红蜻稚虫对浙江省不同类型水体污染的抗逆性适应表现
2021-08-13俞玲园季政权胡耀文徐风娇王琪玮王世贵
俞玲园,季政权,胡耀文,徐风娇,王琪玮,魏 莹,唐 斌,王世贵
(杭州师范大学生命与环境科学学院,杭州市动物适应与进化重点实验室,杭州 310036)
水体污染一直是人们关注的焦点问题,其中尤以重金属污染为重。重金属污染不仅加速了生态环境的破坏,同时给生物多样性造成了巨大的不利影响(何佳等, 2019),水体重金属污染监测与治理值得深入研究。水生昆虫包括双翅目、蜉蝣目、襀翅目、毛翅目、蜻蜓目等种类,它们对水体污染响应比较敏感,被广泛用来监测和评价水质,是水质生物监测的主要指示生物(Briers and Biggs, 2003;Yoshimura and Maeto, 2004;李金国等, 2007;Jeremiasonetal., 2016)。我国在上世纪 70-80年代开始探索利用包括摇蚊等水生昆虫在内的底栖无脊椎动物多样性指数等来评价河湖水质状况(刘保元等, 1981;杞桑, 1986),其后利用水生昆虫对多条河流的水质污染进行了评估(杨莲芳等, 1992;童晓立和胡慧建, 1995;何佳等, 2019)。
除了利用水生昆虫群落结构及多样性进行水质评价外,水生昆虫的生理响应指标也可以反映水体污染的程度,如抗氧化酶、P450解毒酶和热激蛋白等。抗氧化酶包括超氧化物歧化酶(SOD)、过氧化氢酶(CAT)和过氧化物酶(POD)等(Lushchak, 2011),在活性氧的清除及机体的保护性防御反应中发挥着重要作用。重金属离子、化学农药、石油等物质会影响或阻断呼吸链、电子传递链、酶促反应等体内正常细胞的代谢,诱发活性氧自由基的产生(Reactive oxygen species, ROS),活性氧自由基包括羟自由基(HO·)、羟过氧自由基(HO2·)、超氧阴离子(O2·-)、一氧化氮自由基(NO·)、过氧化氢(H2O2)和次氯酸(HClO)等(Jacob, 1995),为了减少活性氧自由基对机体造成的损害,机体通过增加抗氧化酶的活性以维持活性氧自由基在体内的平衡,从而减少氧化损伤(Slaninovaetal., 2009;Lushchak, 2011)。
热激蛋白70(Heat Shock Protein 70,HSP70)和热激蛋白90(Heat Shock Protein 90,HSP90)是热激蛋白家族中最保守、最重要的两种(马晨曦等, 2013)。HSP70和HSP90在昆虫抗逆境中的作用受到广泛关注,不仅在热应激的条件下对细胞有保护作用,在很多外源刺激(环境污染、农药、重金属等)下,都可以诱导生物体内HSP70和HSP90的大量合成与积累,在细胞应激后反应最明显,在维持细胞结构的稳定和提高细胞对应激原的耐受性中发挥重要作用(Limetal., 2005;杨静等, 2017)。HSP70和HSP90能被重金属离子诱导合成,而且这种诱导与重金属的浓度具有相关性,其表达量可以反映环境中的重金属含量水平(李铸衡, 2014)。多项研究表明HSP70和HSP90表达上调可作为某些重金属污染的生物标志(喻潇, 2013),可作为水体污染的检测指标(张令要, 2008)。
细胞色素P450(Cytochrome P450,P450)是一个古老的基因超家族(Nelsonetal., 1993),参与内源性物质和包括药物、毒素、环境污染在内的外源性物质的代谢,它的一个重要特征是它的可诱导性(Nelsonetal., 1993;Werck-Reichhart and Feyereisen, 2000)。细胞色素P450酶活性是一种便捷的污染物毒性评价机制和早期监测模型,通过研究污染物对生物体的毒性伤害和生物体早期所表现出的药物对细胞水平生物标记物的诱导状况之间的关系,并以毒性伤害为污染物的毒性评判标准,确定污染物与酶活力之间的量效关系,从而判断污染物在环境中的毒性及其残留量(Forkertetal., 1988;Okey, 1990)。
