胡椒PnPAL基因家族全基因组鉴定及表达模式分析
2021-08-18苏岳峰丁元昊郝朝运胡丽松郑其向
苏岳峰,丁元昊,郝朝运,胡丽松,郑其向,3,范 睿
(1.海南大学,海南 海口 570208;2.中国热带农业科学院香料饮料研究所,海南 万宁 571533;3.云南农业大学热带作物学院,云南 普洱 665000)
0 引言
【研究意义】胡椒(Piper nigrumL.)被称为“香料之王”,是胡椒科胡椒属植物,主产我国海南、云南和广东等地区。胡椒瘟病是由辣椒疫霉菌侵染胡椒的根系、叶片、花序、果穗、果实等部位而发病,具有气候依赖性且传染力极强的特点[1];病情严重时可导致全园毁灭,严重阻碍胡椒产业发展,缺乏抗瘟病种质是影响我国胡椒产业发展的主要问题[2]。因此,筛选胡椒关键基因进行功能验证对植物材料的遗传改良尤为重要。【前人研究进展】次生代谢是植物生长发育以及适应环境的重要环节。苯丙烷途径是响应多种逆境的关键途径,如:紫外线损伤、病原侵染、机械损伤、低温胁迫等。苯丙氨酸解氨酶是苯丙烷途径的关键酶和入口酶[3]。玉米的苯丙氨酸解氨酶(Phenylalanine ammonialyase,PAL)抵抗甘蔗花叶病毒,与木质素和水杨酸合成有关,陆地棉和马铃薯等中也有PAL 抵抗病菌侵染的报道[3-4]。此外苯丙氨酸解氨酶还是合成花青素和类黄酮等重要的胁迫响应次生代谢物质的必要酶[5]。目前,PAL的生物信息学分析已在水稻、棉花、桑树等作物中开展,分析结果显示PAL基因响应生物和非生物胁迫反应[6-8]。PAL在单双子叶植物分化之前已完成分化,通常是一个多基因家族,含有5 个左右家族成员[9]。PAL家族在不同作物中保守域不同,主要包括:苯丙氨酸保守结构域、PLN02457 和PLN02457 super family、PAL-HAL、phe-am-lyase 和Lyase aromatic 保守结构域、裂解酶I 类超家族保守结构域等[10-11]。然而,各物种所含保守域不同可能行使不同的生理功能,当归中PAL 催化类似的β 消除反应,与具有裂解酶I 类超家族保守结构域有关[12]。在柱花草中,PAL-HAL 是保守的脱氨位点[11]。目前,胡椒瘟病防控多以预防为主,防控成本极高。在分子生物学方面对胡椒瘟病的研究也仅限于栽培种与野生近缘种的比较转录组学研究和抗瘟病基因挖掘[13]。项目组前期的转录学结果显示胡椒抗/感种质(热引1 号、黄花胡椒)差异基因多富集于苯丙烷代谢途径,且组织化学显示苯丙氨酸解氨酶活性在抗性种质中较高[13]。本研究首次开展胡椒PnPAL基因研究。【本研究切入点】苯丙氨酸解氨酶是苯丙烷途径的入口酶及限速酶,在植物与病原菌互作过程中具有重要意义。因此,研究胡椒PnPAL的家族特征、进化关系及表达模式,能够为进一步确定胡椒PnPAL基因在抗瘟病中的功能奠定基础。【拟解决的关键问题】采用在线分析工具及TB tools 工具,胡椒PnPAL的顺式作用元件、motif、外显子和内含子位置、保守结构域,亲缘关系及接种病原菌下PnPAL基因的表达模式,有利于了解胡椒PnPAL的基因功能,便于胡椒新品种选育。
1 材料与方法
1.1 试验材料
我国主栽胡椒品种热引1 号(Piper nigrumL.)全基因组数据来自美国生物信息中心https://www.ncbi.nlm.nih.gov/bioproject/PRJNA529758。
1.2 试验方法
1.2.1PnPAL基因家族成员鉴定 根据胡椒基因组注释文件信息,鉴定到胡椒PnPAL基因家族成员和胡椒PnPAL的蛋白序列及染色体定位信息,再利用在线预测蛋白信息工具Expasy(https://www.expasy.org/)分析各个PnPAL的蛋白质分子质量、等电点和氨基酸个数,运用BUSCA 在线分析工具预测各PnPAL亚细胞定位(http://busca.biocomp.unibo.it/59a16590-35 06-4818-8017-7032b057b649/showresult/)。
1.2.2PnPAL基因结构及进化分析 根据基因组注释信息中的外显子和内含子长度分布信息,结合基因结构显示在线分析工具(GSDS)绘制胡椒PnPAL外显子、内含子位置示意图。利用Mega 5.0(http://gsds.cbi.pku.edu.cn/)的邻接法构建胡椒各PnPAL的亲缘关系。
1.2.3 胡椒PnPAL保守基序分析 运用motif 基础上的序列分析工具(MEME)预测蛋白序列保守基序(http://meme-suite.org/)和美国生物信息技术中心的CDD 工具分析所得domain和TB tools 绘制成图(https://www.ncbi.nlm.nih.gov/cdd)。
1.2.4 胡椒PnPAL启动子分析 利用在线数据库Plant CARE 和TB tools 分析胡椒PnPAL起始密码子上游2 000 bp 的顺式作用元件(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)。
1.2.5 蛋白系统进化分析 运用美国生物信息中心的Blast 工具,获得注释信息为苯丙氨酸解氨酶的蛋白序列,并结合拟南芥(Arabidopsis thaliana)、烟草(Nicotiana tabacumL.)、小麦(Triticum aestivumL.)、玉米(Zea mays)、水稻(Oryza sativa)、高粱(Sorghum bicolor)、大麦(Hordeum vulgareL.)、菠萝(Ananas comosus)、香蕉(Musa nana Lour.)、橡胶(Hevea brasiliensis)、棉花(Gossypium arboreum)、油菜(Brassica napusL.)、鹅掌楸(Liriodendron tulipifera)、牛樟(Cinnamomum micranthumf.kanehirae)、鳄梨(Persea americana)、睡莲(Nymphaea colorata)、无油樟(Amborella trichopoda)等作物的蛋白序列,采用Mega5.0 的邻接法分析物种间苯丙氨酸解氨酶的亲缘关系。数据来源于美国生物信息中心NCBI(https://www.ncbi.nlm.nih.gov/),拟南芥信息网站(https://www.arabidopsis.org/index.jsp),国家水稻数据中心(http://www.ricedata.cn/)。
1.2.6 针刺法接种病原菌 在扦插苗靠近土层的地上部用无菌注射器刺3 个小洞,呈三角形,用打孔器取一块直径为1 cm 的菌丝快,将有菌丝的一面覆在刺破的洞上,用浸湿的棉花缠绕固定菌块,起到保湿和固定的作用。
1.2.7PnPAL表达模式分析 使用Prime primer 5.0设计引物,引物见表1。针刺法在胡椒茎基部接种辣椒疫霉菌(参照1.2.6),以接种病原菌后0、8、12、24、48 h 的胡椒根系为材料,运用Real Time PCR 的方法分析7 条含有完整保守序列的PnPAL表达模式。

