谷子产量和品质相关性状的杂种优势及遗传特性分析
2021-08-10史关燕王啸旗韩渊怀杨成元麻慧芳赵雄伟乔治军
史关燕, 王啸旗, 韩渊怀, 杨成元, 麻慧芳, 赵雄伟, 乔治军
谷子产量和品质相关性状的杂种优势及遗传特性分析
史关燕1, 王啸旗2a, 韩渊怀2a, 杨成元1, 麻慧芳1, 赵雄伟2b*, 乔治军3*
(1. 山西农业大学经济作物研究所, 山西 汾阳 032200;2. 山西农业大学, a. 农学院; b. 生命科学学院 山西 太谷 030801;3. 山西农业大学农业基因资源研究中心, 山西 太原 030000)
为了解谷子()两系杂交后代的产量和品质相关性状的杂种优势,以3个高度雄性不育系为母本(A1、A2和A3),6个抗除草剂恢复系为父本(R1、R2、R3、R4、R5和R6),采用不完全双列杂交(NCⅡ)设计配制了18个组合,分析组合的株高、粒重和蛋白含量等11个性状的配合力和杂种优势特性。结果表明,不同亲本间及杂交组合间的性状均存在显著差异。其中,两系杂交组合的小区产量与千粒重、单穗粒重、株高、穗长显著相关,而与小米的品质(粗蛋白、粗脂肪、总淀粉)无显著相关。同一性状不同亲本的一般配合力(GCA)效应值不同,A1 (不育系)和R1 (恢复系)的产量和品质性状的GCA正效应值较大,是改良谷子高产优质的潜在优良亲本。而杂交组合的特定特殊配合力(SCA)效应值与亲本GCA效应值表现并不一致,其中A1×R5、A1×R6、A2×R1、A2×R4的SCA效应值较高,属于优异组合。此外,各性状的广义遗传力和狭义遗传力均大于65%,主要受加性效应和非加性效应共同控制。谷子杂交组合的株高、穗重、小区产量具有显著的中亲优势,部分杂交组合具有较强的超亲优势。株高、穗长、穗码、千粒重、脂肪的狭义遗传力较高,加性遗传作用较大,而穗重、茎节数、蛋白非加性遗传作用更为突出。虽然产量性状和品质性状的杂种优势特性不同,但均由加性效应与显性效应共同作用决定。
谷子;产量;品质;杂种优势;配合力;遗传力
杂种优势是自然界普遍存在的生物学现象, 极大促进了作物产量和品质提升,已在玉米()、水稻()、小麦()、棉花()等[1–4]重要农作物生产上得到广泛应用。成功利用杂种优势的关键问题在于如何确定和选择具有产出高优势杂交组合的潜力亲本。然而,大部分具有优良农艺性状的亲本并不能完全把这些优异性状遗传给子代。为了解决这一问题,在植物杂交育种中经常将配合力效应值作为筛选具有潜力亲本和特定组合的标准。如Chen等[5]利用96个栽培稻和4个雄性不育系用NCII组配384个杂交组合,对12个农艺性状的一般配合力(general combining ability, GCA)和特殊配合力(special com- bining ability, SCA)效应值进行分析,结果表明GCA和SCA)不同农艺性状的贡献不同,通过GCA效应值的QTL定位方法挖掘到、和优势等位基因,能解释30.03%的高产量GCA效应值表型变异率。因此,杂种优势是GCA和SCA位点效应积累的结果,通过选择配合力高的亲本能加快高产优质新品选育进程。
相比其他作物的栽培地区,中国是世界上唯一一个大规模、系统性研究谷子()种质遗传育种的国家[6]。而且谷子具有抗旱、耐逆等特性,是旱作生态农业不可替代的作物,也是适宜在西北干旱地区种植的主要杂粮作物之一。由于谷子产量提高不明显、经济效益较低等原因,每年的种植面积逐年降低。1942年谷子种植面积从9.21×106hm2减少到2013年的6.7×105hm2[7]。虽然谷子是自花授粉作物之一,其花器小、人工杂交难度大,但科学家经过多年努力,通过两系配套技术完成了杂交种的组配,很好地利用了杂交优势的生物学现象。如赵治海等[8]利用谷子雄性不育系选配了14个杂交种(张杂谷系列),在中国西北、华北部分地区种植较为广泛,并在非洲试种;王玉文等[9]也培育了长杂谷系列杂交谷子。因此,采用两系配套已实现谷子产量杂种优势的大规模利用。