蜻蜓目昆虫分布十分广泛,其生活史特征独特,稚虫常栖息于水中砂粒、泥水或水草间,取食水中的小动物,是典型的水生昆虫,被视为非常适合作为研究水生昆虫与水体质量关系的模式昆虫(Jeremiasonetal., 2016),如长叶异痣蟌Ischnuraelegans对水体重金属Hg、Cd和Pb有明显的富集作用(席玉英和韩凤英, 2000;韩凤英和席玉英, 2001;韩凤英等, 2002)。因此,研究蜻蜓幼虫对水体污染的生物指示作用有着重要意义。本文以浙江地区普遍存在的红蜻Crocothemisservilia作为试验对象,分别对浙江杭州、绍兴等地不同污染类型水域中的红蜻稚虫进行调查取样,对红蜻稚虫的SOD、CAT酶活力以及HSP70、HSP90和P450家族CYP303a1的表达水平进行比较分析,结合水体质量各项理化指标的测定,分析水体理化指标及重金属含量对蜻蜓稚虫的影响,明确其响应重金属污染胁迫的抗逆性适应表现及潜在机制,为利用蜻蜓稚虫作为水质监测的生物指示物提供参考依据。
1 材料与方法
1.1 采样点布设与样品采集
采样点的布设主要考虑了浙江省内不同地区的污染状况,如杭州、绍兴和宁波工业废水排放严重,是浙江省工业废水排放的主要区域(蔡临明, 1999)。皮革加工业、纺织业和塑料制品业等第二产业是温州市的支柱,企业生产工艺落后是水体污染主要污染来源(徐越等, 2014)。高耗能、高排放的粗放型生产企业使宁波市的河流及其他水均受到不同程度的污染,其工业区更是使水体污染严重(王桂林等, 2008;周正勇, 2016)。绍兴市水污染主要为有机污染, 以印染为主(祁华宝和鲁玉龙, 1995;陈昕和章科锋, 2009)。台州地区是国内电子产品拆解中心的典型,其造成的重金属污染已处于较高水平(潘海燕等, 2015)。基于此,本次调查分别在浙江省内的杭州市下沙高教园区杭州师范大学下沙校区鸽子湖(N30°19′11″, E120°23′17″)、温州市平阳县水头镇下林村河道(N27°38′44″, E120°20′22″)、台州市临海区下沙周村河道(N28°53′32.1″, E121°13′21.1″)、绍兴柯桥安昌街道九鼎村河道(N30°10′3″, E120°28′19″)和宁波市镇海区骆驼街道英雄河(N29°58′59.1″, E121°33′33.8″)等5个地区选取一处较大的水体作为采样地点。
采样时间为2016年7月中旬和2018年8月中旬。采样时第一时间测取各采样点水体的物理指标(水温、压强、电导率EC、氧化还原电位Eh、pH、溶解氧DO),并用2.5 L有机玻璃采水器采集表层水样,每采样点各取3瓶水样。采样后将水样放置在冷藏箱中,做好水样的储存和低温运输工作,以防水样变质。蜻蜓稚虫的采集参考Barbour等(1996)的采样方法并适当改进,使用D形抄网(MY-031,0.3 m宽,40目尼龙纱网)取样调查,分别在堤岸、缓水区、稳水区、大型水草根部和淤泥等蜻蜓稚虫栖息的不同栖境类型,按照栖境出现的比例分配取样数,分别用D型网紧贴水底基质清扫0.5 m长区域,将所有基质等抄入网内,进一步用40目分样筛分离出其中的蜻蜓目稚虫。每采样点清扫20次,总采样面积约3.0 m2。将红蜻稚虫放入样品低温运输箱中保存,尽快带回实验室后-80℃保存,其余蜻蜓稚虫用50%酒精保存。
1.2 实验内容与方法
1.2.1水体理化性质、重金属含量的测定
