的埋藏深度中大、中、小3种尺寸的珠子埋藏率呈逐渐降低的趋势,且差异显著(P<0.05)。在2~5 cm的埋藏深度中小珠子的埋藏率最高,与大、中珠子差异显著(P<0.05)。在5~10 cm埋藏深度中,大珠子的埋藏率为0,与中、小珠子差异显著(P<0.05)。3种尺寸的珠子在>10 cm的埋藏深度中差异不显著(P>0.05)。
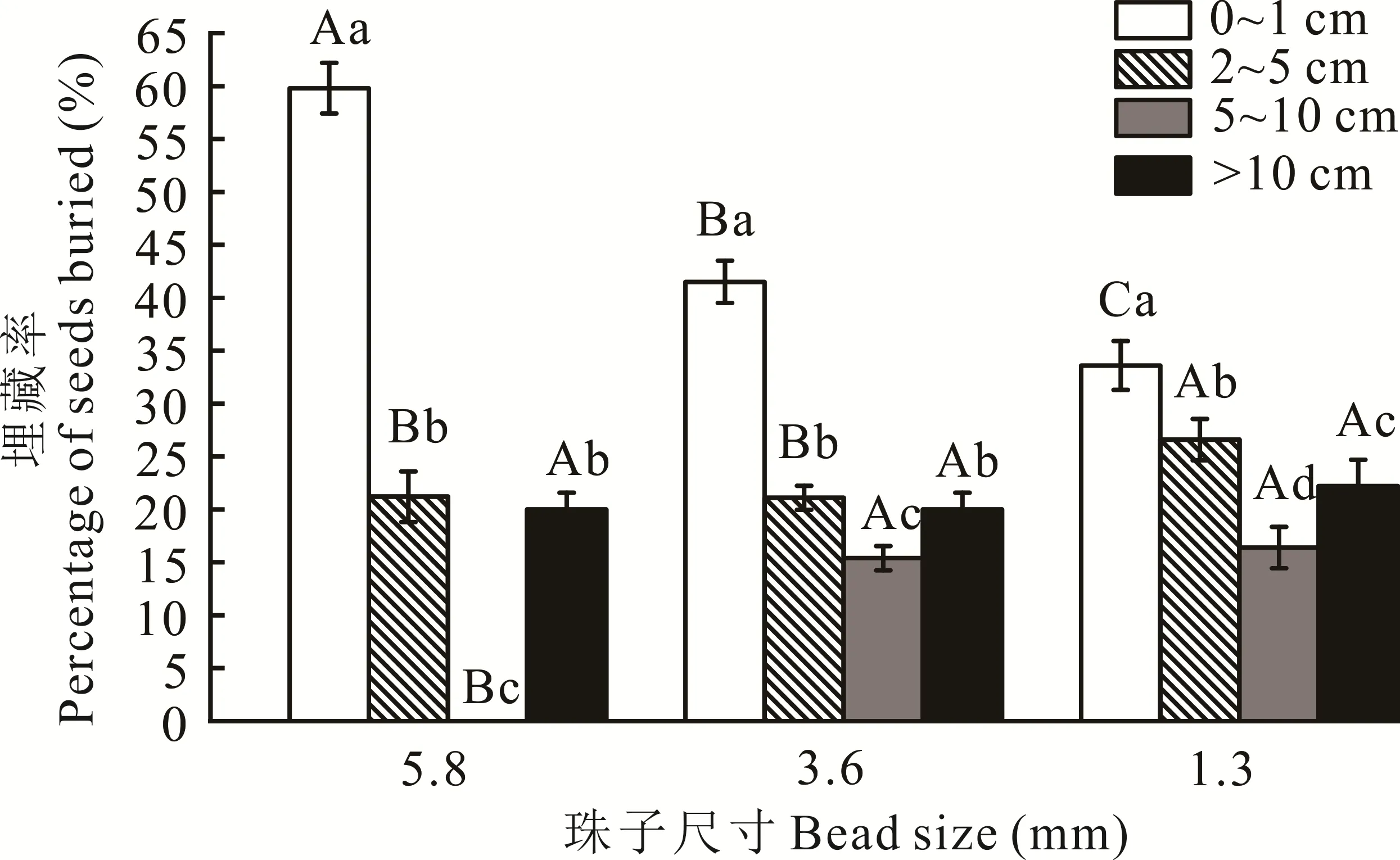
图3 粪甲虫对不同尺寸珠子的埋藏率Fig.3 Burial rate of dung beetle to different bead sizes
3 讨论
3.1 粪甲虫的多样性特征
粪甲虫多样性的研究主要集中在热带雨林和非洲稀树草原中,研究发现草原上粪甲虫的数量是森林的8倍,草原上远生型粪甲虫白天主导,外生型粪甲虫晚上主导。森林里外生型粪甲虫在白天和晚上都是主导,物种丰富度很低[17]。在印度的牧区研究中,内生型粪甲虫在春天至秋天活动频繁,远生型粪甲虫在春至夏季以及冬天之前活动频繁[18]。温带草原的研究发现低纬度地区远生型粪甲虫和外生型粪甲虫占主导地位,北部温带地区内生型粪甲虫占主导地位并有少量的外生型粪甲虫伴随,本研究结果与此一致。干燥地区的远生型粪甲虫数量比潮湿地区的多,可能是因为潮湿地区的土质湿黏不易滚动以及树叶凋落物过厚阻碍其活动[19]。
本试验在取样过程中除粪甲虫外还在粪便中采集到不少食粪性昆虫,在数据统计结果中会直接影响到牛粪中物种丰富度,但由于采集到的粪甲虫数量庞大,所以对其他指数影响较小。作为荒漠草地生态系统的主要成员,粪甲虫对生态系统的平衡有重要意义。通过本试验分析认为导致不同月份牛粪中粪甲虫群落多样性差异的主要因素有2个。一是气候因素决定了粪甲虫群落的主要组成,温度、湿度、风速和光照强度都对粪甲虫有影响。二是季节性放牧影响粪甲虫的种类和数量,研究地作为春秋牧场,承担了两季放牧压力。粪甲虫依靠嗅觉寻找粪便[20],粪便的数量和密度会直接影响粪甲虫种类和数量。
3.2 粪甲虫拜访行为的日变化及其与环境因子之间的关系
影响6和10月粪甲虫拜访行为日变化不同的主要原因是粪甲虫的种类和数量,6月的牛粪中活动的优势种为内生型粪甲虫红亮蜉金龟,其次是远生型粪甲虫粗纹弯边蜣螂。10月的优势种为内生型粪甲虫直蜉金龟。不同功能类群的粪甲虫对粪便处理方式的差异导致利用时长不一致。6月粪甲虫数量少竞争小,因而能在粪便中获得足够的食物再离开。但10月个体数量大粪便是稀缺资源,当粪便中的养分不能维持取食或产卵育幼时粪甲虫也会自行离开。其次环境因子也对粪甲虫的活动有重要影响,随着气温的升高或风速过大粪便失水干燥,开始结块,粪甲虫很难利用此类粪便也会离开。粪甲虫数量与环境因子之间的皮尔森相关系数有明确的季节性差异。6月物种丰富度高,温度和湿度对粪甲虫数量没有表现出显著相关性。10月的粪甲虫物种较为单一且数量大,环境因子对其影响较纯粹,粪甲虫表现出喜热喜干的特性。这与热带雨林研究的粪甲虫喜热喜湿情况不符,可能的原因是环境和气候的差异造成不同地区不同物种的聚集,而绢蒿荒漠炎热干燥,物种不断适应当地气候所致。
