铜绿丽金龟线粒体全基因组及其系统发育分析
2023-03-07曲春娟朱悦江晨曲明静王向誉李晓
曲春娟 朱悦,2 江晨 曲明静 王向誉 李晓
(1. 山东省花生研究所,青岛 266100;2. 塔里木大学农学院,阿拉尔 843300;3. 山东省蚕业研究所,烟台 264000)
铜绿丽金龟Anomala corpulenta隶属于鞘翅目Coleoptera 多食亚目Polyphaga 金龟总科Scarabaeoidea金龟科Scarabaeidae 丽金龟亚科Rutelinae,在我国及日本、朝鲜、东南亚等国广泛分布,其幼虫危害玉米、花生、大豆、小麦等多种作物根系或荚果,造成直接或间接经济损失,成虫经常聚集暴食叶片,危害榆树、杨树、葡萄等多种林果木[1-2],对其全面深入的研究对农林生产具有重要意义。
丽金龟亚科昆虫全世界已知有235 属4 200 余种,其中有许多种类是重要的农林害虫[3]。目前丽金龟亚科的系统发育研究主要是纳入在金龟总科和金龟科的研究中,针对丽金龟的研究则相对缺乏。而且尚有诸多问题需要解决,比如丽金龟亚科的单系性有待明确、丽金龟外部形态趋同现象普遍导致的近缘种区分困难等[4]。随着分子生物学和测序技术的发展,基因序列被广泛应用到昆虫的系统发育学研究中。相比传统形态学手段,基因序列在解决趋同进化问题方面更有优势。尤其是线粒体基因因其母系遗传、进化速率快等特点,在这一领域被广泛应用[5-6]。对于金龟总科的分子系统学研究,过去主要借助于线粒体单基因、单个核基因或者少数2-3 个基因的联合,例如cox1、cox2、16S rRNA、12S rRNA、cytb、nad1等线粒体基因都是经常用于进化分析的分子靶标[7]。然而单个基因不仅所包含的进化信息较少,难以完全解析物种间的系统发育关系,在选择压力和进化速率方面也存在差异,容易导致“长枝吸引”等问题,而完整的线粒体基因组能够克服上述弊端,成为研究后生动物进化历史和亲缘关系更加强有力的工具[8-9]。以前传统的线粒体基因组测序主要依赖于通用引物和长片段PCR扩增,效率低下,费事费力;尤其是控制区高AT含量、连续重复碱基较多的特点,导致Sanger 测序获取全长序列比较困难。随着高通量测序技术的发展,该策略被逐步应用到线粒体基因组测序中来,成为获取线粒体基因组序列的更有力手段[10]。