此外,对于作物杂种优势研究主要集中于产量相关性状方面,而现在大众的市场需求已经转向品质。品质是一个涉及多个指标的综合性状,研究不同组合间的品质性状杂种优势特性对配置高产优质新品种有着非常重要的作用。王昌华等[10]对水稻品质性状杂种优势、配合力和遗传效应的研究表明,不同组合间、不同恢复系间和不育系间的配合力差异达极显著水平, 杂交粳稻品质性状均受亲本的影响,且受恢复系的影响大于不育系。目前缺乏对谷子产量和品质相关性状杂交优势的研究。鉴于此,本研究以18个杂交组合及其双亲作为试验材料,通过测定产量性状和品质性状,研究不同杂交组合产量和品质性状的中亲优势和超亲优势,进一步了解亲本一般配合力和杂交组合的特殊配合力,为探索谷子两系杂种优势规律、指导谷子高产优质强优势组合选配、提高杂交育种效率提供一定的参考。
1 材料和方法
1.1 材料
本研究以谷子()的3个高度雄性不育系(A1、A2、A3)为母本,6个抗除草剂恢复系(R1、R2、R3、R4、R5、R6)为父本(表1)。供试材料均由山西农业大学(山西省农业科院)经济作物所提供。
1.2 方法
2018年冬在海南育种基地,采用不完全双列杂交设计(NCII),以3个不育系为母本,6个恢复系为父本进行组配,产生18个杂交组合(F1)。2019年4月下旬,将18个杂交组合F1及其亲本种植于山西农业大学经济作物所试验地,试验地前茬为蔬菜,采用随机区组设置共设3个小区重复,每小区面积为2 m×2 m,6行区,后期留苗密度设置为3.45× 105ind./hm2,按常规生产进行田间管理。待谷子成熟期后,在特定小区的第2、3、4、5行采收20株,晾干后测定株高、主茎节数、穗长、穗粗、单穗重、千粒重、穗码数,计算小区产量。参照国家标准GB/T 5511-2008谷物和豆类氮含量和粗蛋白质含量凯氏法测定谷子脱壳后小米的蛋白质含量;参照国家标准GB 2906-1982谷类、油料作物种子粗脂肪测定方法测定小米的脂肪含量,并使用紫外可见分光光度计比色法测定小米的总淀粉含量。
1.3 数据的统计分析
利用Excel软件对数据进行整理和分析,利用SPSS软件对亲本和不同杂种的产量和品质相关性状进行方差分析和LSD (least significant difference)多重比较分析。中亲优势(%)=100×(F1-MP)/MP, 超亲优势(%)=100×(F1-HP)/HP,F1为杂交组合F1代的表型,MP和HP分别为双亲表型性状的平均值和高值亲本的表型[11]。然后根据国家和各省谷子杂交种的审定标准,将特定组合的产量超亲优势值≥20%认定为强优势组合,5%~20%为中优势组合,0%~5%为弱优势杂交组合[12–13]。采用R语言som- mer软件包[14]估算亲本的一般配合力(GCA)、杂交组合的特殊配合力(SCA)效应值以及遗传相关参数。
表1 亲本材料
2 结果和分析
2.1 亲本的产量和品质性状的比较分析
对3个不育系和6个恢复系的田间产量性状和品质性状进行分析(图1),结果表明不同材料间在同一性状上存在显著差异。除了千粒重、总淀粉含量外, 其他性状在不同亲本间的变异系数为25.88%~ 105.52%,其中变异系数最高的为小区产量。此外,不育系A2和A3均属矮秆型,其平均株高显著低于恢复系的29.06%。在6个恢复系中,R5和R6的株高显著低于其他恢复系,而R4的穗长、单穗重、产量显著低于其他恢复系。不育系和恢复系的蛋白质、脂肪含量和总淀粉含量也存在显著差异, 如不育系的总淀粉含量显著低于恢复系。在恢复系中,除R1的脂肪和总淀粉含量有显著差异外,其他恢复系的差异均不显著。
2.2 杂交组合的产量和品质性状的杂种优势分析