测定分为现场和室内测定。现场测定水体的水温、压强、电导率(EC)和氧化还原电位(Eh)、pH和溶解氧(DO),pH值用酸式笔式酸度计(HI 98127)测定,水温和溶解氧使用哈希HQ40D型溶氧分析仪测定。室内测定内容有总氮(TN)、总磷(TP)、高锰酸盐指数(CODMn)、叶绿素a(Chl-a)、透明度(SD)和6种重金属(Cu2+、Zn2+、Cd2+、Pb2+、Cr6+和总铁)含量。TP使用钼酸铵分光光度法(GB 11893-89),CODMn使用滴定法(GB 11892-89),叶绿素a采用浮游植物荧光仪(Phyto-PAM)法,SD采用赛氏盘法(顾晓光等, 2005)。
Cu2+、Zn2+、Cd2+、Pb2+采用原子吸收分光光度法(GB 7475-87),Cr6+的测定依据国家标准二苯碳酰二肼分光光度法(GB 7467-87),总铁的测定依据其他国家标准邻菲啰啉分光光度法(HJT 345-2007),水质的消解处理依据其他国家标准微波消解法(HJ 678-2013)进行处理,TN使用碱性过硫酸钾消解紫外分光光度法(HJ 636-2012),采用TLI综合营养指数法运用TP、TN和CODMn等对调查区域水质状况进行评价(陈丽华等, 2018)。
1.2.2蜻蜓稚虫鉴定和实验物种的选择
对采集的蜻蜓稚虫进行数量统计及鉴定,其中红蜻稚虫鉴定到种,其余稚虫均鉴定到科(赵修复, 1995)。红蜻稚虫为5个调查样点的共有种类,因此选择红蜻Crocothemisservilia作为本次调查的实验物种,进一步分析其在污染胁迫下的抗逆性生理适应表现。
1.2.3超氧化物歧化酶和过氧化氢酶的酶活性测定
按照重量(g)∶体积(mL)=1∶19的比例加入19倍生理盐水,首先使用超声破碎仪冰水浴破碎红蜻稚虫,离心取上清,得到5%组织匀浆液。超氧化物歧化酶(Superoxide Dismutase, SOD)活性测定采用亚硝酸盐法,过氧化氢酶(Catalase, CAT)活性测定采用钼酸铵法,试剂盒购于南京建成生物工程研究所。总蛋白含量采用BCA蛋白试剂盒(TaKaRa)进行测定,试剂盒购于宝生物工程(大连)有限公司。酶活性计算方法参照试剂盒配套的使用手册完成,酶活单位用U/mg蛋白质表示。
1.2.4HSP70、HSP90和CYP303a1的基因表达分析
各采样点选取大小龄期相似的红蜻稚虫各 3头,分别置于1.5 mL EP管中,置于冰上,加100 μL Trizol(Lifetech Scientific Corporation, USA),电动匀浆器研磨;补足Trizol至1 mL,用力振荡3 min,室温静置5 min;加入200 μL氯仿,混匀后静置10 min,于4℃ 12 000 g离心15 min;转移上清至新的1.5 mL EP管,加入500 μL异丙醇,上下颠倒混匀,室温静置10 min;离心,弃上清,并加入1 mL DEPC水配制的75%乙醇,将沉淀吹离管壁,洗涤沉淀,4℃ 7 500 r/min离心5 min;倒去上清,重复前一步骤;离心后倒掉上清,将离心管倒置在超净台上,加入适量的RNA Free H2O水溶解,吹打混匀,于-80℃保存。提取后用1%琼脂糖检测总RNA的质量,然后用微量测定分光光度计(NanoDropTM2000)测定提取RNA的浓度及纯度。使用Prime Script©RT Reagent Kit with gDNA Eraser试剂盒(TaKaRa)配置体系,进行cDNA第1链的合成,于-20℃保存待用。