目前已完成线粒体全基因组测序的丽金龟只有日本弧丽金龟Popillia japonica[11]、棉花弧丽金龟Popillia mutans[12]和墨绿彩丽金龟Mimela splendens(MZ064554)。此外还有非全长序列JX412777(Popilliasp.)和JX412788(Adoretussp.)。其中仅弧丽金龟属Popillia两个种的线粒体基因组是基于高通量测序组装获得。而异丽金龟属Anomala的线粒体基因组却未见报道。该属是丽金龟亚科中种类丰富度最高的属,已记录的超过1 000 种[4]。其中的代表性物种之一——铜绿丽金龟是我国华北农田三大优势金龟甲之一[13-14],受到昆虫学家及植保工作者密切关注。针对其开展线粒体全基因组的研究,不仅可以丰富金龟科尤其是丽金龟亚科昆虫的线粒体基因组信息,而且能够为铜绿丽金龟的分子系统学、群体遗传学及分子生态学研究提供重要的基础信息。
本研究利用高通量测序技术Illumina 平台对铜绿丽金龟进行线粒体基因组测序,完成组装和注释后,对其线粒体基因组的结构和基因排列、碱基组成、密码子使用情况,以及转运RNA(transfer RNA,tRNA)的结构等进行预测分析,结合已发表的金龟科线粒体基因组序列对金龟科各亚科、属和种之间的系统发育关系进行探讨,旨在为丽金龟的系统发育研究和线粒体基因组学研究提供方法参考和依据。
1 材料与方法
1.1 材料
供试铜绿丽金龟成虫于2020年7月采自山东省花生研究所莱西望城试验基地,将活体试虫放入无水乙醇中于-20℃冰箱保存备用。选择试虫胸部和足的肌肉组织提取基因组总DNA,所用提取方法为改进的CTAB 法,采用1%琼脂糖凝胶电泳和核酸分析仪NanoDrop 2000 分别测定提取DNA 的质量和浓度。
1.2 方法
1.2.1 Illumina HiSeq 高通量测序 DNA 样品送至武汉百奥维凡生物科技有限公司构建350 bp 的小片段测序文库和进行高通量测序。基于边合成边测序(Sequencing By Synthesis, SBS)技术和Illumina HiSeq X 测序平台对所构建的测序文库进行双端150 bp 测序,利用NGS QC Toolkit 2.3.3[15]将原始测序数据进行过滤,去除adapter 序列、低质量末端、含N>10%的reads 以及长度小于25 bp 的短片段后,得到11.29 Gb 的clean reads。
1.2.2 序列拼装、注释及特征分析 利用SPAdes v3.11.1(http://cab.spbu.ru/software/spades/)[16]拼 接软件对clean reads 进行拼接,构建contigs。使用SSPACE 软件[17]对contigs 进行扩展延伸,获得最终的完整线粒体基因组序列。利用MITOS 在线服务器(http://mitos.Bioinf.uni-leipzig.de)[18]对线粒体基因组序列进行功能注释。并进一步通过与已知金龟科物种的线粒体基因进行同源比对注释结果进行核对验证。利用tRNAscan-SE 软件(http:∥lowelab.ucsc.edu/tRNAscan-SE/)[19]对tRNA 基因进行查找及二级结构的预测。利用Mega 11[20]分别计算铜绿丽金龟线粒体基因组中各编码基因的碱基组成、密码子使用频率、AT-skew 和GC-skew。