通过分析两系杂交组合的产量和品质性状杂种优势表现(表2),结果显示杂交组合存在广泛的中亲优势和超亲优势,不同性状间存在较大差异,同一母本或父本杂交后代也存在明显的差异。从变异系数来看,杂交组合的穗重变异系数最大(61.24%),穗重为20.46~42.34 g;千粒重的变异系数最小(15.80%),为2.82~3.34 g。从株高的中亲优势来看,18个组合均表现出正向中亲优势(2.17%~37.02%),且组合间差异较大。其组合A2×R1中亲优势最大,组合A3×R3中亲优势最小。从株高高亲优势来看,组合的超亲优势为–19.99%~10.43%,变异系数为–11.14%,其中8个组合表现出超亲优势,组合A1× R4 (10.44%)、A1×R6 (10.05%)和A2×R1 (9.39%)的株高超亲优势最高,最低的组合有A3×R2 (–19.99%)、A3×R3 (–18.82%)、A2×R2 (–11.98%)和A2×R3 (–10.43%)(表3)。
杂交组合的穗长、单穗重和小区产量中亲优势变异系数分别为–2.61%~25.24%、5.07%~93.24%、22.72%~97.15%。除组合A3×R1外,其他组合的中亲优势均为正值(表2)。而穗长、单穗重和小区产量的超亲优势在组合间差异明显,分别为–15.19%~ 24.22%、–18.61%~75.68%和–15.58%~46.03%,以单穗重超亲优势的平均值最大。正向超亲优势组合超过8个,其中组合A2×R3、A2×R4和A3×R4的中亲优势和超亲优势较大(表3)。另外,组合A2× R6、A2×R1、A3×R2、A3×R5和A3×R6的超亲优势并不高,其平均值为-8.59%。品质性状的中亲优势和超亲优势平均值来看,蛋白质含量分别为–4.79%和–10.44%,脂肪含量分别为3.43%和–4.50%,淀粉含量分别为–1.46%和–6.79%。虽然品质性状的超亲优势均值为负值,但一些特异组合有正向的优势, 如蛋白质超亲优势为正值的组合有A2×R1和A2× R2,淀粉超亲优势为正值的有4个组合,表明就这些品质性状在杂交谷子配组上是可以选择的。
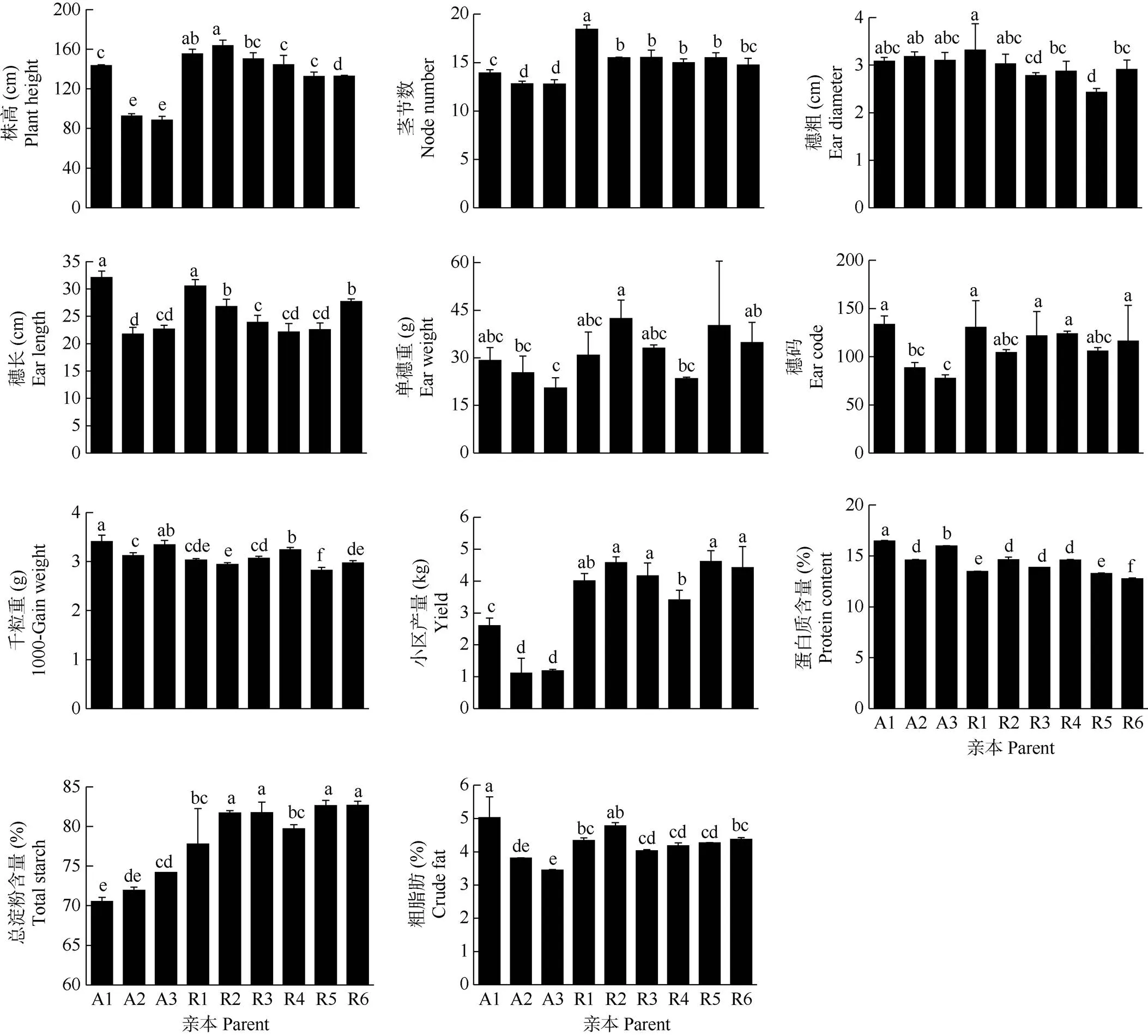
图1 不同亲本的产量和品质性状。A1~A3和R1~R6见表1。柱上不同字母表示差异显著(P<0.05)。以下图表同。
综上所述,所配的组合在株高、穗长、穗重、小区单产等性状的杂种优势较为明显。从小区产量超亲优势来看,A1×R4和A2×R4为产量强优势组合,A1×R6、A3×R4、A1×R1、A2×R1、A1×R2和A2×R3属于中优势组合。
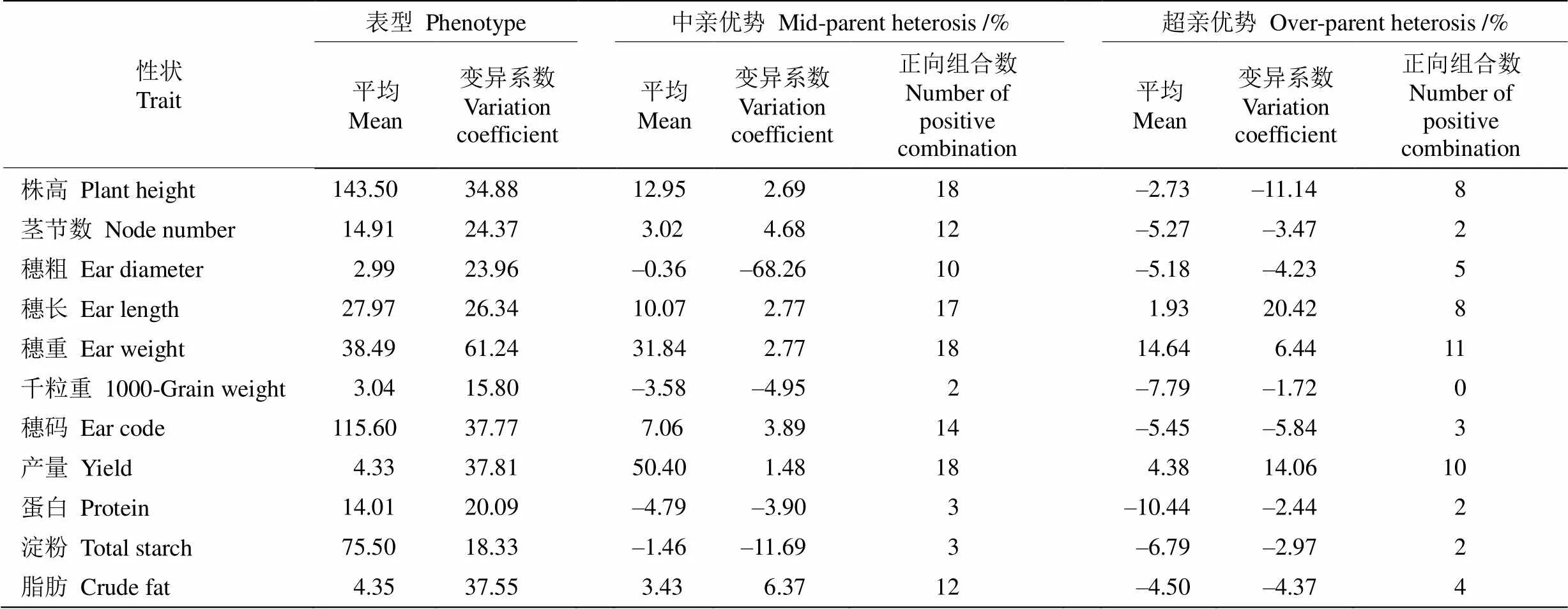
表2 谷子杂交组合产量和品质性状的中亲优势和超亲优势变异分析
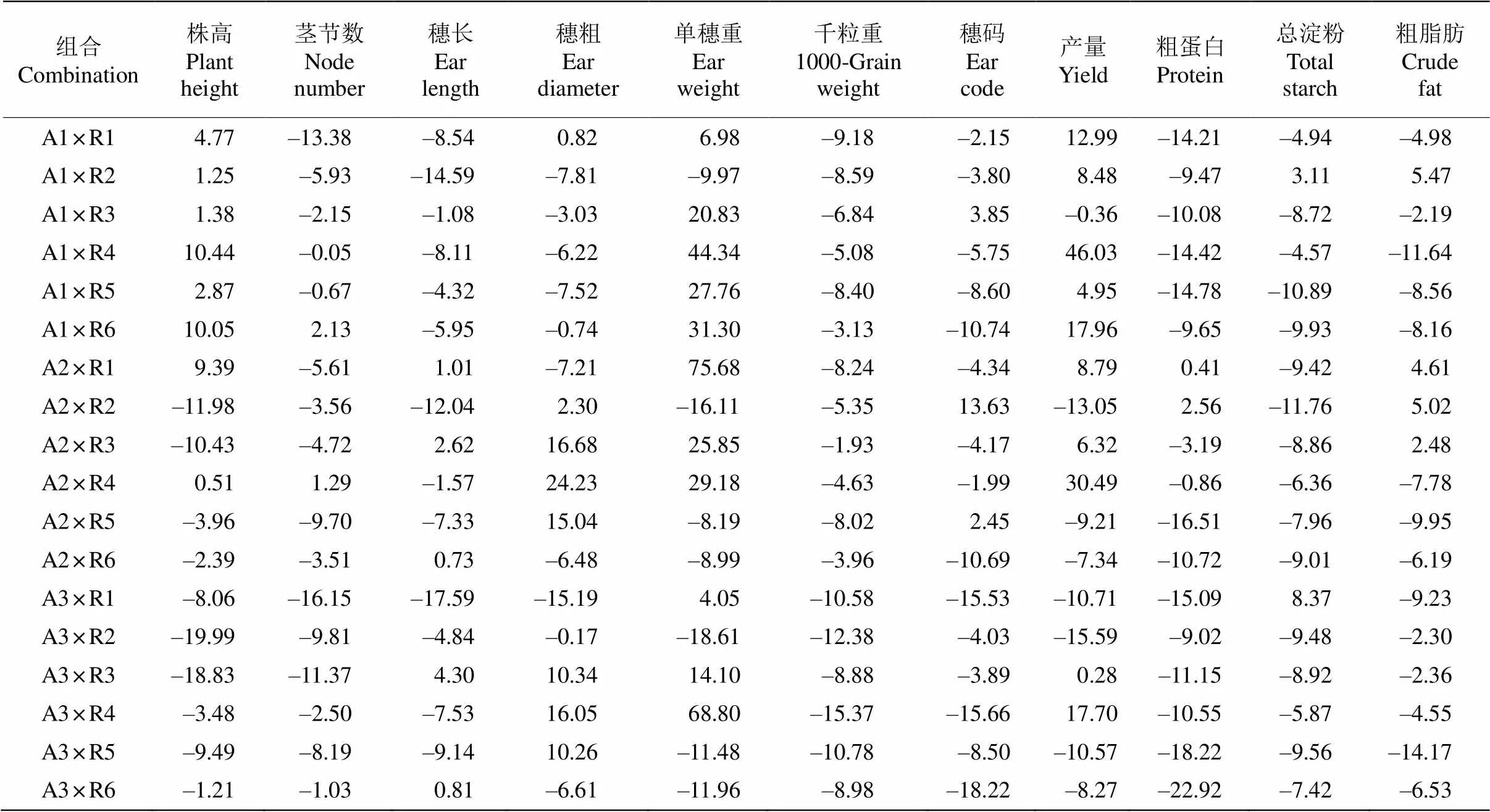
表3 谷子两系杂交组合的产量和品质性状的超亲优势(%)
2.3 杂交组合产量和品质性状的配合力方差分析
对杂交组合间的株高、穗长、蛋白质含量等11个性状进行方差分析(表4),结果表明除茎节数外,其他性状在区组间差异不显著。除单穗重和小区产量外,其他性状的表型在不同组合间存在显著或极显著差异(<0.05)。组合间方差是由父母本一般配合力(GCA)效应值方差和各组合特殊配合力(SCA)效应值方差分量组成。本研究以组合表型数据的平均值为单位,对亲本间的GCA效应值和杂交组合间的SCA效应值进行方差分析(表5),结果显示除单穗重外,其他性状的GCA效应和SCA效应均达极显著水平(0.01)。从GCA与SCA的效应值均方比值来看,小区产量的加性遗传均方为非加性遗传均方的10倍以上,而穗粗、单穗重、总淀粉和粗脂肪等性状的加性及非加性遗传均方间的差异较小。

表4 18个组合11个性状和配合力的方差分析
MSs: 一般配合力均方; MSg: 特殊配合力均方; **:<0.05; *:<0.01。
MSs: Mean square of general combining ability; MSg: Special combining ability; **:<0.05; *:<0.01.
