濒危植物香果树种子萌发特性研究
2021-08-19郭连金肖志鹏殷崇敏吴原榕胡金平刘艳艳钟友春
郭连金, 肖志鹏, 殷崇敏, 吴原榕, 胡金平, 刘艳艳, 钟友春
濒危植物香果树种子萌发特性研究
郭连金1,*, 肖志鹏1,2, 殷崇敏1, 吴原榕1, 胡金平1, 刘艳艳1, 钟友春1
1. 上饶师范学院生命科学学院, 上饶 334001 2. 中南林业科技大学风景园林学院, 长沙 410004
论文研究了濒危植物香果树()种子萌发特性。通过室内萌发实验初步检测了香果树种子的休眠, 采用靛蓝检验法检测了生活力, 通过室内室外条件的贮藏检验了种子寿命, 通过人工气候箱研究了萌发对温度、光强条件(9—24 ℃, 500—9000 lx交互)的响应。结果表明: 香果树果实成熟后, 种子无明显的休眠, 有生活力种子75.3%±7.9%, 无生活力10.9%±9.2%, 空粒6.5%±1.8%, 涩粒7.4%±6.4%; 贮藏四个月, 室内外贮藏种子萌发率高于78.6%±6.5%, 贮藏八个月, 室外贮藏种子全部失去活力, 贮藏一年, 室内贮藏种子萌发率20.2%±8.3%。无光照时香果树种子不萌发; 在500—9000 lx处理下, 12—24 ℃香果树种子最终萌发率均高于90%, 9 ℃下均低于80%。9 ℃时, 光强显著影响最终萌发率, 500 lx下为33.3%±6.9%, 3000 lx下为70.3%±4.2%。温度升高, 香果树种子平均萌发周期缩短, 萌发指数及活力指数上升。在12℃、15 ℃下的2000 lx、3000 lx下平均萌发周期显著低其他光强处理下, 萌发指数显著高于其他光强处理。总体上, 12 ℃、15 ℃时, 光强增强, 活力指数在500—3000 lx上升, 在3000—9000 lx间下降。在温度较高的18 ℃、21 ℃、24 ℃下, 平均萌发周期, 萌发指数, 活力指数受光强的影响明显弱于12 ℃、15 ℃下。野外香果树种子萌发期间, 野外环境温度在12—18 ℃之间,光照强度对香果树种子萌发有重要影响。合适的光照环境(1000—3000 lx)可能是野外香果树种子萌发、幼苗建植的关键。
濒危植物; 香果树; 种子萌发; 种子特性; 环境因子
0 前言
种子萌发受多种因素的影响。濒危植物毛柄小勾儿茶(var.)和秦岭冷杉()种子生活力差导致其濒危[1–2]。望天树()及闽楠()种子寿命短是其濒危的重要原因[3–4]。此外, 环境因子对一些濒危植物种子萌发有重要影响。濒危植物合柱金莲木()及濒危红树植物红榄李()因种子萌发所需温度范围十分狭窄致其濒危[5–6]。
香果树为我国特有的单属种植物, 为第四纪冰川期孓遗种, 种群衰退, 濒临灭绝, 现为林业部公布的国家珍贵树种、国家Ⅱ级重点保护植物[7–10]。目前, 对香果树种子特性已有初步研究, 有研究认为香果树种子在5 ℃低温下保存较室温下的种子寿命更长[11], 20 ℃恒温有利于香果树种子萌发[12], 1000 lx光强有利香果树种子萌发[13]。然而, 对香果树的种子休眠, 自然(室外)条件下种子寿命, 种子生活力, 以及高于1000 lx的光强条件下香果树种子的萌发及香果树种子萌发温度范围尚无相关报道。此外, 光照对一些植物种子萌发的影响随温度变化而改变[14–16], 目前香果树种子萌发此方面的尚无相关研究。