1.2.3 系统发育分析 为研究铜绿丽金龟在金龟科的系统发育地位,利用线粒体基因组的13 个蛋白质编码基因(protein-coding genes,PCGs)的核苷酸序列进行系统发育分析。选取已报道的19 种金龟科昆虫的线粒体基因组作为参考序列,以牙甲科昆虫Sphaeridium bipustulatum为外群,采用最大似然法(maximum likelihood,ML)和贝叶斯法(Bayesian,BI)构建系统发育树。利用Clustal X 2.0[21]对核苷酸序列进行多重比对后,使用Gblocks v0.91b[22]过滤比对结果,然后采用SequenceMatrix v1.7[23]对每个基因的比对结果进行串联连接。基于SMS 软件[24]和ModelFinder[25]对建树数据集评估得到的最适替代模型为GTR+I+G。以PhyML3.0 在线分析软件[26]进行1 000 次bootstrap 运算,构建ML 系统发育树。利用MrBayes 3 软件[27]计算200 000 代,每运算100 代取样保存,舍弃25%的老化样本,构建BI 系统发育树。最后使用FigTree v.1.4.3(http://tree.bio.ed.ac.uk/software/figtree)对系统发育树进行绘制。
2 结果
2.1 铜绿丽金龟mtDNA基因组结构组成和分布特征
铜绿丽金龟线粒体基因组全长16 673 bp(GenBank 登录号:OL449520),呈现典型双链闭合环状结构,包括13 个PCGs、2 个rRNA、22 个tRNA 共37 个基因以及1 074 bp 的AT 富集区,与后生动物线粒体基因组的经典基因组成一致。昆虫线粒体基因组的多数基因在同一条链上编码,该链称为J 链(majority strand);其余少数基因在另一条链上编码,该链称为N 链(minority strand)。对铜绿丽金龟线粒体而言,23 个基因位于J 链上,包括9 个PCGs(nad3、cox3、atp6、atp8、cox2、cox1、nad2、cytb和nad6)和14 个tRNA 基因;14 个基因位于N链,包括4 个PCGs(nad5、nad4、nad4l和nad1)、8 个tRNA 基因和2 个rRNA 基因(图1)。与其他昆虫线粒体一样,铜绿丽金龟线粒体基因组也存在基因重叠和间隔现象,表现为30 处长度1-41 bp 的碱基重叠现象和5 处长度1-15 bp 的碱基间隔现象(表1)。基因排布与已报道的其他丽金龟:墨绿彩丽金龟、日本弧丽金龟和棉花弧丽金龟一致,与果蝇代表的祖先模式也一致。