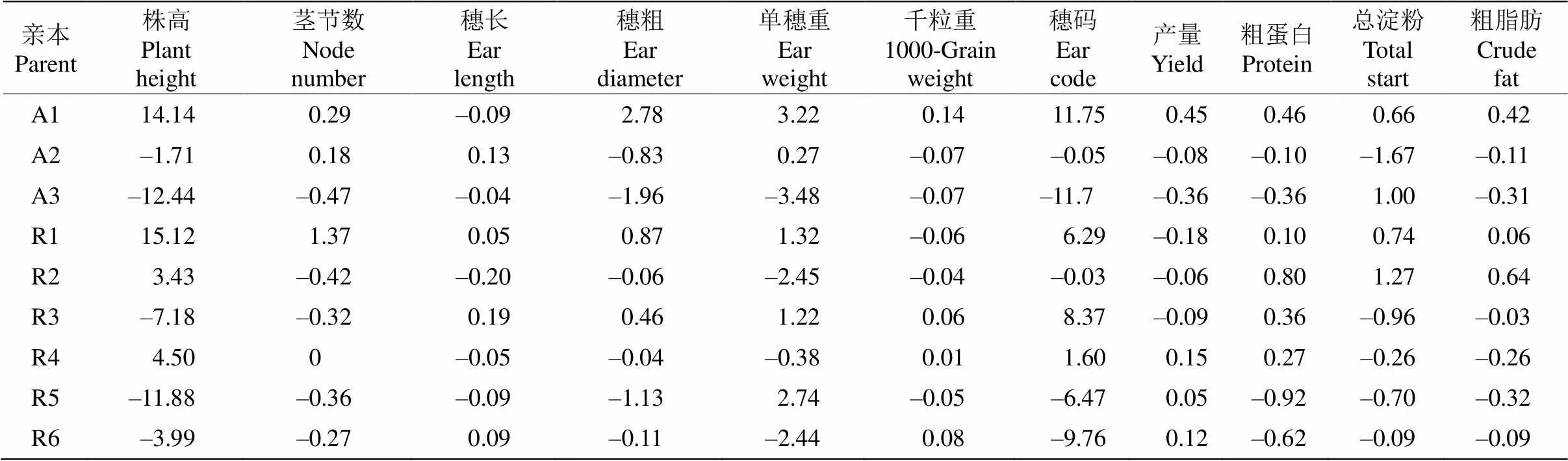
表5 亲本产量和品质相关性状的一般配合力(GCA)效应
2.4 亲本产量和品质性状的一般配合力效应
株高、产量和品质性状是杂交谷子育种最为重要的数量性状,对亲本的产量和品质性状的一般配合力效应值进行分析,从表5可见不同亲本的同一性状、同一亲本不同性状的GCA效应值存在较大差异。在3个不育系和6个恢复系中,亲本A1、R1、R2和R4的株高和茎节数GCA效应值为正值,而亲本A2、A3、R3、R5和R6的相应GCA效应值为负值。其中亲本A1的GCA效应值最高(14.14),亲本A2的最低(-12.44)。恢复系R5为父本,不育系A2、A3做母本组配的组合F1株高显著降低了10%~21% (图2: A)。不育系A3、A2和恢复系R2、R4、R5的穗粗、穗长、穗重、千粒重、穗码和小区产量GCA效应值为负值的较多,而A1、R1、R3、R6相应性状效应值均为正值的较少(图2: B)。由此表明,以A1、R1、R3和R6为亲本较易配制出高产的杂交组合,如组合A1×R6小区产量显著提高了22% (=0.008)。此外,品质性状是评价小米加工特性研究的主要内容。从粗蛋白质、总淀粉和粗脂肪含量的GCA效应值来看(表5),A1、R1、R2相应性状的GCA效应值均为正值,可组配出高蛋白的小米品种。

图2 组合的株高、茎节数(A)和亲本产量性状的一般配合力(B)
综上所述,不同谷子亲本在产量和品质性状上的GCA效应值有明显区别。从亲本GCA效应值来看,使用A1、R2较易组配出具高结实率、穗码多、穗长等特点的高产优质杂交新组合。
2.5 组合的产量和品质性状的特殊配合力分析
特殊配合力(SCA)是两个自交系通过相互作用才能表现出的非加性效应,即基因的显性和上位效应,虽不能稳定遗传,但可为杂种优势的利用和杂交种的选育提供重要信息。由表6可见,18个组合的SCA效应值变化最大的性状是株高、千粒重和总蛋白含量。株高的特殊配合力较低的组合有3个,分别是组合A1×R1 (–9.89)、A2×R6 (–8.25)和A3× R2 (–3.52),其中A2×R6、A3×R2的株高和茎节数最低。穗粗、穗长、穗重、千粒重和穗码的SCA效应值为-7.40~4.59,其SCA效应值为正值的组合有8个,分别为A1×R5、A1×R6、A2×R1、A2×R2、A2×R3、A2×R4、A3×R3和A3×R5。其中组合A2× R4、A3×R3千粒重的SCA效应值最高,分别为4.25和4.59。品质性状粗蛋白、总淀粉、粗脂肪的SCA效应值为-4.11~7.06,表现正向效应的组合分别有9、8和6个。其中,总淀粉SCA效应值最大的组合是A3×R1,最小的组合是A2×R1;粗蛋白SCA效应值最大的组合是A1×R6,最小的组合是A2× R5;粗脂肪SCA效应值最大的组合是A2×R1,最小的组合是A2×R4。