利用Prima 5.0设计Actin、HSP70、HSP90和CYP303a1定量引物如表1。采用实时荧光定量PCR法(Real-time fluorescent quantitative PCR, qRT-PCR)测定各水体中红蜻稚虫的HSP70、HSP90和CYP303a1的基因相对表达量。按以下体系进行反应:SYBR GREEN 5 μL,primer F(10 pmol/μL)0.4 μL,primer R(10 pmol/μL)0.4 μL,模板cDNA 1 μL,RNase Free ddH2O 3.2 μL。反应程序:94℃预变性3 min;94℃变性5 s,59℃退火延伸25 s,循环40次;最后绘制熔解曲线。采用2-ΔΔCT法计算qRT-PCR试验中的基因相对表达量,计算公式为:基因相对表达量=2-ΔΔCT。其中,ΔΔCT计算公式为:ΔΔCT=ΔCT(试验组)-ΔCT(对照组);试验组或对照组的ΔCT计算公式为:ΔCT=CT(目的基因)-CT(内参基因)。
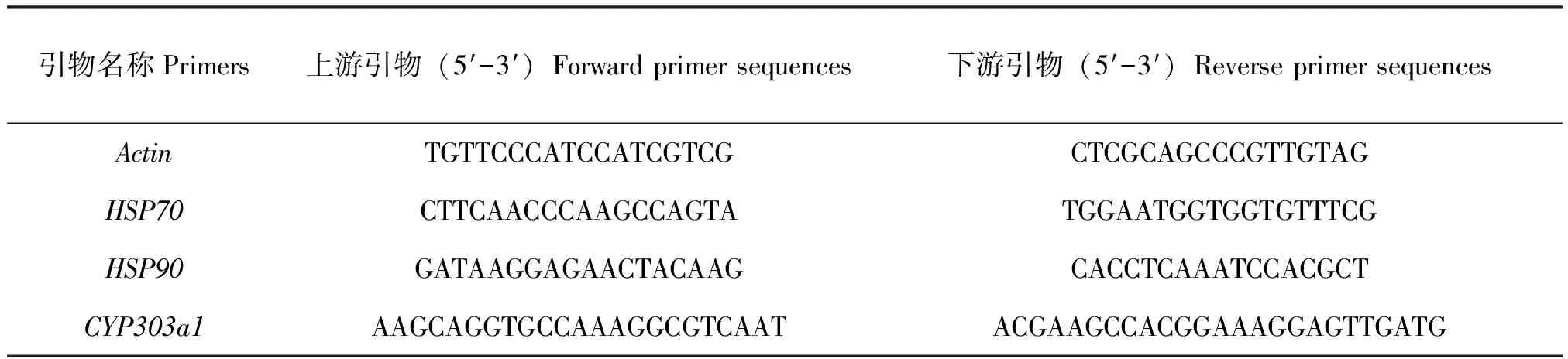
表1 荧光定量PCR特异性引物
1.2.5数据分析
对数据进行正态分析和方差齐性分析。实验数据统计分析通过SPSS 20.0完成,采用单因素方差分析(One-way ANOVA)对水质理化系数、酶活力和基因表达量进行显著性差异分析,采用Duncan氏检验,其中P<0.05被认为有显著性差异,P<0.01时,则说明差异极显著;采用皮尔逊相关系数(Pearson correlation coefficient)对红蜻稚虫SOD和CAT酶活力,HSP70、HSP90和CYP303a1基因的表达与环境因子的关系。用Origin 2017进行柱状图绘制。
2 结果与分析
2.1 环境因子分析
2.1.1水域水体的理化性状和富营养化水平
对各水域水体的理化性状进行测定并根据地表水环境质量标准(GB 3838-2002)进行分类,各地水质依DO指标显示杭州属于I类水质,温州属于III类,台州属于II类,绍兴属于II类,宁波属于III类(表2)。
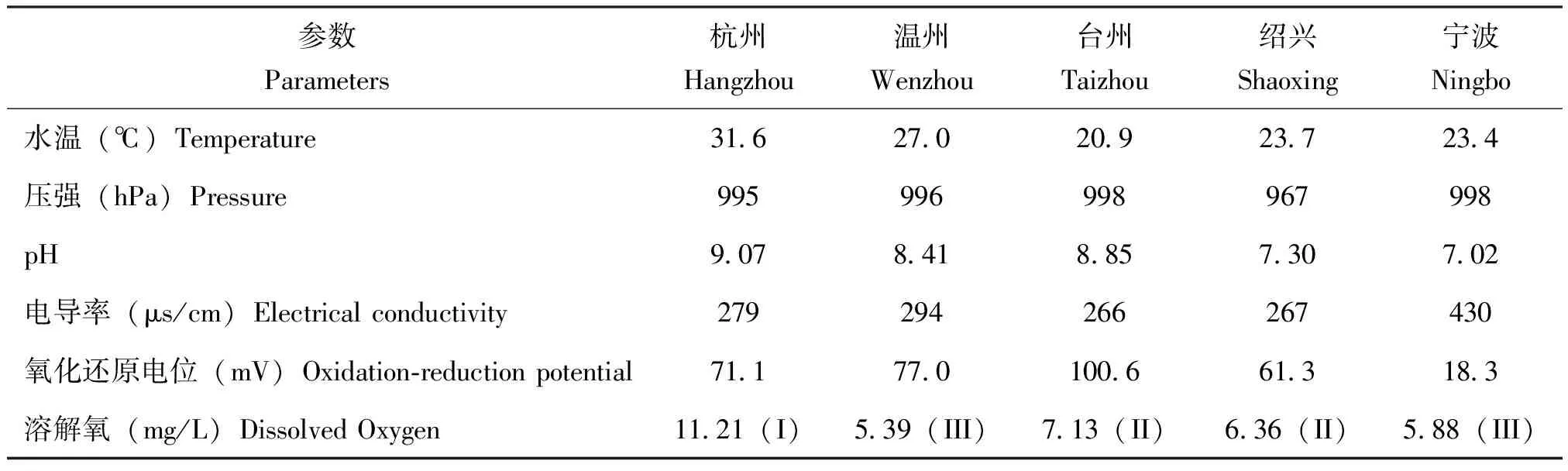
表2 浙江省各采样点水体的理化性状
不同水域水体的营养化水平有显著的差异(图1-A~E),其中杭州采样点的TP和叶绿素a皆为最低,SD最高;温州采样点TN和CODMn含量均最高;宁波采样点TP含量最高,SD最低;台州采样点TN和CODMn低于其他3个地区。根据地表水环境质量标准(GB 3838-2002),各地水质依TN指标显示杭州属于II类水质,温州属于IV类,台州属于II类,绍兴属于II类,宁波属于III类;依TP指标则杭州属于I类水质,温州属于III类,台州属于IV类,绍兴属于IV类,宁波属于V类;依CODMn指标显示杭州属于I类水质,温州属于IV类,台州属于I类,绍兴属于III类,宁波属于III类。从图1(F)可以看出,温州、宁波、绍兴采样点的TLI富营养化指数都高于50。由此得出,温州和宁波采样点为重度富营养化,绍兴采样点为中度富营养化,杭州和台州采样点为中营养,各采样点之间水质差异较大。
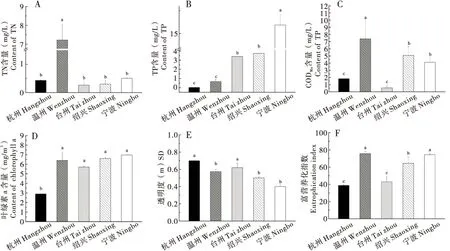
图1 浙江省各采样点总氮、总磷、高锰酸钾指数、叶绿素a、透明度和TLI富营养化指数Fig.1 Eutrophication index and TN, TP, CODMn, chlorophyll a, SD and TLI eutrophication indices in Zhejiang Provaince注:图中数据为平均值±标准误(n=3),柱上不同字母表示经Duncan’s检验后差异显著(P<0.05)。下图同。Note:Data in the figure are mean±SE(n=3), and different letters above bars indicate significant difference by Duncan’s test(P<0.05).The same for following Figures.