表1 铜绿丽金龟线粒体基因位置与起始终止密码子Table 1 Locations and start/stop codons of mitochondrial genes in A. corpulenta

图1 铜绿丽金龟线粒体基因组结构图Fig. 1 Mitochondrial genome structure of A. corpulenta
2.2 线粒体基因组碱基组成
总的来看,铜绿丽金龟线粒体基因组中A+T 含量占比75.87%,G+C 含量占比24.13%,表现出明显的AT 偏向性。其中PCGs、tRNA 基因和rRNA 基因的A+T 含量分别为74.87%、76.54%和75.79%。全基因组的AT-skew 为正值(0.027),GC-skew 为负值(-0.237),表明整个基因组更偏好于使用A 碱基和C 碱基。整个线粒体基因组不同位置对碱基偏好性具有一定差异。22 个tRNA 基因和2 个rRNA 的AT-skew 均为正值,表明其偏好A 碱基;13 个PCGs和2 个rRNA 的AT-skew 均为负值,表明其偏好T碱基;从不同的单基因来看,除atp8外其余12 个PCGs 的AT-skew 外均为负值,J 链上的9 个PCGs的GC-skew 均为负值,N 链上的4 个PCGs 和rRNA基因的GC-skew 均为正值,与大多数昆虫的线粒体基因组AT/GC 偏斜一致(表2)。

表2 铜绿丽金龟线粒体基因组碱基组成Table 2 Base composition in the mitochondrial genome of A. corpulenta
2.3 线粒体基因组蛋白质编码基因及密码子使用情况
铜绿丽金龟线粒体基因组含有13 个PCGs,序列全长11 212 bp(占比67.25%),除去终止密码子(37 bp)共编码3 725 个氨基酸残基。使用最频繁的密码子为UUU(Phe)、UUA(Leu)、AUU(Ile)和UAU(Tyr),而CGC(Arg)、CGG(Arg)、CCG(Pro)和GCG(Ala)使用相对较少(表3),由此也反映了核苷酸组成的AT 偏好性。
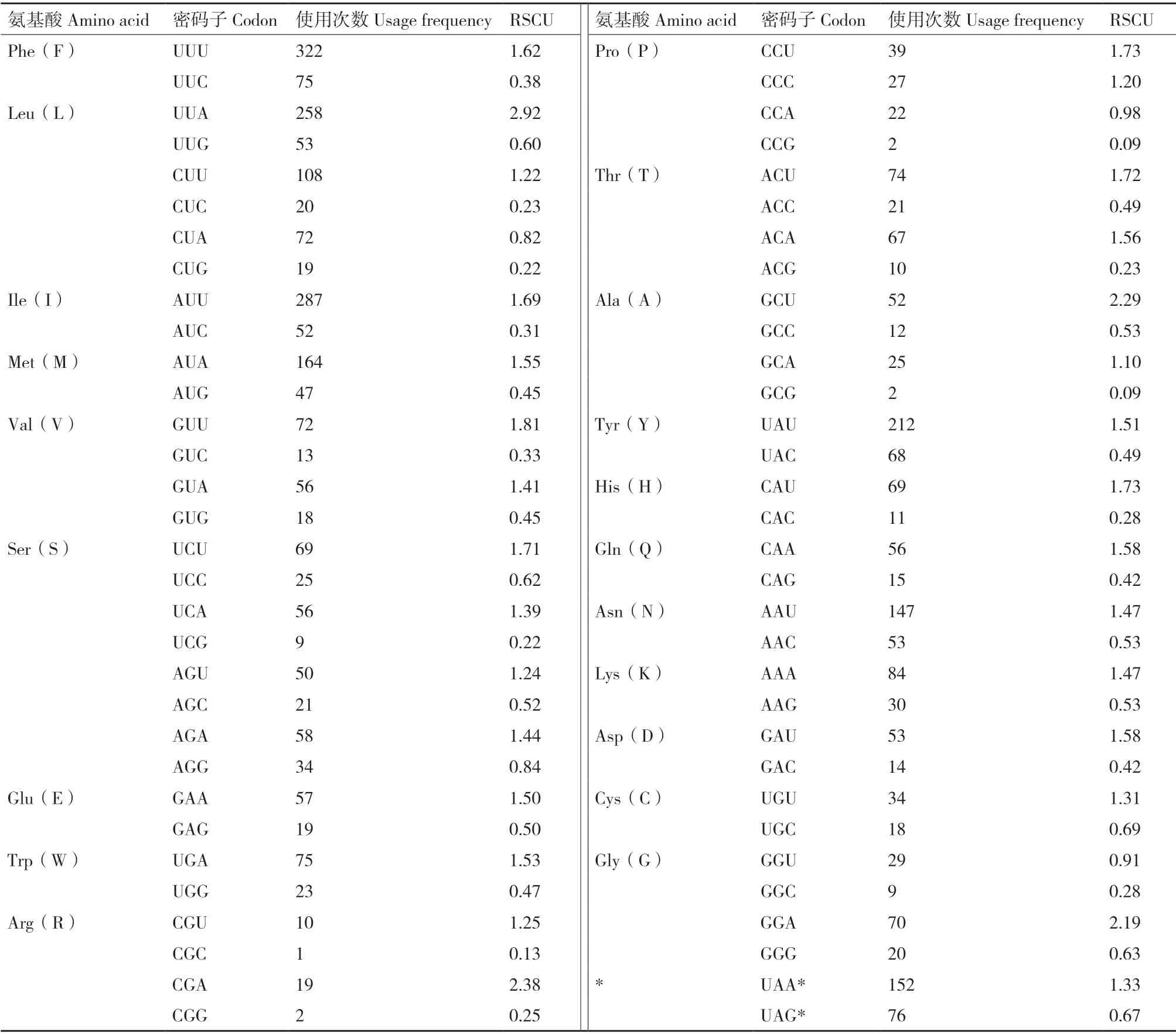
表3 铜绿丽金龟线粒体基因组相对同义密码子使用频率(RSCU)Table 3 Relative synonymous codon usage(RSCU)in the mitochondrial genome of A. corpulenta