从谷子亲本及其组合产量和品质性状的GCA和SCA效应值综合来看,有的双亲GCA效应值呈现较高正向效应,但其后代的SCA效应值并不一定高,甚至表现为负向效应。如A1和R1株高的GCA为正效应,分别为14.14和15.12,而组合A1×R1的SCA为负效应值(–9.89)。有的双亲的GCA效应值为负值,而组合的SCA效应值为正值。如亲本A3和R5株高的SCA效应值分别为–12.44和–11.88, 其组合A3×R5的SCA为0.80。此外,大多数组合不同性状的SCA表现并不一致,其效应值呈现正负两种效应。如A1×R2组合的穗粗、穗长和穗重SCA效应值均为负值,而其千粒重和穗码的SCA效应值为正值,A1×R5组合除穗重和总淀粉含量外,其他产量和品质性状的SCA效应值均为正值。这表明GCA与SCA间的关系较为复杂,不同组合杂种优势的产生与特异性状条件关系密切,不同性状对组合SCA的影响不尽相同。
2.6 杂交组合的产量和品质性状遗传参数分析
本研究对产量和品质性状的遗传力和环境效应对后代表型性状的影响进行了分析(表7),结果表明,除穗重、穗粗、小区产量外,其他性状都具有较高的广义遗传力和狭义遗传力,其中茎节数、千粒重和蛋白含量的广义遗传力与狭义遗传力相差较大。这表明以上性状的非加性遗传作用较为突出,受环境影响较大。而其他性状的广义遗传力与狭义遗传力相当,如株高、穗长、脂肪含量的广义遗传力和狭义遗传力较大,均超过65%,加性方差大于显性方差,表明这些性状主要受加性效应影响,受环境影响较小,可通过选择来改良育种材料的遗传组成,并且可早期世代选择。此外,穗重、蛋白和淀粉含量的显性方差大于加性方差,说明显性效应起主导作用。因此,品质性状中小米总淀粉含量受显性效应决定,产量性状中穗长、千粒重、穗码、产量受加性效应决定,而其他性状受显性效应、加性效应的共同作用。总淀粉含量的狭义遗传力为0,广义遗传力为97.12%,说明其非加性遗传作用更为突出,适合晚期世代选择。
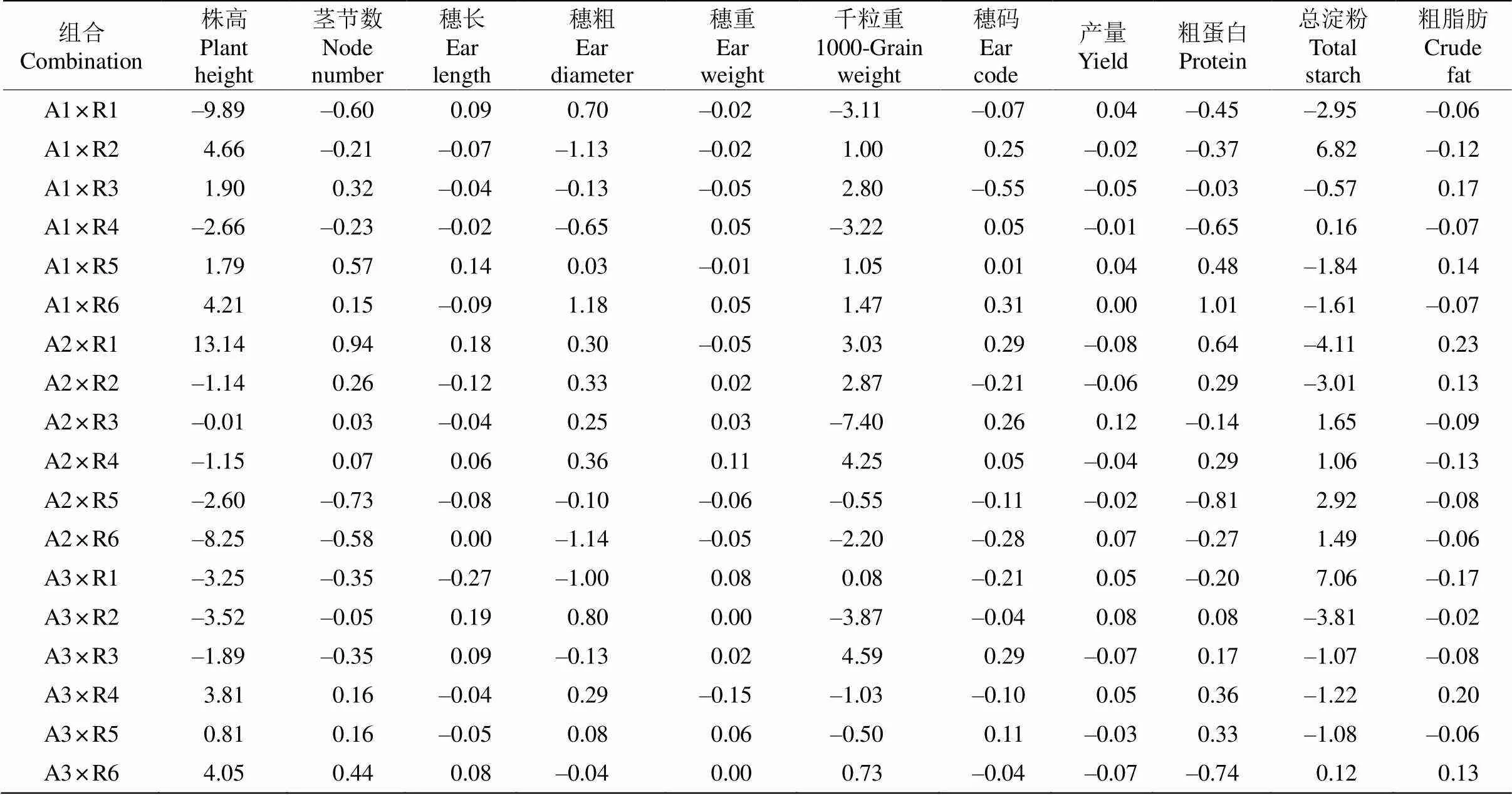
表6 18个谷子组合的产量和品质性状SCA效应值
表7 谷子杂交组合的产量和品质性状遗传参数估计
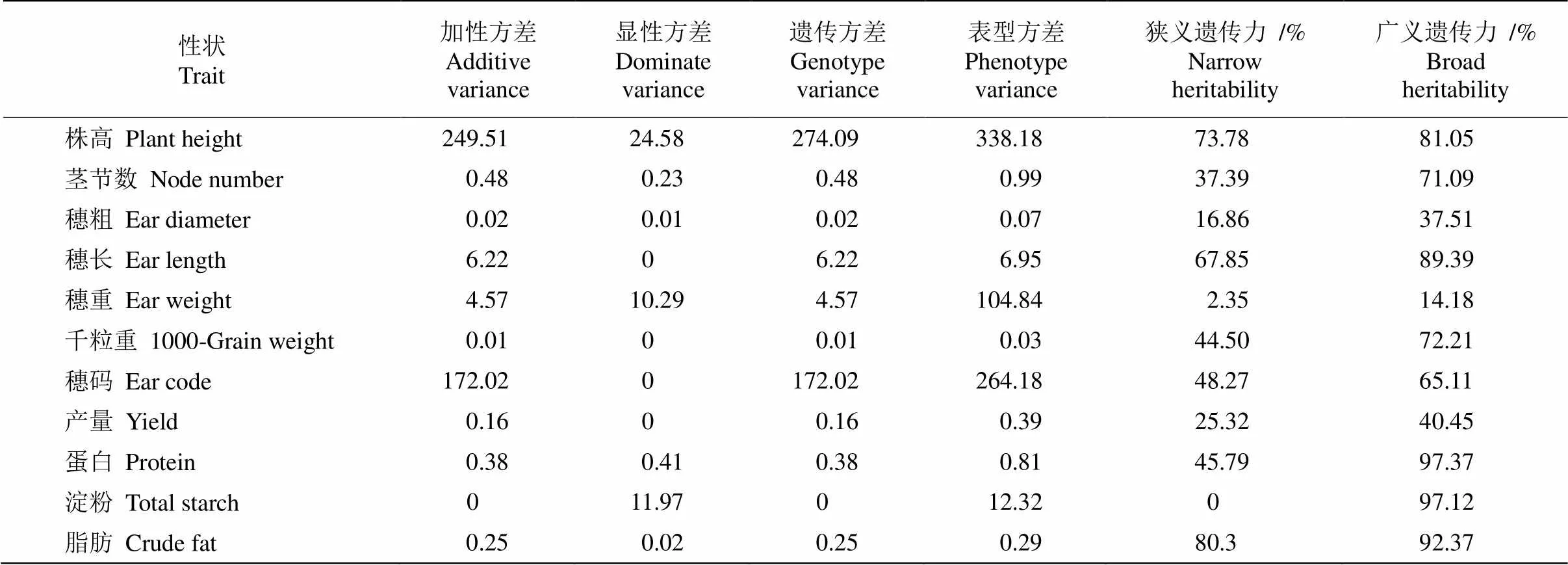
Table 8 Estimation of genetic parameter for yield and quality characters of hybrid combinations in Setaria italica
产量和品质性状是作物最重要的数量性状,其表现受基因和环境因子的共同作用。由于基因的一因多效及多因一效导致不同性状间存在不同程度的联系,性状间也相互产生一定的联系和影响。为探究产量、品质性状间的关系,对谷子两系杂交组合产量与品质相关的11个性状进行相关性分析。图3结果表明,株高、茎节数、穗长、穗重、千粒重、穗码、小区产量、粗蛋白和粗脂肪含量间存在显著或极显著正相关,小区产量还与穗码、千粒重、穗重、穗长、茎节数、株高存在显著正相关(<0.05)。粗蛋白、粗脂肪含量分别与穗码、穗长和株高性状存在显著相关,总淀粉含量与穗粗间的相关系数为–0.59 (<0.05)。而总淀粉与粗蛋白、粗脂肪间的相关系数分别为–0.16和0.015,均达显著水平。这表明产量构成因素(千粒重、单穗粒重、株高、穗长)与品质性状(粗蛋白、粗脂肪、总淀粉)间无显著关联。
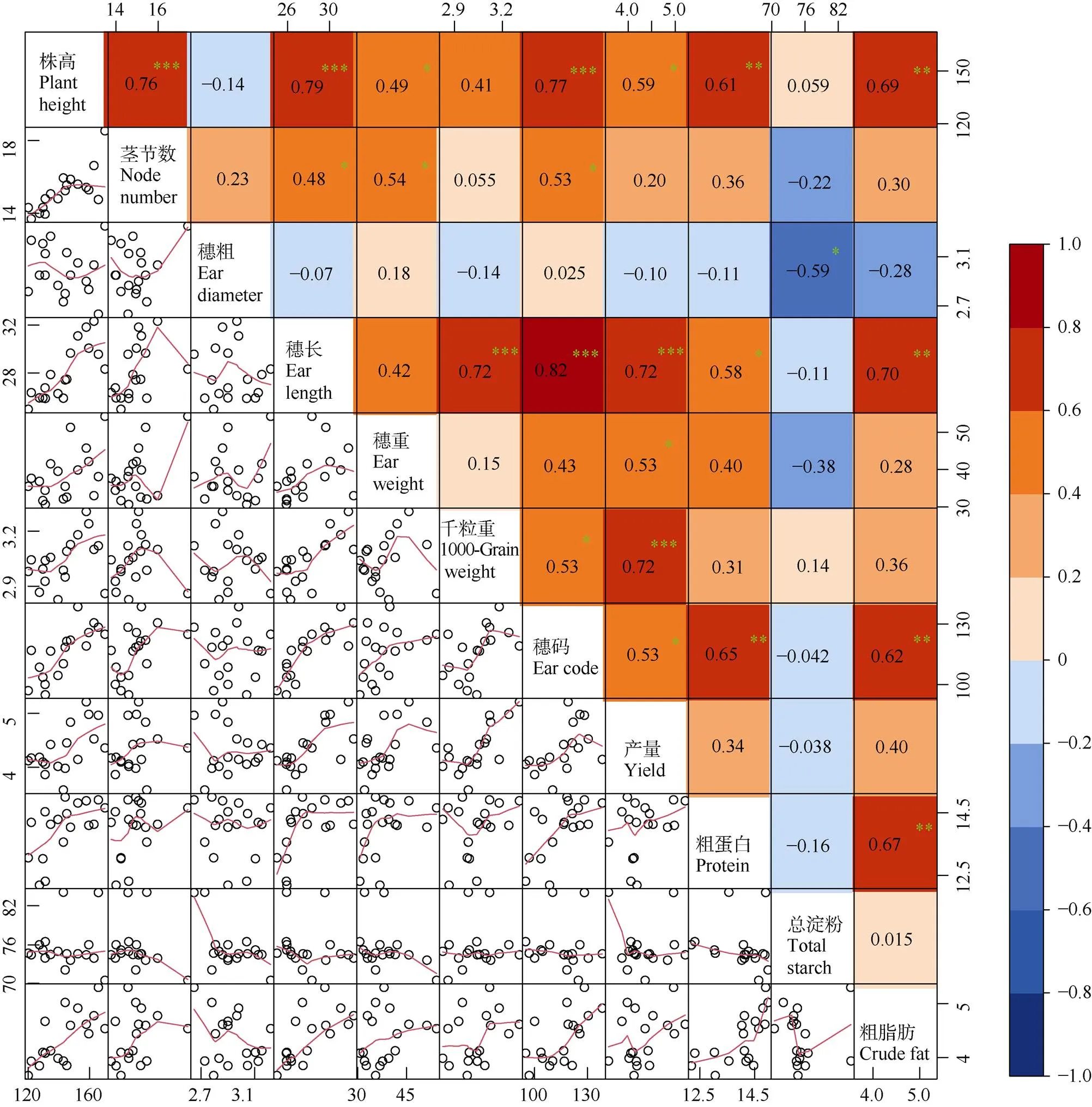
图3 谷子杂交组合产量性状与品质性状的相关性。右上: 相关系数; 左下: 散点图; **: P<0.05; *: P<0.01。
3 结论和讨论