2.1.2各水域水体中重金属含量水平
各水域水体中总铁、Cu2+、Zn2+、Cd2+、Pb2+和Cr6+含量存在地区间差异(图2)。比较而言温州采样点的Cd2+、Cr6+和总铁含量较高,台州采样点的Cd2+和Cu2+的含量偏高,绍兴采样点含量偏高的金属离子有Cr6+、Pb2+和Zn2+,宁波采样点Zn2+含量最高,杭州采样点的Cd2+、Cr6+和Pb2+显著低于其他4个采样点。根据地表水环境质量标准(GB 3838-2002)对各采样点水体的重金属含量进行分析,温州和台州采样点Cr6+较其他采样点高,约为0.003 mg/L,但仍属于I类水质;温州和绍兴采样点Cr6+较其他采样点高,约为0.03 mg/L,但仍属于II类水质;Cu2+含量最高的台州采样点也属于II类水质;Pb2+含量较高的绍兴和宁波也仍属于III类水质;连Zn2+含量最高的宁波采样点也属于II类水质。综合分析可得:杭州采样点属于I类水质,温州采样点属于III类水质,台州采样点属于III类水质,绍兴采样点属于I或II类水质,宁波采样点属于II类水质。
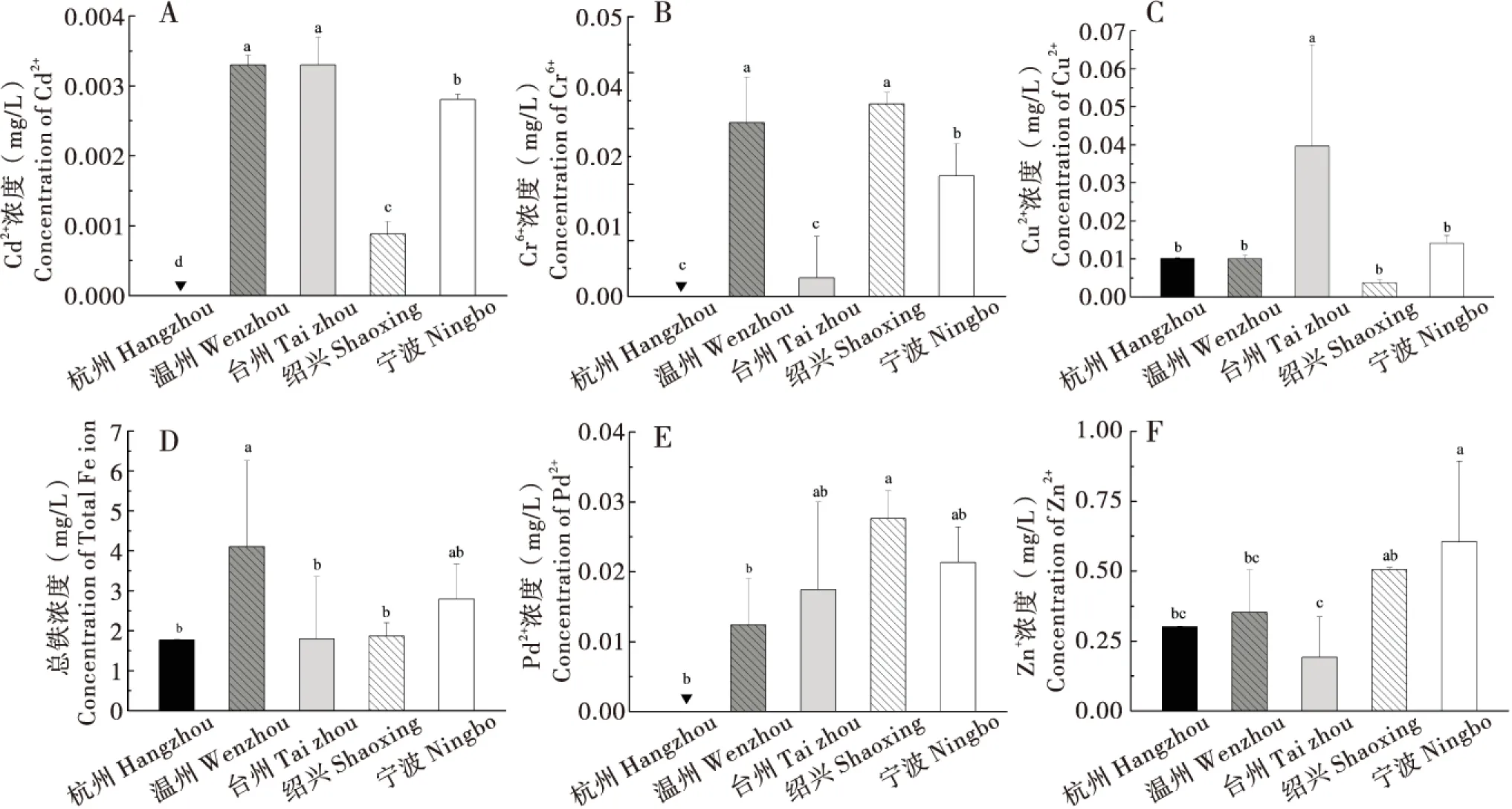
图2 浙江省各采样点重金属离子浓度Fig.2 Heavy metal ion in Zhejiang Province注:图中“▼”表示水样中重金属未检出。Note:“▼” means that heavy metal has not been detected in the samples.
2.2 各水域水体中采集的蜻蜓稚虫分科鉴定
本次调查共采集到蜻蜓稚虫169头,经分类鉴定分属7科(蜻科Libellulidae、春蜓科Gomphidae、大蜓科Cordulegastridae、蜓科Aeshnidae Rambur、色蟌科Calopterygidae、扇蟌科Platycnemididae和丝蟌科Lestoidea)。其中杭州采样点采集到4科31头,隶属于蜻科、春蜓科、大蜓科和色蟌科;温州采样点采集到2科40头,隶属于蜻科和春蜓科,其中采集红蜻23头,为所有采样点中最多;台州采样点采集到的蜻蜓数目最多,共5科65头,隶属于蜻科、春蜓科、蜓科、扇蟌科和丝蟌科;绍兴和宁波都只采集到蜻科、春蜓科和扇蟌科3类蜻蜓。其中红蜻是5个采样点的共有种类。
2.3 不同水域水体中红蜻稚虫超氧化物歧化酶和过氧化氢酶的活力
不同水域水体中红蜻稚虫SOD和CAT的活力有显著差异(图3)。台州采样点红蜻稚虫的SOD和CAT酶的活力最高,温州采样点的红蜻稚虫略低于台州采样点,杭州采样点红蜻稚虫的酶活力最低。结果表明,在各采样点采集的红蜻稚虫的SOD和CAT活性差异显著,且SOD和CAT酶活力在各采样点所表现的趋势相同。

图3 浙江省各采样点红蜻稚虫SOD、CAT活性Fig.3 Enzymatic activity of Crocothemis servilia nympho SOD and CAT in Zhejiang Province
2.4 不同水域水体中红蜻稚虫HSP70、HSP90和CYP303a1的基因表达分析