所有PCGs中nad5最长(1 717 bp),atp8最短(157 bp)。除nad4l以TTG 为起始密码子外,其余PCGs都以ATN 作为起始密码子,其中nad2、cox3、nad4和cytb以ATG 为起始密码子,atp6和nad1以ATA为起始密码子,cox1、atp8和nad5以ATT 为起始密码子,cox2、nad3和nad6以ATC 为起始密码子。cox3和nad4以不完整的TA 为终止密码子,缺失的核苷酸由转录后3 苷端多聚腺苷酸化补齐[28],其余PCGs 均以TAA 或TAG 为终止密码子,与昆虫线粒体基因组普遍使用的终止密码子一致。
2.4 线粒体基因组tRNA和rRNA
铜绿丽金龟的22 个tRNA 基因长度范围为63 bp(tRNACys)到72 bp(tRNALys),总长度为1 450 bp。预测的二级结构如图2 所示,除tRNASer(AGN)缺失DHU 臂外,其余tRNA 序列都可以折叠成典型的三叶草式二级结构。此外,tRNASer和tRNALeu以双拷贝形式存在,其余tRNA 基因均仅显示单拷贝。铜绿丽金龟的两个rRNA 基因均在N 链上,16S rRNA位 于tRNALeu(CUN)和tRNAVal之 间,长 度1 350 bp;12S rRNA在tRNAVal和控制区之间,长度800 bp。

图2 铜绿丽金龟线粒体基因组tRNA 二级结构Fig. 2 Putative secondary structures of tRNAs in the mitochondrial genome of A. corpulenta
2.5 系统发育分析
以 牙 甲 科(Sphaeridiinae) 中 的Sphaeridium bipustulatum为外群,使用已报道的丽金龟亚科(Rutelinae)、金龟亚科(Scarabaeinae)、花金龟亚科(Cetoniinae)、鳃金龟亚科(Melolonthinae)、犀金龟亚科(Dynastinae)、蜉金龟亚科(Aphodiinae)共19 种金龟科昆虫及本研究测得的铜绿丽金龟线粒体基因组的13 个PCGs 的核苷酸序列,构建金龟科昆虫的ML 和BI 系统发育树。结果显示两种建树方法得到的拓扑结构完全一致,金龟亚科和蜉金龟亚科聚为1 个分支,丽金龟、花金龟、犀金龟和鳃金龟亚科聚为1 个分支,置信值均为100%。除鳃金龟亚科外,各亚科均聚为一支且置信值较高。但与传统分类不同,鳃金龟亚科的5 个种并未聚为一个支系。各亚科之间的系统发生关系为(蜉金龟亚科+金龟亚科)+(鳃金龟亚科(花金龟亚科(犀金龟亚科+丽金龟亚科)))(图3)。
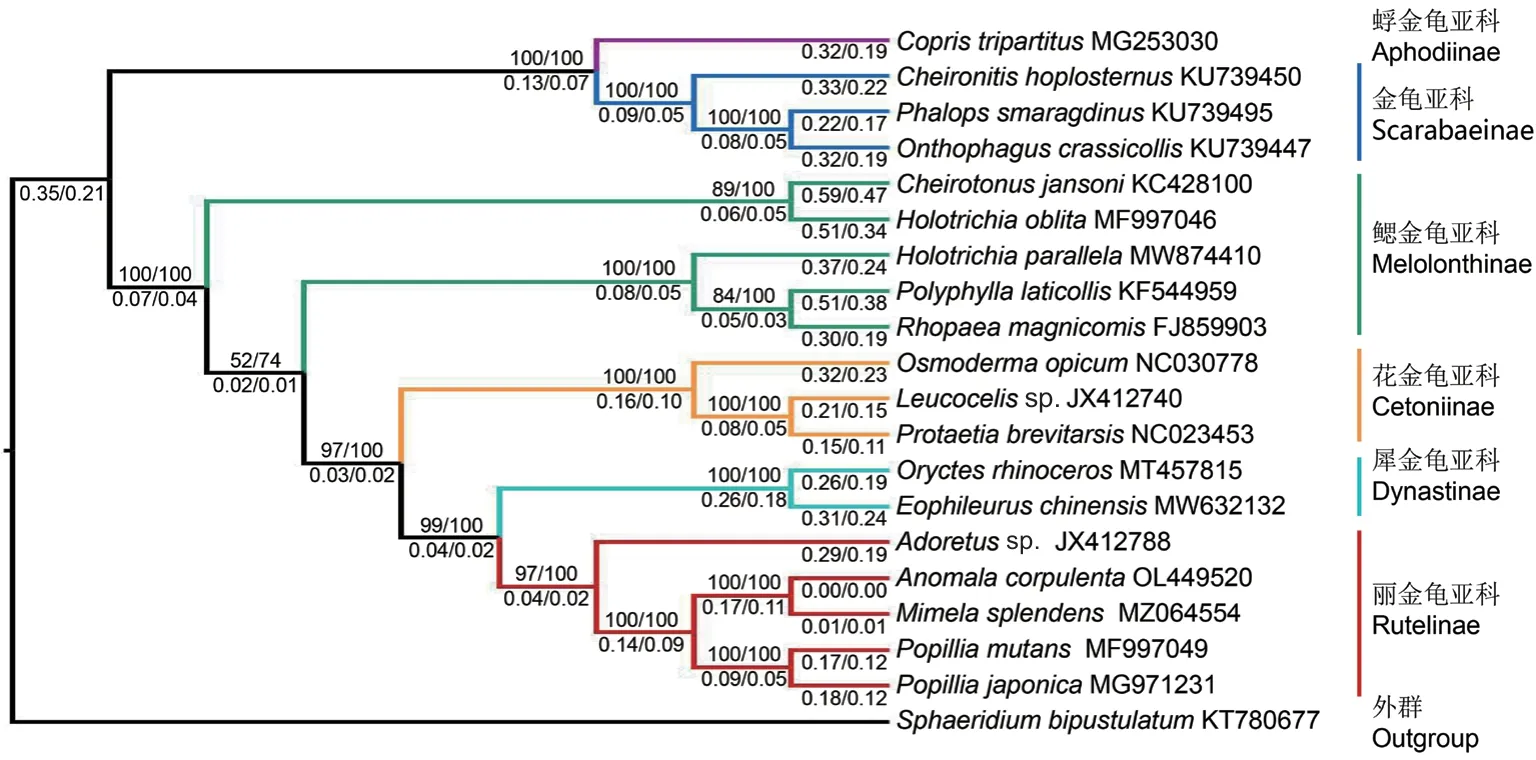