因此, 为进一步探究香果树种子萌发特性, 探讨在萌发阶段是否存在致危因素, 本文对香果树种子休眠, 生活力、种子寿命、香果树种子对环境因子的响应等特性进行研究, 以期探究如下问题: (1)香果树种子生活力如何, 是否存在休眠现象, 自然贮藏条件下, 种子寿命如何。(2)香果树种子萌发对温度、光照的响应, 以及香果树种子萌发过程中光照与温度的交互作用。
1 材料和方法
1.1 实验仪器
对人工气候箱(杭州钱江仪器设备有限公司, 型号GZP—450)进行加光改造, 在培养箱一侧安装LED灯条(5730型), 黑色卡纸局部遮光以生成不同光强环境。照度计(TES—1332A)实测培养箱内每个处理位置的光强。
武夷山香果树分布地林下温度测定采用嘉兴米速电子有限公司提供的 WH—2081型手持气象站。
1.2 供试材料
供试种子2015年11月采自福建省武夷山国家级自然保护区(27°33′—27°54′ N, 117°27′—117°51′ E), 将采集的种子混合均匀, 以干燥牛皮纸袋室温保存备用。
1.3 实验方法
种子生活力:2016年4月, 随机取100粒混合均匀的种子, 重复4次。采用靛蓝检验法[17],
种子休眠:于2015年11月利用人工气候箱进行萌发实验, 观察香果树种子是否萌发, 以露白为标志, 种子萌发则表明香果树无明显休眠实验环境条件为20 ℃恒温, 光强1000 lx, 每日照光12 h。实验处理下3次重复, 每次重复100粒种子, 种子取自混合均匀的种子。
种子寿命:2015年11月开始实验, 设置室内贮藏及室外贮藏两种贮藏方式, 室外贮藏将种子置于干燥不积水的林内空地, 种子以无纺布包裹避免动物取食或种子散落, 2015年12月, 2016年4月, 2016年8月, 2016年12月分别取回种子, 并在20 ℃, 1000 lx人工气候箱中进行种子萌发实验观察种子最终萌发率, 每个处理50粒种子, 重复3次。于肉眼下人工筛选大而饱满的香果树种子作为供试种子。
种子萌发:于2016年4月上旬进行实验。利用加光改造后的人工气候箱, 设置六个恒温条件: 9 ℃、12 ℃、15 ℃、18 ℃、21 ℃、24 ℃, 在其中每个温度条件下各设置八个光强条件9000 lx、7000 lx、5000 lx、3000 lx、2000 lx、1000 lx、500 lx及黑暗条件。照光处理下每日定时照光12 h。于肉眼下人工筛选大而饱满的香果树种子作为供试种子, 置于装有加垫双层滤纸的培养皿内, 每日定时观察, 并更换培养皿中的蒸馏水, 使种子始终保持湿润状态。每个处理3个重复, 每个重复30粒种子。实验开始后每日记录种子萌发情况, 萌发以露白为标志。胚根长度以电子游标卡尺(精度0.01mm)进行测量, 每种处理下三次重复。自种子停止萌发5 d后结束实验。
1.4 数据处理与统计分析方法
(1)最终萌发率(Final germination):=萌发种子数量/试验用种子数量100%
(2)平均萌发周期(Mean period of final germination):()
(3)萌发指数(Germination index):()
(4)活力指数(Vigor index):()
为第天发芽数,为发芽的天数,为幼苗的胚根长。
采用方差分析(ANOVA)和LSD多重检验(< 0.05)方法对不同处理下的种子萌发参数进行分析, origin 9.0进行作图, SPSS 12.0进行显著性分析(SPSS Inc. Chicago, Illinois, USA)。
2 结果与分析
2.1 种子特性检测
生活力测定结果: 有生活力香果树种子占香果树种子总数为75.3%±7.9%, 无生活力种子占10.9%± 9.2%, 空粒种子占6.5%±1.8%, 涩粒种子占7.4%±6.4%。