杂种优势的广泛利用有效提高了作物的产量、抗逆性和适应性,为解决世界粮食短缺和品质差等问题做出了重大贡献[15]。在杂种优势利用方面,配合力是作物强优势亲本选择的重要基础,育种实践表明杂种后代的性状表现通常与亲本的配合力密切相关[16–17]。本研究对谷子18个杂交组合及其亲本的产量、品质性状配合力分析表明,除蛋白质含量外不育系其他性状的配合力显著低于恢复系,且同一亲本不同性状的GCA效应值存在显著差异, 同一性状不同亲本的GCA效应值也不同。而且同一组合不同性状或同一性状不同组合的SCA效应值也存在明显差异。这与张征等[18]对杂交水稻农艺性状配合力的研究结果一致。本研究结果还表明, 有些亲本的产量性状或品质性状的一般配合力高, 但后代对应性状的SCA效应值不高,甚至表现负向效应,如A1和R1株高的GCA为正效应,而组合A1×R1的SCA效应值为负。所以在谷子传统育种过程中必须重视配合力效应值,不仅要考虑双亲特定性状的GCA效应,而且还要考虑特定组合的SCA效应值。
就优质育种的综合利用价值来看,本研究结果表明,产量性状具有较强的中亲优势,部分组合具有较高的超亲优势。在9个亲本中,A1 (不育系)和R1 (恢复系)的产量和品质性状的GCA正效应值较大,是改良高产优质的优良亲本。谷子品质性状的杂种优势效应均介于亲本之间,并倾向于表现值较低的亲本。优异的杂交组合为A1×R5、A1×R6、A2× R1、A2×R4,与超高亲本的产量相比,杂交组合的产量能够提高15.5%。而A2×R2组合的粗蛋白和粗脂肪含量分别提高了2.5%和5.0%,产量却下降了11.7%。由于脱壳后小米的品质性状与产量性状的遗传关系较为复杂,暗示这些性状的基因加性效应及非加性效应间有着本质上的差异。此外,王昌华等[10]对水稻品质性状杂种优势的研究表明,组合的SCA效应值高低与其亲本GCA效应值高低并不一一对应,如亲本GCA效应值高的性状,其组合的SCA效应值不一定高,二者具有相对独立性。本研究结果还表明,小米粗蛋白和粗脂肪含量分别与穗码、株高和穗长存在显著正相关,而总淀粉含量除与穗粗存在显著负相关外,与其他产量性状的相关性均未达到显著水平。这与苗泽志等[19]的研究结果不尽相同,他报道小米粗蛋白含量与株高呈显著正相关,而与穗长和产量呈显著负相关;总淀粉含量与穗重和株高呈正相关,与穗码则呈负相关。水稻的总蛋白含量与结实率存在显著负相关,而直链淀粉含量与穗粒数也存在显著负相关[20]。但与丁璐等[21]报道的玉米杂交组合农艺性状的相关性结果相似,其结果表明玉米籽粒蛋白质和淀粉含量与穗长、百粒重、穗粒重和产量的相关性均未达显著水平。小米品质是一个较为复杂的综合性状,产量较高的组合其品质并不一定较好。因此,在今后的谷子育种工作和生产实践中,可根据育种目标和亲本的GCA效应值选配不同优良性状的组合,尤其是在谷子产量提升过程中还要兼顾小米的品质特性, 从而提高杂交谷子品种的利用和经济整体水平。
[1] PAN G T, YANG K C, LI W C, et al. A review of the research and application of heterotic groups and patterns of maize breeding in southwest China [J]. J Maize Sci, 2020, 28(1): 1–8. doi: 10.13597/j. cnki.maize.science.20200101.