图3 基于线粒体蛋白质编码基因核苷酸序列构建的铜绿丽金龟与其他金龟科昆虫的系统发育树(最大似然法和贝叶斯法)Fig. 3 Phylogenetic tree of A. corpulenta and other insects species of Scarabaeidae based on the mitochondrial protein-coding gene sequences(maximum likelihood and Bayesian)
铜绿丽金龟聚在丽金龟亚科分支系,与墨绿彩丽金龟组成姐妹关系,且置信值均为100%。利用Kimura-2-Parameter 参数模型计算丽金龟亚科5 个种之间的遗传距离,结果所反映的亲缘关系与系统发育树一致:铜绿丽金龟与墨绿彩丽金龟的遗传距离为0.007 9,而与棉花弧丽金龟和日本弧丽金龟的遗传距离分别为0.172 7 和0.177 4,与喙丽金龟属Adoretussp.的遗传距离则为0.209 9(表4)。
3 讨论
线粒体基因组具有母系遗传、结构简单、进化速率快的特点,作为分子标记有其独特优势,目前被广泛应用到遗传学、系统发育学、生物地理学以及物种诊断学等研究领域[5-6]。随着测序技术的进步,越来越多的昆虫线粒体基因组完成测序[5]。这些昆虫线粒体基因组与其他后生动物线粒体一样,呈现共同特点:为共价闭合环状双链DNA 分子,长度为14-20 kb,通常含有37 个基因,包括13 个PCGs、22 个tRNA 基因、2 个核糖体RNA(ribosomal RNA,sRNA)基因及一个或数个AT 富集区[29-30]。
本研究首次测定和分析了铜绿丽金龟的完整线粒体基因组,丰富了金龟科的线粒体基因组信息,尤其是为研究异丽金龟Anomala属在丽亚科中的系统发育位置提供了数据基础。铜绿丽金龟线粒体基因组全长16 673 bp,与NCBI 已公布的其他3 种丽金龟(墨绿彩丽金龟、日本弧丽金龟和棉花弧丽金龟)的线粒体基因组大小近似(15 148-16 541 bp)[11-12]。包含37 个线粒体基因,排布方式与上述3 种丽金龟完全一致,而且与果蝇Drosophila yakuba也一致[31]。D. yakuba线粒体的基因排布方式代表了昆虫线粒体基因排布的祖先模式[31],说明铜绿丽金龟线粒体未发生基因重排现象。
在昆虫纲中,线粒体基因组的蛋白质编码基因已被广泛证实可以有效解决不同分类阶元的系统发育问题[5]。本研究以线粒体基因组的13 个蛋白质编码基因作为分子靶标,探索了金龟科6 个亚科(丽金龟亚科、金龟亚科、花金龟亚科、鳃金龟亚科、犀金龟亚科、蜉金龟亚科)的分类学关系[3]。在已公布线粒体基因组序列的几种丽金龟物种中,日本弧丽金龟、棉花弧丽金龟和墨绿彩丽金龟具有完整的线粒体基因组序列。Adoretussp.(JX412788)的线粒体基因组虽然是部分序列,但包含了完整的13 个PCGs 序列信息,因此也被纳入到建树数据集。但Phylloperthasp.(KX087335)和Popilliasp.(JX412777)的线粒体基因组由于缺失部分PCGs的数据,未被采用。