休眠实验表明, 香果树果实成熟后(外果皮呈褐色), 其种子在20 ℃恒温,每日12小时1000 lx光强条件下即可萌发, 最终萌发率为80.3%±8.1%, 即香果树的果实成熟后, 种子不具明显的休眠现象。
图1为香果树种子在两种贮藏方式(室内及室外)下随贮藏时间增加的最终萌发率, 即香果树种子的寿命。由图1可知, 实验开始时(未贮藏处理时)香果树种子萌发率达到93.2%±5.2%及96.4%±3.2%, 贮藏四个月后, 香果树种子萌发率下降, 但两种贮藏方式下最终萌发率无显著差异, 室内、室外分别为78.6%±6.5%, 86.2%±12.1%; 贮藏八个月时, 处理间差异显著, 室外贮藏种子全部失去活力, 室内贮藏种子33.4%±5.4%的种子萌发, 贮藏一年时, 室内贮藏种子萌发率为20.2%±8.3%。
2.2 种子萌发实验
2.2.1 最终萌发率
无光照时香果树种子在所有实验温度下均不能萌发。
图2—A为不同温度、光强条件共同作用下香果树种子的最终萌发率。由图知, 9 ℃处理下最终萌发率均低于80%, 12—24 ℃处理下, 最终萌发率均高于90%。9 ℃时, 光照强度显著影响最终萌发率(<0.05), 最终萌发率随光强在500—3000 lx的升高而迅速升高, 3000—9000 lx间随光强升高而缓慢升高, 500 lx下为33.3%±6.9%, 3000 lx为70.3%±4.2%, 9000 lx下为75.6%±6.9%。
2.2.2 平均萌发周期
图2—B为不同温度、光强条件共同作用下香果树种子的平均萌发周期。随温度升高, 平均萌发周期缩短, 总体上, 光强增加, 平均萌发周期缩短。9 ℃、12 ℃、15 ℃下, 香果树种子平均萌发周期在2000 lx及3000 lx下平均萌发周期缩短明显。12℃和15 ℃下的2000 lx、3000 lx光强处理下, 香果树种子平均萌发周期显著同温度其他光强处理(<0.05)。
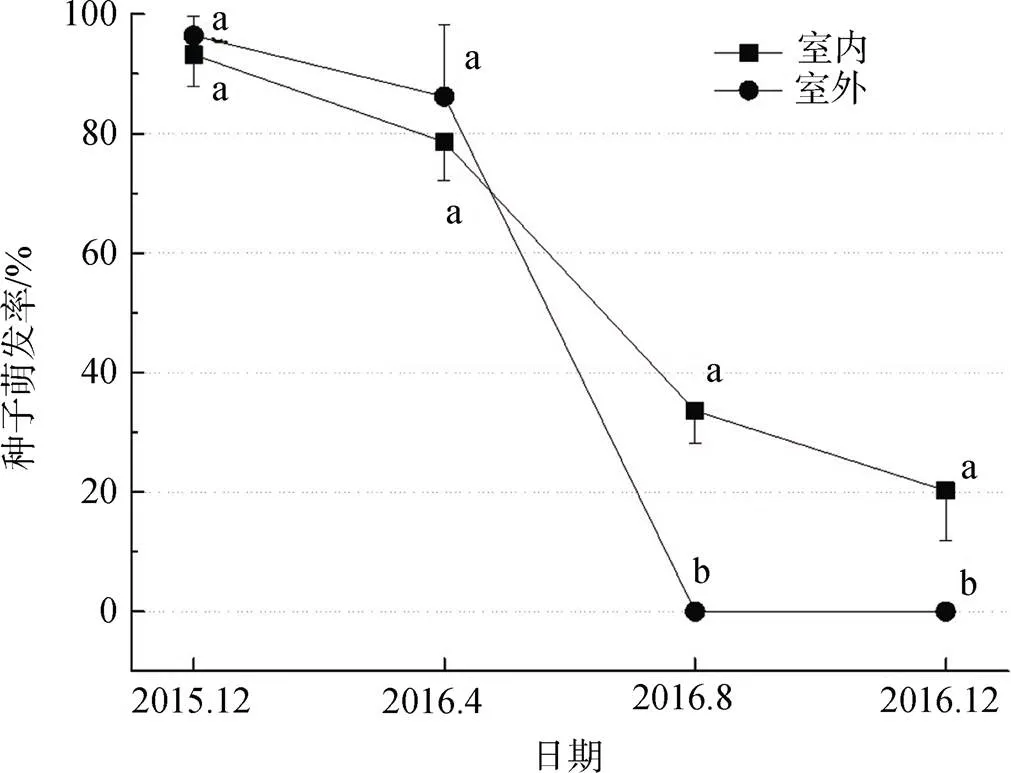
注: 同一时间香果树种子萌发率标注有不同字母, 表示萌发率之间在0.05水平差异显著。
Figure 1 The effect of the storage method on the seed longevity of
所有处理下的平均萌发周期中, 9 ℃和500 lx下最长, 为(18.3±0.05) d, 24 ℃和9000 lx下最短, 为(5.71± 0.42) d; 12 ℃的各光强处理下, 2000 lx下最短, 为(10.3± 0.03) d, 500 lx下最长, 为(14.45±0.05) d; 15 ℃的各光强处理下, 3000 lx下最短, 为(8±0.07) d, 500 lx下最长, 为(11.1±0.05) d。18 ℃的各光强处理下, 2000 lx下最短, 为(7.11±0.03) d, 500 lx下最长, 为(8.68±0.05) d。
2.2.3 萌发指数
图2—C为不同温度、光强条件共同作用下香果树种子的萌发指数。由图知, 9 ℃—24 ℃间, 随温度升高, 香果树种子萌发指数升高。总体上光强增强, 萌发指数上升, 9 ℃—18℃间, 萌发指数在2000 lx及3000 lx处理下上升明显; 温度12℃、15℃和光照2000 lx、3000 lx条件下, 香果树种子萌发指数显著高于同温度其他光强处理(<0.05)。