表1 实时荧光定量PCR 分析所用引物Table 1 Primers used in RT-PCR analysis
2 结果与分析
2.1 胡椒PnPAL 全基因组鉴定
如表2 所示,根据基因组注释信息,共鉴定到14 条胡椒苯丙氨酸解氨酶PnPAL基因,不均匀分布在9 条染色体上,其中有4 条位于2 号染色体上,3 条位于6 号染色体上,3 号、5 号、8 号、9 号、18 号、28 号和38 号染色体上各含有1 条。理化性质分析表明,PnPAL所含氨基酸残基数目在72~736,其中PnPAL14 含有的氨基酸残基数目最多,PnPAL8含有的氨基酸残基数目最少,所编码氨基酸残基数量均值为500 个。各个PAL 蛋白质分子量在7.374 41~83.431 07 kDa,蛋白质平均分子量为50.933 kDa。胡椒PnPAL编码的蛋白质等电点在5.76~9.77,平均等电点为7.18,其中PnPAL1、2、3、4、10、13、14 为酸性蛋白(理论等电点pI<7),PnPAL5、6、7、8、9、11、12 为碱性蛋白(理论等电点pI>7)。亚细胞分析表明,PnPAL1、2、3、4、10 位于细胞质中,PnPAL5、7、8、9 位于细胞核中,PnPAL6、11 位于叶绿体中,PnPAL12 位于内膜系统中,PnPAL13、14 位于质膜中。