以杭州地区采集的红蜻稚虫作为参照,通过计算得出各采样点红蜻稚虫HSP70、HSP90和CYP303a13个基因的相对表达量(图4)。绍兴采样点CYP303a1和HSP90基因的相对表达量明显高于其它3个采样点,而HSP70基因的相对表达量在温州采样点红蜻稚虫体内表达量最高。水质最好的杭州地区采样点红蜻稚虫HSP70、HSP90和CYP303a13个基因的表达量都是各组间最低值。通过统计分析得出,各地区采集的红蜻稚虫体内HSP70、HSP90和CYP303a13个基因的表达量存在地区间显著性差异。

图4 浙江省各采样点红蜻稚虫HSP70 、HSP90和CYP303a1相对表达量Fig.4 Relative expression leval of Crocothemis servilia nympho HSP70、HSP90 and CYP303a1 in Zhejiang Province
3 结论与讨论
3.1 不同水域水体污染状况及其与蜻蜓稚虫之间的关系
根据浙江省环境厅于2019年11月公开的浙江省地表水环境质量月报所示:全省地表水总体水质为良。221个省控断面中,Ⅰ类占12.2%,Ⅱ类占38.9%,Ⅲ类占33.0%,Ⅳ类占12.2%,Ⅴ类占3.2%,劣Ⅴ类占0.5%;满足水环境功能区目标水质要求断面占86.9%。水体的主要污染指标为总磷、氨氮、五日生化需氧量和化学需氧量,而造成浙江省农村河道工业污染源的类型因各地的经济支柱产业结构不同而有差异,主要污染源来自采矿业、纺织业和皮革毛皮及制品业、食品饮料制造加工业等;农业污染源则以农业种植为主,其它有农村生活污水及固体废弃物、分散式畜禽养殖等(卢卫和应聪慧, 2009)。
就本研究采样点实地考察发现:宁波镇海采样点则位于镇海石油炼化企业污染辐射区内,而温州市平阳水头镇以皮革产业出名,推测温州和宁波采样点的富营养化主要与当地的工业污水、农业污水和生活污水有关(张松达等, 2010;金矛, 2015)。绍兴柯桥采样点附近有绍兴第一印染厂、绍兴萧绍印染厂、大和集团印染三厂等多家纺织印染企业,水体中度富营养化程度与纺织业的有机污染有关(祁华宝和鲁玉龙, 1995;陈昕和章科锋, 2009;吴绩新等, 2018)
本次实验对各地区采样点的6种重金属含量进行了测定。绍兴采样点含量偏高的重金属离子有Cr6+、Pb2+和Zn2+,应该与当地发达的纺织印染产业有关(李萍等, 2012);温州平阳水头镇采样点的Cr6+浓度比其他地区较高,应该与当地发达附近的皮革制造业有关(杨斌等, 2016),同时Cr6+在台州和宁波两个地区都较高,这与Cr6+被广泛应用于电镀工业、化工业、电子业、冶炼和印染等行业有关。台州采样点Cu2+和Cd2+含量要高于其它几个地区,这可能和该地区的电子回收产业有关(潘海燕等, 2015)。宁波镇海地区水体中Zn2+含量最高,当地发展电镀企业,而锌是主要的原材料之一(王宗雄, 2012;杨斌等, 2016)。
水质的差异直接或间接影响水生昆虫的群落结构,比如氮浓度、磷浓度、生化需氧量与水生昆虫密度呈极显著正相关,与水生昆虫的种数呈极显著负相关(罗清荣, 2012),TN和Cd2+等环境因子也会影响物种的分布(刘祥等, 2016),而底层氧含量往往直接决定了水生昆虫的分布和密度(陈含墨等, 2019)。本次调查在水质较好的杭州和台州采集到的蜻蜓种类最多,而温州、宁波和绍兴等地采集到的蜻蜓种类较单一,也支持这一观点。张潋波等( 2013)在研究浙江省钱塘江中游流域水生昆虫功能多样性对土地利用变化的响应时,发现人类对土地的利用会导致溪流生态质量退化,水体中营养盐浓度和电导率明显上升,溶氧含量明显下降,一些生物学性状的改变,进而影响到水生昆虫群落及其功能多样性,这些研究都说明人类活动所产生的水体污染已经严重影响了水生昆虫的群落结构和多样性。