Table 4 Pairwise genetic distances of mitochondrial protein-coding gene sequences between A. corpulenta and other phytophagous species of Scarabaeidae based on Kimura-2-Parameters
已知的金龟科昆虫全世界超过30 000 种,按照食性分为粪食性和植食性两大类[32-33]。其中粪食类包括金龟亚科和蜉金龟亚科,植食类包括犀金龟、花金龟、丽金龟和鳃金龟。本研究构建的系统发育树也清晰得显示了金龟科是由这两个主要群体构成的。植食性金龟子各亚科之间的系统发生关系为鳃金龟亚科+(花金龟亚科+(犀金龟亚科+丽金龟亚科)),与Ayivi 等[33]和Song 等[12]基于线粒体基因组ML 树和BI 树的分析结果一致。丽金龟亚科和犀金龟亚科互为姐妹群,也与目前多数学者基于形态学和分子证据得出的主流观点一致[4]。此外,丽金龟、犀金龟、花金龟和金龟亚科的单系性均得到很高的支持,说明系统发育关系可信度较高。同时我们发现鳃金龟亚科并非单系群,这与之前报道一致[12,33-35]。
在属级阶元关系上,系统发育树的结果显示丽金龟亚科一支中,异丽金龟属Anomala与彩丽金龟属Mimela的亲缘关系更近,弧丽金龟属Popillia次之,而喙丽金龟属Adoretus较远,且在该分支上两种分析方法的置信值均为100%。遗传距离分析的结果同样证明了上述关系,支持了Bouchard 等[3]关于丽金龟亚科的分类系统。但齿爪鳃金龟属Holotrichia两个种(暗黑鳃金龟H. parallela和大黑鳃金龟H.oblita)并没有聚到一起,还需要进一步研究。
本研究对铜绿丽金龟线粒体基因组全序列的测定,丰富了金龟科线粒体基因组的序列信息以及结构和组成信息,为后续丽金龟科系统发育关系研究奠定基础,同时也为铜绿丽金龟的群体遗传学和分子生态学研究提供了基础信息。
4 结论
获得了铜绿丽金龟线粒体全基因组,其基因的排布方式与墨绿彩丽金龟、日本弧丽金龟和棉花弧丽金龟相同,且与祖先模式一致,系统发育分析支持丽金龟亚科的单系性,异丽金龟属Anomala与彩丽金龟属Mimela的亲缘关系较其与弧丽金龟Popillia和喙丽金龟属Adoretus更近。