表2 胡椒PnPAL 基因家族成员的理化性质Table 2 Physiochemical properties of PnPAL family in black peppers
2.2 胡椒PnPAL 基因结构及进化分析
如图1 所示,运用mega 对鉴定到的14 条PnPAL聚类分析表明,PnPAL4、10、3、13、14、1、2、5、7 聚为一类,PnPAL11、12 聚为一类,PnPAL6、8 聚为一类,表明胡椒PnPAL家族可分为3 个进化单元。运用GSDS 分析基因结构结果表明,家族成员外显子个数在1~5 个,内含子个数在1~4 个,其中PnPAL2、7、8、12、13 没有内含子,PnPAL3、4、5、10、14 含有两个外显子,PnPAL1 含有3 个外显子,PnPAL6 含有4 个外显子,PnPAL11 含有5 个外显子,平均外显子个数约为2 个。
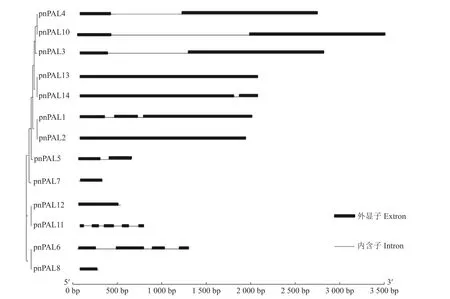
图1 胡椒PnPAL 基因家族基因结构分析Fig.1 Structure of PnPAL family in black peppers
2.3 胡椒PnPAL 保守基序分析
如图2 所示,在线分析工具MEME 鉴定到8 个保守基序,命名为Motif1~Motif8。在14 条胡椒PnPAL中,每条家族成员所包含motif 0~8 个不等,其中7 条PnPAL所含motif 个数最多,含有完整的8 个motif,3 条不含有motif,1 条含有5 个motif,1 条含有3 个motif。

图2 胡椒PnPAL motif 分析Fig.2 PnPAL motifs of black peppers
如图3 所示,NCBI CDD 结果表明,7 条含有PLN02457 结合位点,4 条PnPAL属于PLN02457 超家族,1 条含有保守结构域NEMP,1 条属于FNRlike 超家族,1 条属于bHLH-MYC-N 超家族。

图3 胡椒PnPAL domain 分析Fig.3 PnPAL domain of black peppers
综上,7 条含有完整motif 的家族成员均含有PLN02457 结合位点,推测此7 条家族成员为胡椒PnPAL家族成员。
2.4 胡椒PnPAL 启动子分析
如图4 所示,胡椒PnPAL基因家族成员除基本的顺势作用元件之外,还包括多个与激素和胁迫有关的顺式作用元件,包括低温响应元件、防御和应激相关元件、光反应元件以及水杨酸、茉莉酸甲酯响应元件等,其中含有脱落酸响应元件的PnPAL最多,达13 条,含有茉莉酸甲酯响应元件的PnPAL次之,有11 条,8 条含有生长素响应元件,5 条含有赤霉素响应元件,5 条含有防御和胁迫相关的元件,4 条含有水杨酸响应元件,4 条含有低温响应元件,1 条含有机械损伤响应元件,表明PnPAL参与生物和非生物胁迫响应,在植物防御和激素信号转导方面至关重要。PnPAL1所含顺式作用元件最多,达36 个;PnPAL8所含顺式作用元件最少,仅为17 个,平均顺式作用元件个数约为23 个,表明PnPAL响应多项逆境,其中PnPAL1含有启动子元件明显多于其他家族成员,PnPAL1可能响应多项逆境。

图4 胡椒PnPAL 启动子分析Fig.4 PnPAL promoter of black peppers
2.5 蛋白系统进化分析
苯丙烷途径是在裸子植物向被子植物进化过程中形成的,如图5 所示,本研究将具有完整结构的7个胡椒PnPAL基因与拟南芥(Arabidopsis thaliana)、烟草(Nicotiana tabacumL.)、小麦(Triticum aestivumL.)、玉米(Zea mays)、水稻(Oryza sativa)、高粱(Sorghum bicolor)、大麦(Hordeum vulgareL.)、菠萝(Ananas comosus)、香蕉(Musa nana Lour.)、橡胶(Hevea brasiliensis)、棉花(Gossypium arboreum)、油菜(Brassica napusL.)、鹅掌楸(Liriodendron tulipifera)、牛樟(Cinnamomum micranthumf.kanehirae)、鳄梨(Persea americana)、睡莲(Nymphaea colorata)、无油樟(Amborella trichopoda)等植物的PAL基因进行进化分析,这些基因主要分为单子叶与双子叶两类。然而,胡椒的PnPAL基因均处于双子叶植物的位置,且与木兰类的植物亲缘关系较为接近。此外,PnPAL1,PnPAL2,PnPAL13,PnPAL14 位于系统发生树的基部,表明这4 个基因属于较原始的类型;另一方面,PnPAL3,PnPAL4,PnPAL10 位于离系统发生树基部较近的位置,且与木兰类的植物聚在一起。现代植物学家普遍承认“真花学说”,认为木兰类的植物属于较原始的类群,因此,胡椒的PnPAL基因较为古老,与胡椒的系统地位相似[14-15]。