2.2.4 活力指数
图2—D为不同温度、光强条件共同作用下香果树种子萌发幼苗的活力指数。由图知, 9 ℃时活力指数接近于0(实验中观测到9 ℃时香果树种子仅露白)。12—24 ℃间随温度升高, 活力指数上升。总体上, 12 ℃、15 ℃时, 活力指数随光强在500 lx—3000 lx间升高而上升, 在3000 lx—9000 lx间随光强升高而下降。12 ℃、15 ℃下同一光强(在2000 lx、3000 lx、5000 lx下)对活力指数的影响明显, 较18 ℃、21 ℃、24下有明显不同。

注: 同一温度下香果树种子萌发率标注有不同字母, 表示萌发率之间在0.05水平差异显著; 2—A.香果树种子最终萌发率; 2—B.香果树种子平均萌发周期; 2—C.香果树种子萌发指数; 2—D.香果树种子活力指数。
Figure 2 Response of seed germination ofto light and temperature

图3 2016年3—4月武夷山香果树分布地林下温度
Figure 3 Environmental temperature of the distribution area ofin Wuyi mountain from march to April 2016
3 讨论
种子休眠有利于种子避免在不合适的时间地点萌发[18], 本研究表明, 香果树种子不具有明显的休眠特性。野外条件下香果树种子在4月大量发现萌发[19–20], 但室外贮藏四个月后(即野外香果树种子萌发时间), 室内室外香果树种子萌发率无显著差异, 并达到78.6%±6.5%以上, 说明香果树种子无明显休眠及种子寿命不是限制香果树萌发的原因。秦岭冷杉种子生活力仅为25%左右, 致其濒危[2], 香果树果实成熟后, 其种子生活力达75.3%±7.9%, 生活力不是其濒危的原因。9 ℃情况下, 香果树种子最终萌发率可达到70 %以上, 9 ℃—24 ℃间香果树种子最终萌发率均大于90%(图3)。香果树种子萌发温度范围较大, 最终萌发率高, 表明萌发温度范围狭窄不是其种群更新的主要限制因素。
本研究中, 3000 lx下平均萌发周期与500 lx下相比, 在12 ℃、15 ℃分别可提前4.06 d、3.10 d(图2—B)。3000 lx下的萌发指数与500 lx下的萌发指数的比值, 在12 ℃、15 ℃, 分别为1.68、1.37(图2—C)。3000 lx下活力指数与500 lx下活力指数的比值在12 ℃、15 ℃分别为7.27、1.82(图2—D)。12 ℃、15 ℃下光强对香果树种子萌发有重要影响。野外调查中发现, 香果树种子在其飘落次年四月开始萌发[19–20]。图3为2016年3—4月武夷山香果树分布地林下温度, 由图知, 四月时环境温度在基本稳定在12—18℃间(图3)。
根据室内萌发实验的结果, 此时光强对香果树种子的萌发有较大影响, 处于适宜光强条件下的香果树种子能够快速萌发并快速完成幼苗建立, 香果树种子小, 种子贮藏的营养物质少, 合适的光照环境(1000 lx—3000 lx)可能是野外香果树种种子萌发并建植的关键。对香果树土壤种子库的研究表明, 香果树种子多数散落于枯落物中[21]。这导致多数香果树种子缺少合适的光照环境, 可能是香果树种群衰退的一个原因。
[1] 党海山, 张燕君, 江明喜, 等. 濒危植物毛柄小勾儿茶种子休眠与萌发生理的初步研究[J]. 武汉植物学研究, 2005, 23(4): 327–331.