3.3 粪甲虫对种子的搬运和埋藏特征
随着种子大小的增加,埋入种子的比例降低[21]。模拟试验结束,在土壤表面仅可见少量珠子。不同功能类群的粪甲虫对粪便处理方式的差异导致不同尺寸珠子的埋藏率不同。本试验认为0~1 cm层的珠子大部分是由内生型粪甲虫加工的[22],3种尺寸的珠子在此深度埋藏率最高,与6月的优质种内生型粪甲虫红亮蜉金龟相对应。2~10 cm的珠子则由粪金龟加工,多数粪金龟会将珠子埋在2~5 cm粪便下方挖掘的洞中,少数粪金龟会因竞争将珠子深埋。遗失的珠子或被掩埋深度大于10 cm被认为是远生型粪甲虫粗纹弯边蜣螂移出了试验区。粗纹弯边蜣螂将粪便团成球状,滚到一定距离后掘洞并把粪球埋到洞中,作为食物球或孵化球,其中珠子会掺杂在粪球中被二次传播。
在主要通过哺乳动物排便传播种子的情况下,粪甲虫虽然不是专性主动的种子传播者,但通过利用哺乳动物粪便在多阶段种子扩散过程中起关键作用。粪甲虫对于微环境主动而严格的选择性使得粪便内种子的存活率大大增加,具有传播方式上的优越性。粪甲虫在将其用于喂养或产卵之前将其掩埋。在此过程中,嵌入粪便中的一粒或几粒种子可能会意外地埋在粪便下的隧道中或远离粪便的粪球中[23]。从粪甲虫的角度来看,粪便中存在的种子是污染物[24]。从种子的角度来看,离开母株和埋葬可能会极大地影响其生存[25-26]。埋在地下的种子比地面上残留的种子更有可能逃脱捕食[21]。此外,种子最终可能处于潮湿的环境中,更适合发芽。粪甲虫对种子的埋藏有利于建立土壤种子库,种子在土中休眠,随后成功发芽[27]。从植物的角度来看,深度很重要,因为当种子被更深地埋藏时,它们更容易被掠食者发现[28]。然而,粪甲虫深埋种子也会抑制出苗[29],即使深度小于5 cm的微小增加也可以显著降低出苗和成功建植的可能性[30]。
4 结论
1)绢蒿荒漠粪甲虫的物种多样性特征存在季节性差异,春季粪甲虫物种丰富度高但个体数量少。秋季粪甲虫物种丰富度低但个体数量大。
2)6月的粪甲虫对粪便的拜访行为数量高峰在8 h,累计100+,24 h后数量开始减少,活动持续48 h完全结束。10月的粪甲虫对粪便的拜访行为数量高峰在4 h,累计2800+,6 h后数量开始减少,活动持续23 h完全结束;粪甲虫的拜访数量与环境因子之间存在显著相关性。
3)不同尺寸的珠子之间的埋藏率差异显著,不同深度之间的埋藏率有显著性差异。随着种子大小的增加,埋入种子的比例逐渐降低。