本次调查在5个地区采样点都采集到了红蜻稚虫,其中温州样点采集的红蜻稚虫最多,而水质分析显示该采样点水体中Cd2+、Cr6+、TN和CODMn都较高,且水体富营养程度为重度富营养,推测红蜻稚虫对不同水体均有较强的抗逆性适应能力,即使在富营养化程度较高的水体中也能较好的生存。
3.2 红蜻稚虫SOD、CAT酶活力与环境因子的关系
当生物体受到污染物刺激时,会产生大量的活性氧,如果不及时去除,会对生物体造成损伤。当昆虫暴露在污染物下时,机体内的SOD和CAT会同时发挥作用来消除过多的自由基从而达到让自己能够生存下去的目的(Subalaetal., 2017;Zhangetal., 2007;Zhangetal., 2017)。如果长期在含较低浓度污染物的环境下暴露,生物体内的SOD和CAT酶活力将会被诱导(宋志慧和王庆伟, 2011)。吴国星等(2008)研究了棕尾别麻蝇Boettcheriscaperegrina幼虫的脂质过氧化作用与重金属镉和铜存在着浓度依赖,其体内的SOD和CAT酶活力随重金属浓度升高而明显被抑制。王慧等(2006)研究几种蝇类在镉和铜胁迫下的抗氧化酶活性变化发现:镉和铜明显抑制蝇类幼虫的SOD和CAT酶活性,且随着重金属的浓度升高其抑制作用增强。在对各个采样点红蜻稚虫体内SOD和CAT进行测定时,发现不同采样点的红蜻稚虫其体内抗氧化酶活力存在显著差异,可能与红蜻稚虫对各自水体环境的胁迫响应有关。当外界Cr6+浓度较高时,红蜻稚虫体内的SOD和CAT酶活力上升(P<0.05),SOD和CAT酶活力和Cr6+呈正相关(r>0);当Cu2+含量较高时,CAT酶活力上升(P<0.05),CAT酶活力和Cu2+呈正相关(r>0)。本研究还发现尽管不同水体内红蜻稚虫体内SOD和CAT的酶活力不同,但是它们在各地有相似的表现,都呈现出台州>温州>宁波>绍兴>杭州的趋势,进一步证明红蜻稚虫体内的SOD和CAT酶活力与其生活的水体环境具有一定的联系。结合各地环境因子污染情况,本研究发现SOD与CAT酶活力最高的两个地区所对应的最突出的环境污染因子是Cr6+,而对其它重金属的反应并不强烈,由此可以推测红蜻体内的SOD和CAT酶活力在一定程度上能反应出某一水体的Cr6+污染情况。
3.3 红蜻稚虫HSP70、HSP90、CYP303a1基因的表达与环境因子的关系