[2] 赖江山, 李庆梅, 谢宗强. 濒危植物秦岭冷杉种子萌发特性的研究[J]. 植物生态学报, 2003, 27(5): 661–666.
[3] 严理, 云朝光, 秦武明, 等. 珍稀濒危树种望天树的研究进展[J]. 林业科技, 2014, 39(2): 59–62.
[4] 吴大荣, 王伯荪. 濒危树种闽楠种子和幼苗生态学研究(英文)[J]. 生态学报, 2001, 21(11): 1751–1760.
[5] 柴胜丰, 蒋运生, 韦霄, 等. 濒危植物合柱金莲木种子萌发特性[J]. 生态学杂志, 2010, 29(2): 233–237.
[6] 杨勇, 钟才荣, 李燕华, 等. 濒危红树植物红榄李种子形态及萌发特性[J]. 分子植物育种, 2016, 14(10): 2851–2858.
[7] 傅立国, 金鉴明. 中国植物红皮书—稀有濒危植物(第一册)[M]. 北京: 科学出版社, 1992.
[8] 于永福. 中国野生植物保护工作的里程碑—《国家重点保护野生植物名录(第一批)》出台[J].植物杂志, 1999, 5(3): 3–11.
[9] 郭连金. 濒危植物香果树()种群结构与动态[J]. 武汉植物学研究, 2009, 27(5): 509–514.
[10] 陈子林, 康华靖, 刘鹏, 等. 大盘山自然保护区香果树群落结构特征[J]. 云南植物研究, 2007, 29(4): 461–466.
[11] 万军, 曹小军, 张小平, 等. 香果树种子贮藏试验[J]. 四川林业科技, 2016, 37(5): 33–35.
[12] 甘聃, 陈发菊, 梁宏伟, 等, 徐伟. 珍稀濒危植物香果树种子萌发特性研究[J]. 种子, 2006, 25(5): 27–29.
[13] 管康林. 香果树种子的光萌发特性初步研究[J]. 浙江林学院学报, 1985, 2(2): 47–50.
[14] 杨期和, 宋松泉, 叶万辉, 等. 种子感光的机理及影响种子感光性的因素[J]. 植物学通报, 2003, 20(2): 238–247.
[15]GALLAGHER RS, CARDINA J.Phytochrome-mediatedgerminationⅠ: effect of seed burial and germination temperature[J].Weed Science, 1998, 46(1), 48–52.
[16] 赵笃乐. 光对种子休眠与萌发的影响(下)[J]. 生物学通报, 1995, 30(8): 27–28.
[17] 唐润琴, 李先琨, 欧祖兰, 等. 濒危植物元宝山冷杉结实特性与种子繁殖力初探[J]. 植物研究, 2001, 21(3): 403–408.
[18] 付婷婷, 程红焱, 宋松泉. 种子休眠的研究进展[J]. 植物学报, 2009, 44(5): 629–641.