潘光堂, 杨克诚, 李晚忱, 等. 我国西南玉米杂种优势群及其杂优模式研究与应用的回顾 [J]. 玉米科学, 2020, 28(1): 1–8. doi: 10. 13597/j.cnki.maize.science.20200101.
[2] LEI Y Q, SONG S F, LI X Q. Development of technologies for hete- rosis utilization in rice [J]. Hybrid Rice, 2017, 32(3): 1–4,9. doi: 10. 16267/j.cnki.1005-3956.201703001.
雷永群, 宋书锋, 李新奇. 水稻杂种优势利用技术的发展 [J]. 杂交水稻, 2017, 32(3): 1–4,9. doi: 10.16267/j.cnki.1005-3956.201703001.
[3] ZHAO J P, REN J C, GUO P Y, et al. Discussion about the high-yield wheat breeding and the utilization of hybrid vigor [J]. J Shanxi Agric Sci, 2018, 46(11): 1963–1966. doi: 10.3969/j.issn.1002-2481.2018. 11.45.
赵吉平, 任杰成, 郭鹏燕, 等. 论小麦高产育种与杂种优势的利用[J]. 山西农业科学, 2018, 46(11): 1963–1966. doi: 10.3969/j.issn.1002- 2481.2018.11.45.
[4] XING C Z, GUO L P, LI W, et al. Ten-year achievements and future development of cotton heterosis utilization [J]. Cotton Sci, 2017, 29 (S1): 28–36. doi: 10.11963/1002-7807.xczxcz.20170825.
邢朝柱, 郭立平, 李威, 等. 棉花杂种优势利用研究十年成就和未来发展 [J]. 棉花学报, 2017, 29(S1): 28–36. doi: 10.11963/1002-7807. xczxcz.20170825.
[5] CHEN J X, ZHOU H, XIE W B, et al. Genome-wide association analyses reveal the genetic basis of combining ability in rice [J]. Plant Biotechnol J, 2019, 17(11): 2211–2222. doi: 10.1111/pbi.13134.
[6] JIA G Q, HUANG X H, ZHI H, et al. A haplotype map of genomic variations and genome-wide association studies of agronomic traits in foxtail millet () [J]. Nat Genet, 2013, 45(8): 957–961. doi: 10.1038/ng.2673.
[7] QIU F C, FENG X L. Review, current situation and development direction of utilization of hybrid millet in China [J]. China Seed Ind, 2013(3): 11–12. doi: 10.19462/j.cnki.1671-895x.2013.03.006.
邱风仓, 冯小磊. 我国谷子杂优利用回顾、现状与发展方向 [J]. 中国种业, 2013(3): 11–12. doi: 10.19462/j.cnki.1671-895x.2013.03.006.
[8] LI S D, DONG X K, FAN G Y, et al. Comprehensive profiling and inheritance patterns of metabolites in foxtail millet [J]. Front Plant Sci, 2018, 9: 1716. doi: 10.3389/fpls.2018.01716.
[9] TIAN G, WANG Y W, LI H X, et al. Breeding and studies of foxtail millet hybrid Changzagu 2 with herbicide-resistance [J]. J Agric Sci Technol, 2009, 11(S2): 138–141. doi: 10.3969/j.issn.1008-0864.2009. z2.039.
田岗, 王玉文, 李会霞, 等. 抗除草剂谷子杂交种长杂谷2号选育研究[J]. 中国农业科技导报, 2009, 11(S2): 138–141. doi: 10.3969/j. issn.1008-0864.2009.z2.039.
[10] WANG C H, WU T H, ZHANC Y Z, et al. Analysis of relativity between main quality traits, combining ability and heterosis of hybrid rice in North China [J]. SW China J Agric Sci, 2011, 24(2): 385–390. doi: 10.3969/j.issn.1001-4829.2011.02.001.
王昌华, 吴天华, 张燕之, 等. 北方杂交粳稻品质性状杂种优势、配合力及遗传效应分析[J]. 西南农业学报, 2011, 24(2): 385–390. doi: 10.3969/j.issn.1001-4829.2011.02.001.
[11] QU Y J, SUN J L, GENG X L, et al. Correlation between genetic distance of parents and heterosis in upland cotton [J]. Sci Agric Sin, 2019, 52(9): 1488–1500. doi: 10.3864/j.issn.0578-1752.2019.09.002.
曲玉杰, 孙君灵, 耿晓丽, 等. 陆地棉亲本间遗传距离与杂种优势的相关性研究[J]. 中国农业科学, 2019, 52(9): 1488–1500. doi: 10. 3864/j.issn.0578-1752.2019.09.002.
[12] LIU Z L, BAI G H, ZHANG D D, et al. Genetic diversity and popu- lation structure of elite foxtail millet [(L.) P. Beauv.] germplasm in China [J]. Crop Sci, 2011, 51(4): 1655–1663. doi: 10. 2135/cropsci2010.11.0643.
[13] LI S Y, LIU D, LI Q, et al. Phenotype identification in heterosis of yield and yield traits of foxtail millet hybrids [J]. J Henan Agric Sci, 2018, 47(8): 28–34. doi: 10.15933/j.cnki.1004-3268.2018.08.006.
李素英, 刘丹, 李强, 等. 谷子杂交种产量及产量性状杂种优势的表现型鉴定[J]. 河南农业科学, 2018, 47(8): 28–34. doi: 10.15933/j. cnki.1004-3268.2018.08.006.
[14] COVARRUBIAS-PAZARAN G. Software update: Moving the R package sommer to multivariate mixed models for genome-assisted prediction [J]. bioRxiv, 2018: 354639. doi: 10.1101/354639.
[15] YUAN L P. Progress in breeding of super hybrid rice [J]. J Agric, 2016, 8(1): 71–73.
袁隆平. 超级杂交稻研究进展 [J]. 农学学报, 2018, 8(1): 71–73.