图5 胡椒PnPAL 系统发育树
2.6 胡椒PnPAL 表达模式分析
对筛选到完整的7 条PnPAL进行表达模式分析,结果表明PnPAL3、PnPAL10 有表达,其余5 条未检测到信号。其中PnPAL3 和PnPAL10 在接种后0、8、12、24、48 h 的根系中均有表达。如图6 所示,PnPAL3 在接种病原菌初期表达相对较高,这可能与植物的应激反应有关。如图7 所示,PnPAL10表达在处理组中基本呈上调趋势,特别是在接种病原菌后的24、48 h 显著高于对照组基因的相对表达,表明该基因可能在病原菌侵染胡椒根系过程中起主要作用。

图6 胡椒PnPAL3 基因相对表达分析Fig.6 Relative expressions of PnPAL3 in black peppers

图7 胡椒PnPAL10 基因相对表达分析Fig.7 Analysis on relative expressions of PnPAL10 in black pepper
3 讨论
基因组测序技术的不断深入以及功能基因组学的日渐成熟为基因鉴定提供了便利条件,生物信息学的地位也日益重要。苯丙氨酸解氨酶基因家族应对各项生物和非生物胁迫,目前,叶小真等在桉树中克隆获得了PAL基因序列并分析其表达模式,发现该基因在高抗抗焦枯病菌品种中表达最高[16];吴远航等克隆了木薯的MePAL基因,并发现该基因可受低温胁迫诱导增强表达[17];高红胜等克隆了黄瓜的CsPAL基因,发现其在受白粉菌诱导后显著高于对照[18]。本研究在胡椒中鉴定到14 条PnPAL,其中7 条为完整结构,略多于拟南芥、水稻等家族,可能与胡椒基因组复制有关[19-20]。植物在进化过程中一般都比较保守,保守序列可能调控基因表达且含有相同保守序列的基因行使相同功能,甚至一些属于相同超家族的蛋白功能也相同[21]。笔者发现7 条胡椒PnPAL均含有保守的基序,推测它们可能行使相同功能。
由一个祖先通过基因复制或突变为适应不同环境而产生一个基因家族[22]。在植物进化过程中,某个祖先的倍增和变异可以产生多个家族成员,这些家族成员可能成簇的分布在一条染色体上,也可能分布在不同染色体上[23]。胡椒的PnPAL家族成员在染色体上不均匀分布,可能与基因组复制及染色体重复有关,基因组和染色体重复对物种进化有重要意义[24]。内含子的非法交换和变异可能是基因家族形成的原因[25],如植物12-氧-植物二烯酸还原酶(12-oxo-phytodienoic acid reductase,OPR)家族基因各内含子的位置和长度存在较大差异,故胡椒PnPAL成员之间基因结构的差异较大[26]。
苯丙氨酸解氨酶普遍存在于植物体中,鉴定到的胡椒PAL 蛋白理化性质、亚细胞定位、酸碱性、分子量、编码长度等不同,表明胡椒PnPAL有较高的遗传多样性。伴随着物种进化能力逐渐增强,环境适应能力愈强,均是遗传变异丰富造成的,故遗传变异是导致遗传多样性的根本原因[27]。近年来,越来越多研究表明基因表达调控对基因表达差异起重要作用[28]。在胡椒PnPAL中,各家族成员之间启动子略有差异,每个PnPAL在不同的生物或非生物胁迫中起作用;家族成员之间启动子的差异可能是进化过程中遗传变异导致的;由于各个家族成员所含启动子不同,启动子对基因表达的调控又有一定差异,进而表现出PnPAL家族成员参与应对不同的生物和非生物胁迫环境。
胡椒苯丙氨酸解氨酶家族各基因功能还尚不清晰。本文首次运用胡椒基因组信息,鉴定到胡椒的PnPAL家族成员,结合生物信息学方法分析胡椒PnPAL基因家族的结构差异,进化关系及表达模式并筛选到抵抗病原菌的关键家族成员,为进一步验证胡椒PnPAL基因功能及响应各项逆境研究提供基础。