[19] 郭连金, 徐卫红 , 房会普, 等. 不同年龄香果树种子雨和种子库及其更新特征[J]. 西北植物学报, 2016, 36(11): 2273–2282.
[20] 郭连金, 曹昊玮, 徐卫红, 等. 香果树()种群种子雨、种子库及实生苗数量的海拔梯度变化[J]. 植物研究, 2017, 37(3): 377–386.
[21] Guo L J, Xue P P, Li M, et al. Seed bank and regeneration dynamics of, population on the western side of Wuyi Mountain, south China[J]. Journal of Forestry Research, 2017(5): 1–10.
The research on characteristics of seed germination of endangered plant
GUO Lianjin1,*, XIAO Zhipeng1,2, YIN Chongmin1, WU Yuanrong1, Hu Jinping1, LIU Yanyan1, ZHONG Youhun1
1.College of Life Science, Shangrao Normal University, Shangrao 334001, China 2. College of Landscape Architecture, Central South University of Forestry & Technology, Changsha 410004, China
This work aimed to investigate the seed germination characteristics of the endangered plant. The seed dormancy ofwas preliminarily characterized via a germination experiment in the laboratory. Seed viability was determined through the indigo test. Seed longevity during storage under indoor and outdoor conditions was tested, and the response of seed germination to temperature and light intensity (9 ℃-24 ℃, 500-9000 lx interaction) was studied by applying an artificial climate box. Results showed that the seeds ofdid not undergo significant dormancy after fruit ripening. Viable, nonliving, empty, and abortive seeds accounted for 75.3% ± 7.9%, 10.9% ± 9.2%, 6.5% ± 1.8%, and 7.4% ± 6.4% of the total number of seeds. After 4 months of storage, the seed germination rate was higher than 78.6% ± 6.5% regardless of whether the seeds were stored indoors or outdoors. After 8 months of storage, all the seeds stored under outdoor conditions lost their vitality. The germination rate under indoor storage for 1 year was 20.2% ± 8.3%. The seeds ofdid not germinate in the absence of light. Under 500-9000 lx treatment, the final seed germination rate ofat 12 ℃ –24 ℃ was higher than 90% and that at 9 ℃ was less than 80%. At 9 ℃, light intensity significantly affected the final germination rate, which was 33.3% ± 6.9% under 500 lx and 70.3% ± 4.2% under 3000 lx. With the increase in temperature, the mean period of final germination was shortened, and seed germination and vigor indexes increased. At 12 ℃ and 15 ℃, the mean period of final germination was significantly reduced under 2000 and 3000 lx. However, the germination index was significantly reduced under other light intensities. In general, at 12 ℃ and 15 ℃ and as the light intensity was increased, the vigor index increased when the light intensity was within the range of 500-3000 lx and decreased when the light intensity was within the range of 3000-9000 lx. The average final germination period, germination index, and vigor index at 18 ℃, 21 ℃, and 24 ℃ were significantly lower than those at 12 ℃ and 15 ℃. Light intensity had an important influence on the seed germination ofwhen the ambient temperature was between 12 ℃ and 18 ℃. A suitable light environment (1000-3000 lx) may be the key to seed germination and seedling establishment ofin the wild.
endangered plant,seed germination, seed characteristics, environmental factors
郭连金, 肖志鹏, 殷崇敏, 等. 濒危植物香果树种子萌发特性研究[J]. 生态科学, 2021, 40(4): 13–18.
GUO Lianjin, XIAO Zhipeng, YIN Chongmin, et al. The research on characteristics of seed germination of endangered plant[J]. Ecological Science, 2021, 40(4): 13–18.
10.14108/j.cnki.1008-8873.2021.04.002
Q948.11
A
1008-8873(2021)04-013-06
2020-01-03;
2020-03-02
国家自然科学基金项目(32060336); 国家自然科学基金项目(31860200); 国家级大学生创新训练计划项目(201610416003)
郭连金(1976—), 男, 山东滨州人,硕士,教授, 主要从事植物生态学和生物多样性研究, E-mail: guolianjin@163.com
郭连金