[16] XIE F L, SHI Z L, YANG S W, et al. Breeding techniques for improvement of parent utilization of combining abilities in heterosis utilization of crop [J]. J Shanxi Agric Sci, 2016, 44(1): 100–105. doi: 10.3969/j.issn.1002-2481.2016.01.27.
谢福来, 史忠良, 杨三维, 等. 作物杂种优势利用中亲本配合力改良的育种技术[J]. 山西农业科学, 2016, 44(1): 100–105. doi: 10. 3969/j.issn.1002-2481.2016.01.27.
[17] ZHANG L, JING S L, ZHOU Y Q, et al. The summary of research on combining ability [J]. Bot Res, 2013, 2(1): 18–23. doi: 10.12677/BR. 2013.21004.
张磊, 荆胜利, 周晏秋, 等. 亲本配合力的研究概述[J]. 植物研究, 2013, 2(1): 18–23. doi: 10.12677/BR.2013.21004.
[18] ZHANG Z, ZHANG X L, MO B C, et al. Combining ability analysis of agronomic trait in×hybrid rice [J]. Acta Agron Sin, 2017, 43(10): 1448–1457. doi: 10.3724/sp.j.1006.2017.01448.
张征, 张雪丽, 莫博程, 等. 籼型杂交水稻农艺性状的配合力研究[J]. 作物学报, 2017, 43(10): 1448–1457. doi: 10.3724/sp.j.1006.2017.01448.
[19] MIAO Z Z, HAN H K, DU W J, et al. Principal component analysis on traits related to yield and quality of hybrid millet [J]. J Shanxi Agric Sci, 2013, 41(8): 785–788,812. doi: 10.3969/j.issn.1002-2481.2013.08.06.
苗泽志, 韩浩坤, 杜伟建, 等. 杂交谷子产量及品质相关性状的主成分分析 [J]. 山西农业科学, 2013, 41(8): 785–788,812. doi: 10. 3969/j.issn.1002-2481.2013.08.06.
[20] LI G P, GUO J F, TANG N, et al. Study on correlation of quality characters and between quality characters and agronomic characters of hybrid rice [J]. J Anhui Agric Sci, 2007, 35(35): 11439–11440,11443. doi: 10.3969/j.issn.0517-6611.2007.35.044.
李国鹏, 郭建夫, 汤能, 等. 杂交水稻品质性状间及品质性状与农艺性状间相关性研究 [J]. 安徽农业科学, 2007, 35(35): 11439– 11440,11443. doi: 10.3969/j.issn.0517-6611.2007.35.044.
[21] DING L, LIU H X, WANG Y S, et al. Correlation and principal component analysis of the agronomic traits of 26 corn hybrids combi- nations [J]. Mol Plant Breed, 2020, 18(3): 995–1002. doi: 10.13271/j. mpb.018.000995.
丁璐, 刘海学, 王聿双, 等. 26个玉米杂交组合农艺性状的相关性与主成分分析[J]. 分子植物育种, 2020, 18(3): 995–1002. doi: 10. 13271/j.mpb.018.000995.
Heterosis and Genetic Characteristics Analysis for Yield and Quality Related Characters in
SHI Guang-yan1, WANG Xiao-qi2a, HAN Yuan-huai2a, YANG Cheng-yuan1, MA Hui-fang1, ZHAO Xiong-wei2b*, QIAO Zhi-jun3*
(1. Industrial Crop Institute, Shanxi Agricultural University,Fenyang 032200, Shanxi, China; 2a. College of Agronomy; 2b. College of Life Sciences, Shanxi Agricultural University, Taigu 030801, Shanxi, China; 3. Center for Agricultural Genetic Resources Research, Shanxi Agricultural University,Taiyuan 030031, China)
In order to understand the heterosis of yield-related and quality characters in two-line hybrid progenies of, 18 hybrid combinations were designed with three male sterile lines (A1, A2, A3) and six herbicide resistant restorer lines (R1, R2, R3, R4, R5 and R6) by using incomplete diallel cross (NCⅡ), the eleven phenotypic characteristics of combining ability and the hereditability were analyzed, such as plant height, grain weight, protein content, and so on. The results showed that there were significant differences in quantitative traits among two parents and their combinations. Among all the traits, the plot yield of hybrid combination was significantly correlated with thousand-seed weight (TSW), ear weight (EW), plant height (PH), and ear length (EL), but not significantly related with quality traits, such as crude protein, crude fat and total starch. The general combining ability (GCA) effect values of different characters varied significantly in different parents. Meanwhile, A1 (male sterile line) and R1 (restorer line) had high positive effect values of GCA among most of yield and quality traits, which was potential parents for improving the high yield and quality of foxtail millet. However, it was noteworthy that the presentation of specific special combining ability (SCA) in combinations was not completely consistent witheffect values of GCA in their parents. Among eighteen combinations, A1×R5, A1×R6, A2×R1, A2×R4 displayed high SCA effect, and showing excellent combinations. Furthermore, the broad-sense heritability and narrow-sense heritability of all quantitative traits were more than 65%, mainly co-controlled by additive effects and non-additive effects. PH, EW, and yield ofhybrids had significant mid parent heterosis, and some combinations had strong super parent heterosis. In addition, the narrow-sense heritability value of PH, EL TSW, ear code, and crude fat content were higher than that of other traits, while the non-additive genetic effects of EW, stem node number and protein content were more prominent. Although the heterosis characteristics of yield and quality traits were different, they were determined by additive and dominance effect in the total genetic variation.
; Yield; Quality; Heterosis; Combining ability; Heredity
10.11926/jtsb.4296
2020–08–18
2020–11–01
山西农业大学省部共建有机旱作农业国家重点实验室自主研发项目(202105D121008-2-4); 山西农业大学生物育种工程项目(YZGC052);吕梁市重点研发项目(2019NYZDYF20); 财政部和农业农村部国家现代农业产业技术体系项目; 山西省高等学校科技创新项目(2019L0367); 山西省优秀博士来晋工作奖励资金科研项目(SXYBKY2019006)资助
This work was supported by the Project of State Key Laboratory of Integrative Sustainable Dryland Agriculture (in preparation), Shanxi Agricultural University (Grant No. 202105D121008-2-4); the Project for the Project for Biological Breeding Engineering of Shanxi Agricultural University (Grant No. YZGC052); the Project for Key Research and Development inLyuliang City (Grant No. 2019NYZDYF20); the Project for China Agriculture Research System of MOF and MARA; the Project for Scientific and Technological Innovation of Higher Education Institutions in Shanxi (Grant No. 2019L0367); and the Project for Excellent Doctor’s Work Award Fund in Shanxi Province (Grant No. SXYBKY2019006).
史关燕, 女, 副研究员, 主要从事谷子杂种优势利用研究。E-mail: jzsguanyan@126.com
E-mail: xwzhao@sxau.edu.cn; nkypzs@126.com
猜你喜欢
杂志排行

热带亚热带植物学报的其它文章
- Advances in Chemical Constituents and Pharmacological Activities of Pleuropterus multiflorus
- 成都地区19个木芙蓉品种的孢粉学研究及其分类学意义
- 秋石斛花青素提取液成分分析及其体外抗氧化活性和刺激性研究
- 龙珠果茎段离体培养和组培苗耐盐性分析
- Cloning and Expression Analysis on PmPGK1 and PmGPIC Genes in Pinus massoniana
- Michelia taishanensis Y. H. Tong, X. E. Ye, X. H. Ye & Yu Q. Chen (Magnoliaceae), A New Species from Guangdong