HSP家族是生物体中重要的伴侣蛋白,炎症、热应激、缺血、重金属胁迫等一系列的因素都会导致其mRNA的表达(计红等, 2005),HSP70在被诱导后表达水平甚至可增加10~100倍(陈兰英等, 2004;彭乔烽等, 2019)。有研究表明在pH和重金属的暴露下,凡纳滨对虾Litopenaeusvannamei的HSP60、HSP70、HSP90基因会进行不同程度的特异性表达(Qianetal., 2012)。本研究发现,红蜻稚虫在富营养化水体中,通过不同片段长度的HSPs的表达对环境变化作出相应反应,当污染物主要为氮素时,红蜻稚虫的HSP70表达能力上升;当污染物主要为磷元素时,红蜻稚虫的HSP70表达能力下降,HSP90表达能力上升;当污染物为还原性物质时,红蜻稚虫的HSP90表达能力下降。通过测定不同片段长度的HSPs的表达量,可以了解到当前水体的主要污染物质。Mahmoud等研究发现,在无机磷暴露下,鸡体内的HSP90基因有明显的升高,这与本实验的结果相一致(Mahmoudetal., 2004)。通过相关性分析得出,HSP90表达量随TP和CODMn浓度呈正相关关系(r>0),随着水体中TP、CODMn浓度的上升,HSP90基因的表达量随之上调(P<0.05);HSP70的表达量随Pb2+浓度上升而上调(P<0.05),且与Pb2+浓度间呈正相关(r>0),这与邓茗芩关于秀丽影杆线虫Caenorhabditiselegans的研究结果也是一致的(邓茗芩, 2014)。
细胞色素P450酶系(CYP450)是一类重要的多功能氧化酶(Guengerichetal., 2016),参与生物体内各种内源和外源性物质的氧化代谢,大部分药物和外源污染物的代谢都与P450酶系有关(Vaccaroetal., 2002;Lavilleetal., 2004)。通过分析,红蜻稚虫CYP303a1基因的表达水平与水体中Cr6+含量呈负相关关系(r<0),在长期Cr6+胁迫的作用下,会抑制红蜻稚虫CYP303a1基因的表达(P<0.05);CYP303a1的表达量与TP和Cd2+呈正相关(r>0),随着这两种环境因子的浓度上升,会促进红蜻稚虫体内CYP303a1基因表达上调(P<0.05)。
3.4 不同类型水体污染下蜻蜓稚虫的抗逆性表现
通过对水体理化指标的测定,以及红蜻稚虫酶活性和基因表达的测定等,在皮革废水污染下,红蜻稚虫体内的SOD、CAT和P450能帮助红蜻稚虫适应Cr6+浓度较高的水体中。Loumbourdis等(2006)认为Cr6+进入机体后可被体内的抗氧化剂(如谷胱甘肽、甘酸等)迅速还原为低毒的Cr3+,而微量Cr3+对于机体的代谢活动是必不可少的,因此推测低浓度Cr6+不会对蜻蜓稚虫的生长发育产生抑制作用。在高浓度时,进入体内的Cr6+含量长期偏高,会抑制SOD、CAT活性和CYP303a1的基因表达能力。在富营养化水体中,红蜻稚虫通过表达不同片段长度的HSPs,来应对不同类型的富营养化因素,从而更好地适应生活环境的变化。
本研究在通过对红蜻幼虫生存的水体环境和体内的抗氧化酶SOD和CAT以及HSP70、HSP90和CYP303a1分析可得以下结论:
1)红蜻稚虫体内的SOD和CAT酶活力与水体中的重金属Cr6+离子浓度密切相关,可在一定程度上反映出水体的Cr6+污染状况;
2)红蜻稚虫HSP70 的表达量与水体TN、TP和Pb2+浓度间呈正相关,同时HSP90的表达与水体TP和CODMn浓度正相关,可反映水体富营养化的主要污染物类型;红蜻稚虫CYP303a1的表达水平与水体中Cr6+含量呈负相关,而与TP和Cd2+呈正相关。推测HSPs和CYP可能与稚虫抵抗不同类型的重金属或富营养化污染有关系。
